
长牡蛎壳黑选育系F7代的生长规律和模型❋
2020-01-09 19:48徐成勋
中国海洋大学学报(自然科学版) 2020年2期
徐成勋, 李 琪
(海水养殖教育部重点实验室(中国海洋大学),山东 青岛 266003)
长牡蛎 (Crassostreagigas)又称太平洋牡蛎,具有生长快、抗逆性强、肉质鲜美和营养丰富等优点,是世界上分布最广、产量最高的经济贝类。自20世纪80年代,人工育苗技术日趋成熟,我国长牡蛎工厂化养殖技术得到迅猛发展,长牡蛎的养殖产量逐年攀升,至2016年,我国牡蛎产量超过483.4万t,占全国贝类养殖总量的33.4%[1]。近年来,随着养殖集约化程度的提高,养殖长牡蛎开始出现生长缓慢、死亡率高、产量降低等问题[2],不利于长牡蛎养殖业的健康持续发展。通过遗传育种,培育生长快、品质好、抗逆性强的新品种是摆脱长牡蛎产业困境的出路之一。本课题组自2010年对长牡蛎进行人工选择,培育出左右壳和外套膜均是黑色、生长快速的长牡蛎壳黑新品系[3],为长牡蛎新品种的培育提供了重要素材。
使用统计学方法对生物的生长进行研究并建立生长模型,不但可以了解生物的生长过程,还可以预测其生长规律[4]。目前,已有很多学者针对不同物种,从生理学和数学角度提出了多种不同模型,如马氏珠母贝(Pinctadamartensii)[5]、墨西哥湾扇贝(Argopectenirradiansconcentricus)[6-7]、淡水珍珠蚌 (Margaritiferamargaritifera)[8]、华贵栉孔扇贝 (Chlamysnobilis)[9]等重要的经济贝类。在长牡蛎中,仅见孔宁等[10]对长牡蛎F3快速生长选育群体的生长规律做过报道。目前针对长牡蛎壳黑新品系的研究,主要围绕生长性状的遗传改良[3],对环境适应能力[11]及壳色遗传机制[12]等方面,尚未有关于其生长特性方面的报道。
本研究利用定期采集的生物学数据,运用线性回归和3种常见的生长模型,分别对长牡蛎壳黑品系幼虫期和养成期的各生长性状进行了拟合。旨在揭示长牡蛎壳黑品系生长发育特征,为进一步开展选育工作及大范围的养殖生产提供理论依据。
1 材料和方法
1.1 材料来源
本研究所用材料为第七代长牡蛎壳黑选育群体 (F7)。自2010年本课题组对乳山湾 (36.8 °N,121.7°E)自然存在的野生长牡蛎开展选择育种工作,先后经过四代家系选育和两代群体选育,得到了第六代长牡蛎壳黑选育群体 (F6)。2016年6月,以壳黑F6为基础群体,通过对壳高进行选择,获得了壳黑F7。壳黑F7的人工育苗于山东省莱州海益苗业有限公司进行,后转移至山东省荣成市爱莲湾 (37.2°N,122.6°E)进行养成。
1.2 人工育苗及养成
解剖成熟的壳黑长牡蛎亲贝进行性别鉴定,鉴定后分别收集精子卵子,在50∶1的精卵比下将精子卵子充分混合受精。获得的受精卵置于20 m3的水泥池中,于23 ℃水温下进行孵化。
刚孵化出的D形幼虫密度控制为10个/mL,随着幼虫生长逐步调整。幼虫培育前期投喂饵料以球等鞭金藻(Isochrysisgalbana) 为主,壳高增至120 μm后增加扁藻(Platymonassp.)。培育水温稳定在23~24 ℃,持续充气。每天换水2次,换水量1/3~1/2。当眼点幼虫比例达20%左右时,用200片栉孔扇贝(Chlamysfarreri) 壳一串的附着基,垂悬于培育池进行采苗,每片附着基附着20~30个长牡蛎稚贝时结束采苗。
稚贝附着后,置于室外沉淀池暂养3~4周,确定稚贝充分变态后,转移至山东省荣成市爱莲湾,采用筏式吊绳养殖方式进行养成。长牡蛎吊绳养殖所用的夹苗绳为长度3~4 m、直径0.6~0.8 cm的聚乙烯绳,将附有稚贝的附着基加入苗绳,附着基间距15~20 cm,每绳夹15~20片,苗绳间距50 cm。夹苗绳底部挂重约0.2~0.3 kg的石块,以避免其因风浪相互缠绕。
1.3 采样及数据测量
幼虫阶段,每5天随机取样30个D形幼虫进行测量。幼虫的壳高、壳长在Olympus显微镜下用目微尺 (精度0.05 μm)测量记录。养成阶段分别在第120、150、180、200、240、270、290、330、360、380、410和450天时随机取样30个,用电子游标卡尺 (精度0.01 mm)测量壳高、壳长、壳宽,用电子天平测量个体总重;并现场测量记录养殖海域海水温度盐度。(温度使用水温计测量,盐度使用光学盐度计测量)。
1.4 数据处理及生长模型构建
利用SPSS22.0软件进行数据处理及模型构建。使用Levenberg-Marquardt迭代法[13-14]估计生长参数,迭代精度设为10-8,即当各待估生长参数前后两次迭代结果的相对误差与误差平方和的相对误差都小于10-8时,迭代运算结束,以收敛的结果作为参数的最终估计值。幼虫期壳高和壳长的变化采用线性回归,并分析壳高 (SH) 与壳长(SL) 的关系。养成期选择Von Bertalanffy、Gompertz和Logistic 3种常见的动物模型(各模型见表1) 拟合壳黑长牡蛎F7的总重变化过程,并采用曲线拟合方法分析总重与壳高、壳长和壳宽的关系。

表1 3种常见动物生长曲线模型
注:A:总重的总生长量或生长极限;B:增长初始值参数;k:总重的瞬时增长速率。
Note:A:thebiggesttheoreticalgrowthlimitoftotalweight; B:theinitialgrowthparameter; k:instantaneousgrowthrateoftotalweight.
2 结果和分析
2.1 幼虫期壳高与壳长的线性回归与相关
根据幼虫期各生长性状在不同日龄的平均值(见表2),分别作出壳高与日龄、壳长与日龄的散点图,观察发现壳高、壳长与日龄大致呈线性分布(见图1)。因此直接对壳高、壳长与日龄进行线性回归,得到壳高对日龄的回归方程为y=39.723 8+11.697 2x,R2=0.991;壳长对日龄的回归方程为y=31.346 2+9.802 8x,R2=0.986。式中:y为壳高或壳长 (μm);x为日龄 (d);R2为复相关系数。
分析幼虫期长牡蛎壳黑F7的壳高与壳长相关性。由壳高与壳长的散点图 (见图2)可知,壳黑群体幼虫期壳高与壳长大致成线性相关,经线性回归,求得回归方程为:y=4.867 0+1.175 0x,R2=0.989。式中,y为壳高 (μm),x为壳长 (μm), R2为拟合优度。回归直线与散点图相结合(见图2),发现实测值大致位于回归直线上,说明回归效果较好。
2.2 环境因子的变化
养成阶段,各取样时间养殖海域的水温呈明显季节变化(见图3),于200日龄时水温降至最低(3.8 ℃),后逐渐升高,于380日龄时达到最高值 (24.0 ℃)。不同采样时间养殖海域的盐度变化幅度不大,介于30.5~32.8之间。
2.3 养成期总重生长模型的比较
绘制长牡蛎壳黑F7养成期各总重在不同日龄的变化曲线(见图4)。120 ~ 270日龄间,壳黑F7的总重增长较缓,270日龄后其总重增长速度明显增快。360日龄时,总重没有增加,反而相比330日龄时稍有下降。360日龄后,总重增长十分迅速,至410日龄时,总重的增长速度开始减缓。分别使用3种常用的动物模型对养成期长牡蛎壳黑F7的总重与日龄的关系进行拟合。拟合后各模型参数、估计值、标准误、置信区间、拐点及拟合度如表3所示,得到VonBertalanffy、Gompertz和Logistic3种模型的回归方程分别为;y=3.134 8(1+0.186 4e0.0046x)3、y=2.237 6×106e-13.932 3e-0.000 6x和y=8.144 9×108/(1+3.340 0×108e-0.006 5x),其R2分别为0.955、0.940和0.945。作出各方程在不同日龄的预测曲线图 (见图4),由图中观察,3种模型预测曲线呈曲线增长,仅在360~410日龄时期与实测值略有差距,其余时期与壳黑F7总重的实际变化情况相差不大。其中VonBertalanffy模型的R2最高,为0.955,说明其与长牡蛎壳黑F7总重的变化情况最好。
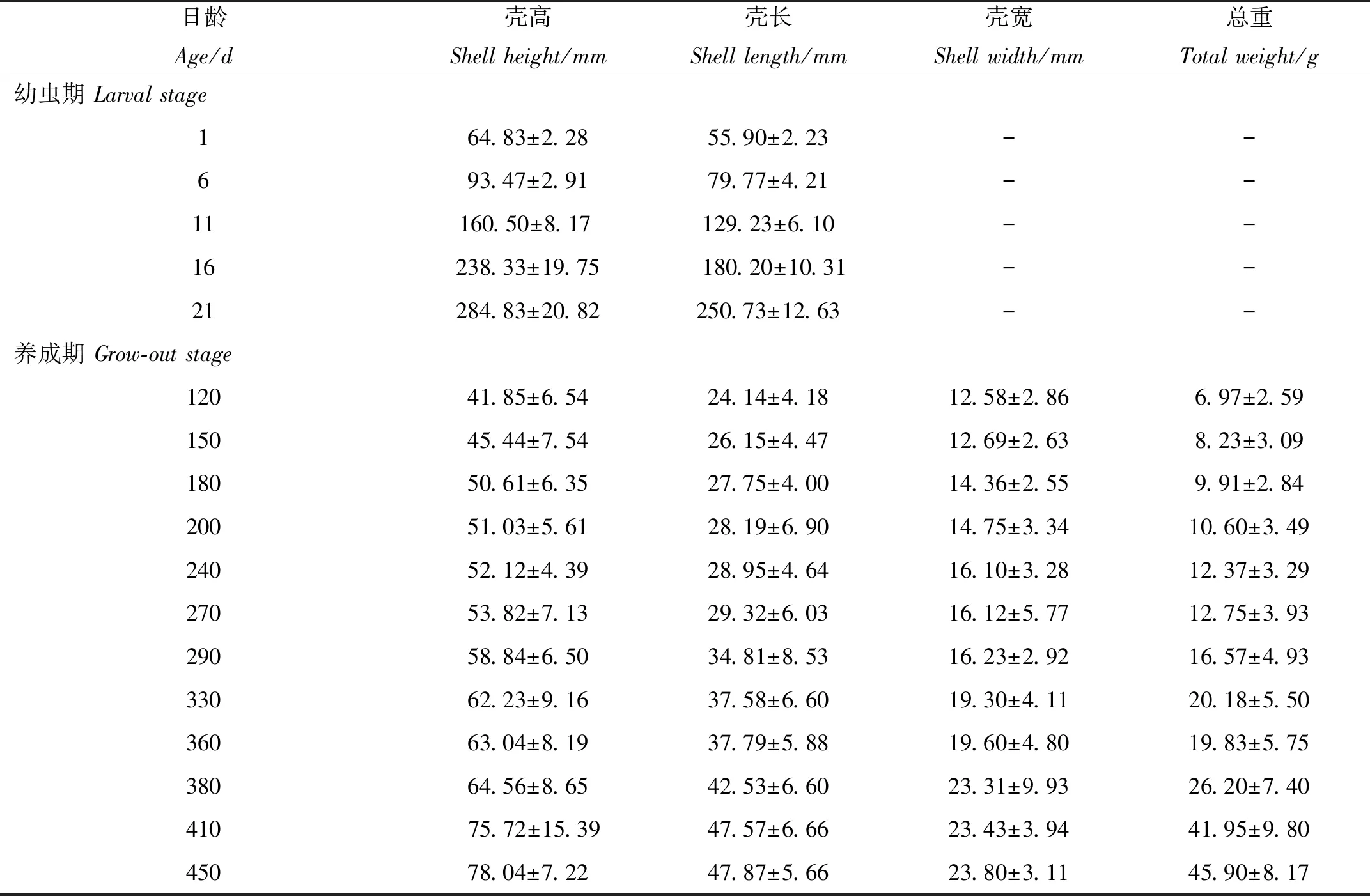
表2 不同日龄壳黑F7的壳高、壳长、壳宽和总重
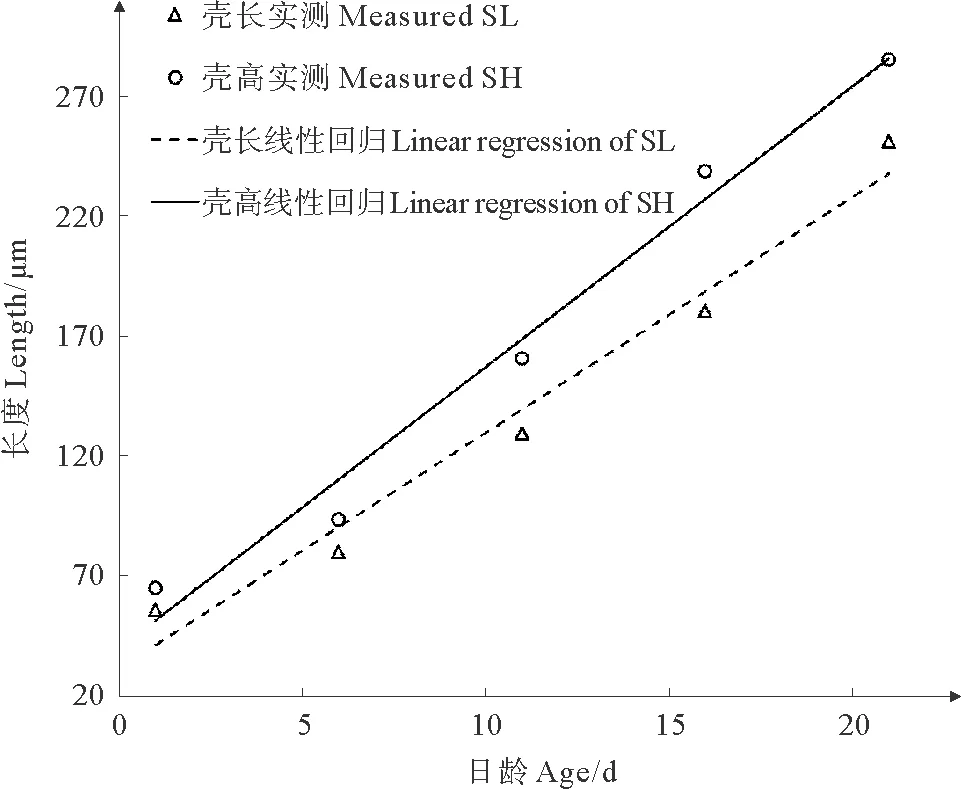
图1 幼虫期壳高、壳长与日龄的线性回归

图2 幼虫期壳高与壳长的线性相关

图3 不同日龄养殖海域海水水温和盐度变化
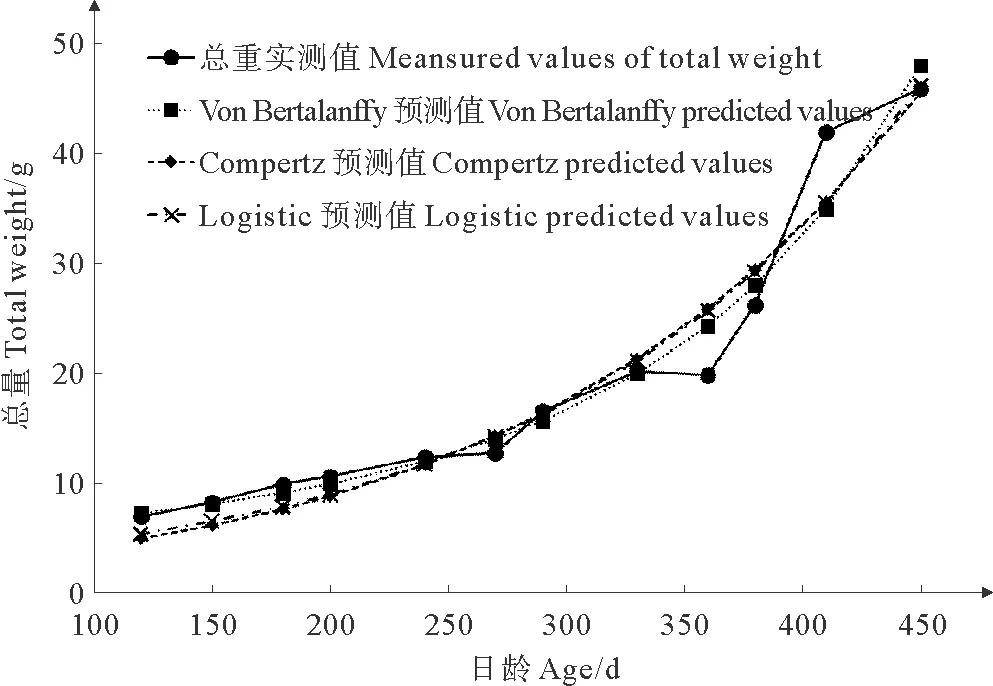
图4 壳长生长曲线与3种拟合曲线的比较
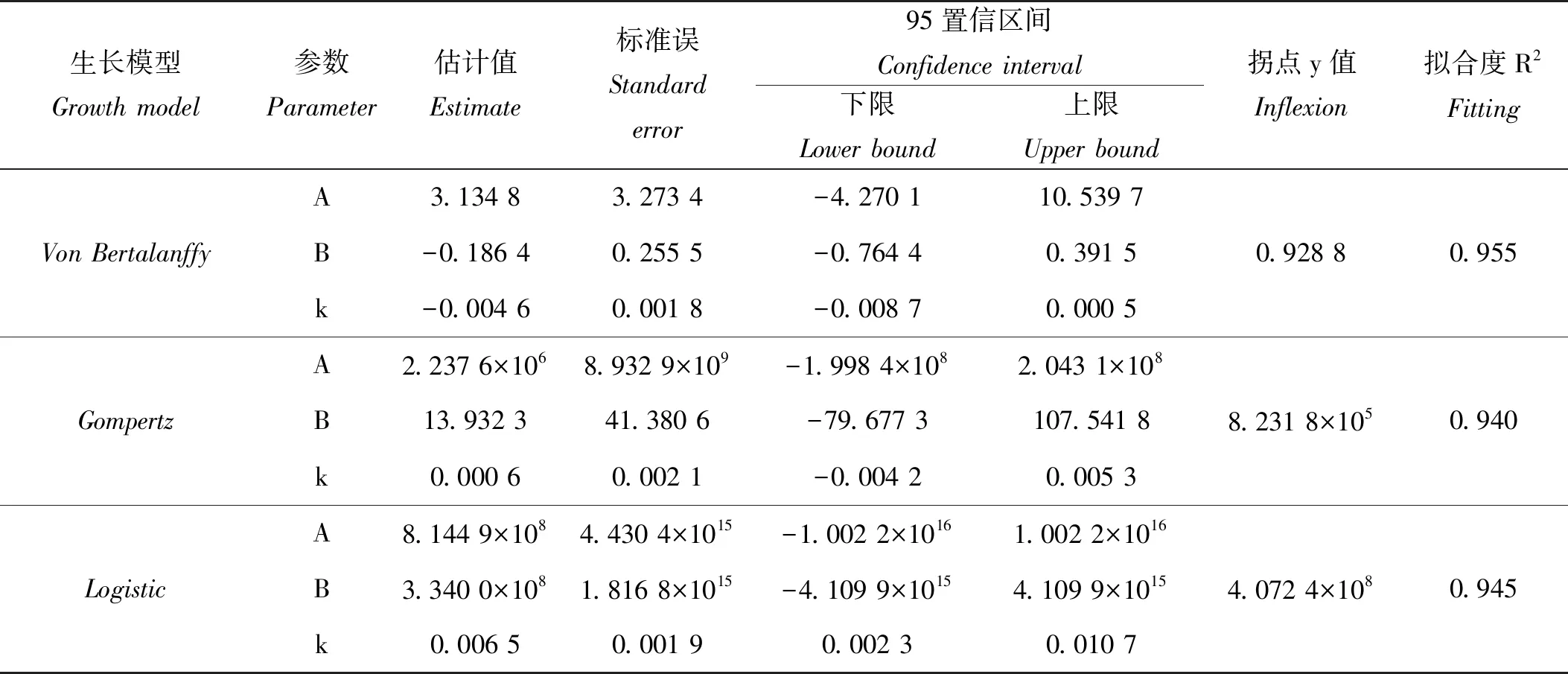
表3 3种生长模型的生长参数估计值与拟合度
2.4 养成期壳高、壳长、壳宽分别与总重的曲线拟合
分别作出壳高与总重、壳长与总重、壳宽与总重的散点图,发现壳高与总重,壳长与总重的散点图分布规律,大致呈曲线相关,而壳宽与总重的散点图分布离散,相关性较差(见图5)。分别采用线性方程等9种方程对养成期壳高、壳长、壳宽与总重的关系进行曲线拟合(见表4)。各方程的R2从0.80到0.99不等,R2越大说明曲线拟合效果越好。从拟合效果图上看(见图4),复合曲线、幂函数曲线、增长函数曲线与指数方程曲线与实测值的分布趋势大致相同,再结合R2及方程简洁程度,复合曲线在各组曲线拟合中均有不错的拟合效果。所以分别采用复合曲线方程y=1.706×1.055x、y=1.336×1.075x和y=1.287×1.154x来反映壳高与总重、壳长与总重、壳宽与总重的关系。
3 讨论
本研究中,长牡蛎壳黑F7壳高和壳长的增长规律符合线性回归方程(R2>0.98)。说明在幼虫期壳高与壳长的生长速率维持在相对稳定的水平,这与孔宁等[10]关于长牡蛎F3选育群体幼虫期生长规律的研究类似,在幼虫附着变态前,幼虫一直处于一个较高的生长速率。此外,壳高和壳长两直线方程斜率不同,壳高回归方程斜率大于壳长回归方程斜率,说明壳高增长速度快于壳长。长牡蛎幼虫期的壳高与壳长呈线性相关,R2=0.989说明拟合度较好,这表明可以通过一个性状有效预测另一个性状的生长规律。
本研究选用的VonBertalanffy、Gompertz和Logistic模型,都是常用于描述水产动物生长规律的模型[5,9-10,15]。符世伟等[5]对马氏珠母贝壳长生长模型的研究中,选用了Brody、VonBertalanffy、Gompertz、Logistic和Richards5种模型,采用Levenberg-Marquardt迭代法求出模型中的生长参数,确定马氏珠母贝壳长的生长过程遵循Brody生长模型。吴杨平等[15]在文蛤红壳色选育系F3的生长规律及模型研究中,确定文蛤红壳色选育系F3的壳长生长遵循Logistic生长模型。而本研究中,3种模型与长牡蛎壳黑F7养成期的生长规律都不能完全符合。比较R2,VonBertalanffy模型的拟合程度最好,但也仅限于对养成期长牡蛎壳黑F7体重的生长规律提供参考。其模型的拐点参数,并不符合实际的生长发育规律。此外,模型预测值与实际测量值在350~450日龄相差很大,这可能与长牡蛎生长的季节性变化有关。近350日龄,正值长牡蛎壳黑F7繁殖季节,其总重略有下降可能与精卵排放有关。360~410日龄期间,长牡蛎壳黑F7总重迅速升高,可能与其产期过后恢复发育有关。这与孔宁等[10]对长牡蛎F3选育群体的生长规律研究类似。此外,长牡蛎的波动性生长与生长环境(水温、饵料丰富度等)的周期性变化有着密切联系,夏季水温升高,光照充足,海水中饵料充足,有利于长牡蛎的快速增长。本实验中,360~410日龄期间正处夏季,有利的环境促使长牡蛎总重出现快速生长现象。冬季,海水温度降低,长牡蛎新陈代谢减缓,通常会导致长牡蛎生长减慢或停滞。与孔宁等[10]对长牡蛎F3选育群体的研究不同,本研究中长牡蛎壳黑F7总重在冬季并未发现明显的生长停滞,其仍能保持缓慢增长。造成这种差别一方面可能是由养成环境的差异造成。另一方面可能是对多代的群体选育对长牡蛎的环境适应能力发生了改变。通过壳高、壳长、壳宽与总重的曲线拟合,结果发现,壳黑F7的壳高、壳长和壳宽与总重的拟合效果较好,R2均大于0.99,如此高的拟合优度表明可以通过一个性状准确预测另一个性状的生长规律。
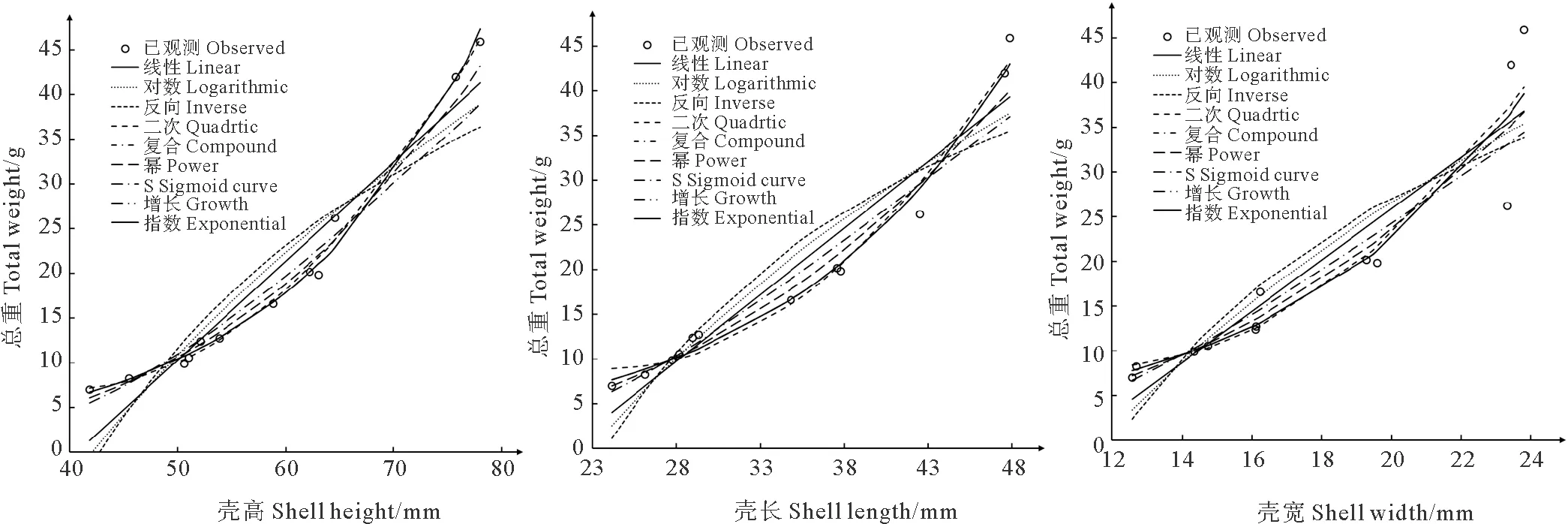
图5 壳高、壳长、壳宽分别与总重的曲线拟合效果
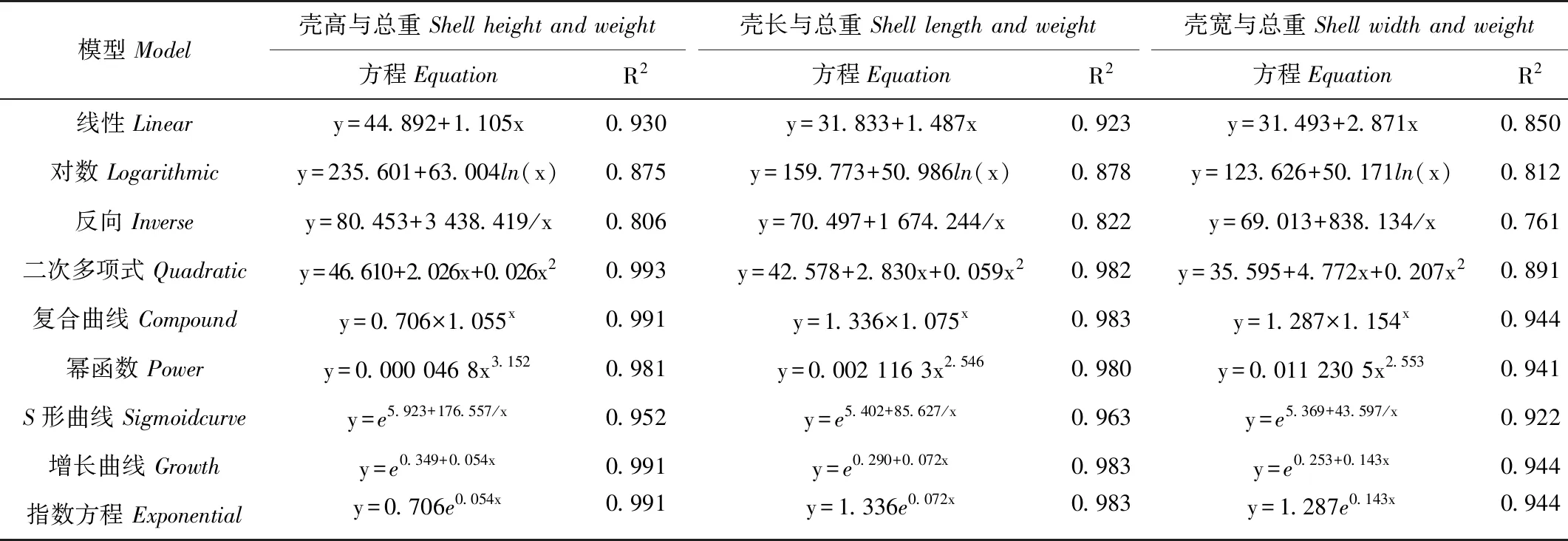
表4 壳高、壳长、壳宽分别与总重的关系
注:表中y为总重,x为对应的壳高、壳长或壳宽。
Notes:ypresenttotalweight, xpresenttheshellheight,shelllengthandshell.
研究普遍认为,利用生长模型预测养殖对象的生长状况,通过生长曲线的变化趋势了解养殖对象在不同发育阶段的生长潜力,实现日常养殖管理的合理安排,达到理想的养殖效果,最终能提高生产水平和经济效益[16]。但是不同的水产动物有不同的生活史及生长规律,进行生长拟合的时候机械套用模型,往往得不到与实际情况符合的拟合方程。因此,应考虑在生物生长的不同阶段用不同的生长模型来拟合,这样才能更好地建立符合实际情况的生长模型。本实验中,采用了不同的生长模型对长牡蛎壳黑F7的生长情况进行模拟,取得了比较好的拟合效果,为合理制定育种计划及生态研究提供了参考。此外,长牡蛎易受环境因素影响,在今后的研究中也需要尽量通过积累不同环境下的生物学数据,以提高生长模型的普适性来更高地指导实际生产。
猜你喜欢
科学与财富(2021年36期)2021-05-10
中学生数理化·高一版(2021年2期)2021-03-19
中学生数理化·高一版(2021年2期)2021-03-19
养猪(2020年1期)2020-02-19
今日农业(2019年11期)2019-08-15
国外畜牧学·猪与禽(2018年11期)2018-05-14
中学生数理化·高一版(2018年2期)2018-04-04
环球时报(2017-12-25)2017-12-25
农村农业农民·B版(2015年9期)2015-10-16
大众考古(2015年6期)2015-06-26
