
植物源抗冻蛋白作用机制及其在食品中的应用
2020-01-08 05:59:06扈莹莹温荣欣秦立刚
食品科学 2019年23期
于 晶,扈莹莹,温荣欣,秦立刚,陈 倩
(1.东北农业大学食品学院,黑龙江 哈尔滨 150030;2.东北农业大学动物科学技术学院,黑龙江 哈尔滨 150030)
植物源抗冻蛋白(antifreeze proteins,AFPs)是植物在外界环境发生改变时为保护细胞免受冻害而产生的一种功能性蛋白质。AFPs在不同植物体内的分布不同,它们都具有一定的抗冻活性,是一类能提高植物细胞抗冻活性的蛋白质的总称[1]。自20世纪发现以来,有关AFPs的主要研究对象是鱼类、昆虫等,如今植物源AFPs已成为研究热点。根据GB 2760—2011《食品安全国家标准食品添加剂使用标准》报道,AFPs可以作为新型食品添加剂添加到冷冻食品中[2]。
AFPs具有3 种特性,分别是热滞活性、修饰冰晶生长形态效应和抑制冰晶再结晶。植物源AFPs相对于鱼类、昆虫源AFPs热滞活性较低,但抑制冰晶再结晶能力较强[3],因此认为植物源AFPs调控抗冻性的主要途径是通过抑制胞外冰晶的生长及抑制冰的再结晶。植物源AFPs通过抑制溶液中冰晶的再结晶现象,减轻因环境温度在零度以下附近波动产生的机械损伤,抑制在冷藏过程中食品因反复冻融造成的质量劣变,比其他来源的AFPs更适合添加到冷藏食品中。本文通过综述植物源AFPs的作用机理、提取方法,以期为AFPs在食品领域的应用提供理论参考。
1 植物源AFPs的概述
1.1 AFPs的来源
植物源AFPs主要来源于寒冷、高海拔地区的植物体内,现植物源AFPs的研究对象有40余种,其中约在20 种高等植物中检测到AFPs[4],但真正被分离纯化的植物源AFPs种类较少,主要分离自冬黑麦、沙冬青、胡萝卜、女贞叶、黑麦草、苜蓿等。表1概括了不同植物源的AFPs,以及它们的存在形式和功能作用。植物源AFPs是一种适应性蛋白,除低温外,干旱[5]、脱落酸[6]、外源乙烯[7]等条件也可诱导植物产生AFPs。经研究发现植物体内AFPs在氨基酸序列、双螺旋结构、冰晶结合单元的特异性与植物表现出的抗冻特性相关。

表1 植物源AFPs来源分类及功能Table 1 Sources and functions of plant-dervied AFPs
1.2 AFPs的特性
1.2.1 热滞活性
一般溶液(如NaCl、蔗糖溶液等)的冰点是固、液两相蒸汽压平衡时的温度,因而冰点应等于熔点。由于AFPs只影响结冰过程,几乎不影响融化过程,使冰点低于熔点,其冰点和熔点的差值称为热滞差值,AFPs的这种活性称为热滞活性[16-17]。
1.2.2 修饰冰晶生长形态效应
在低温环境下,冰晶由于受到AFPs的影响,其生长形态发生改变,由正常冰晶的扁圆型生长为六角形棱锥。随AFPs的浓度增加和作用时间的延长,冰晶的形态趋近针状[18]。
1.2.3 抑制冰晶再结晶
在溶液冰点温度下,溶液中的小冰晶逐渐消失聚集成为大冰晶,对产品的组织结构造成破坏。添加AFPs可抑制溶液冰晶的再结晶现象,使小冰晶均匀分布[19]。
2 AFPs的作用机制
不同植物中的AFPs存在形式不同,并且没有共同的演化规律[20]。然而,它们均具有热滞活性、修饰冰晶生长形态和抑制冰晶再结晶功能。其作用机理为AFPs在溶液中可调节原生质溶液状态,抑制冰晶的生长活性,且在极低浓度条件下就能有效抑制再结晶,但无法降低冰点[21]。AFPs在溶液中作用方式可从能量和分子结合作用角度定义为热动力学作用原理和氢键结合作用。
2.1 热动力学作用原理
AFPs在溶液中遵循热力学定律,在作用机制中,热力学现象从宏观物质热运动角度说明反应可发生的程度,从动力学角度说明具体生长的效率。因此从宏观热动力学角度将刚体能量学作用机制、冰晶吸附抑制作用机制、热滞现象动力学作用机制概括为热动力学作用原理。
2.1.1 刚体能量学作用机制
将溶液中AFPs视为极小的粒子,可根据界面能量推测它们在冰-水界面的平衡位置,在冰晶生长过程中,粒子与晶体和水相互作用。由表面能原理可知,由于“AFPs-冰晶”与“AFPs-水”表面存在差异,水会尽可能地缩小表面积减少界面总能量。因此,当AFPs滞留在冰晶表面时,冰-水界面的面积减少使体系达到平衡。通常,水分子在冰晶表面推进AFPs向前,在溶液中形成冰晶体[22]。在温度降低到水溶液的过冷温度以下1 ℃或是更低时,会产生很大的结晶压力,但由于AFPs与冰晶结构匹配时,水分子产生的结晶压力使AFPs在冰晶表面保持平衡,使粒子固定在冰晶表面无法向前推进。由此,冰晶的生长只会发生在足以吞噬这些粒子的过冷状态下,表现为冰点降低[23]。根据开尔文效应,在弯曲冰晶表面冰点局部下降,冰晶在吸附的AFPs之间向前凸起(图1)。

图1 AFPs降低局部冰晶冰点示意图[23]Fig. 1 Schematic diagram of the role of AFPs in reducing local ice crystal freezing point[23]
从刚体力学热力学角度说明AFPs吸附在冰晶表面的原因与降低冰点的原理,需要进一步讨论AFPs吸附时间永久性、其作用效果与浓度之间的关系。
2.1.2 冰晶吸附抑制作用机制
在低温条件下,AFPs具有选择吸附性,与冰晶混合后吸附在冰晶生长的表面。冰晶在AFPs分子之间的通路与水结合,在溶液中被AFPs分子覆盖的冰晶表面停止生长,而未被覆盖的区域则沿着平面继续向前推进形成一个圆形的表面,使其表面曲率增加,具体见图2。
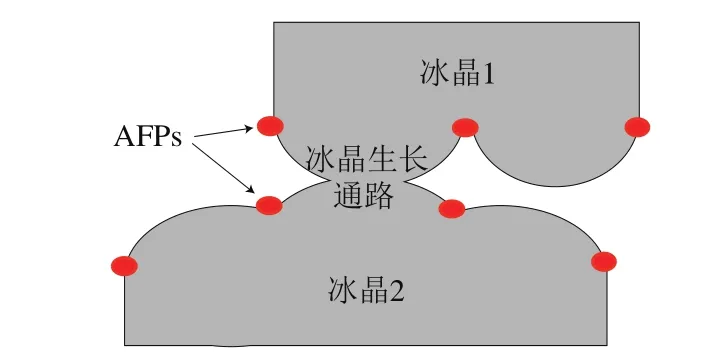
图2 AFPs吸附抑制冰晶生长示意图[24]Fig. 2 Schematic diagram of ice crystal growth inhibition by AFPs adsorption[24]
当冰晶表面积与体积比超过冰晶自发生长的热力学值时,冰晶的生长会受到抑制,从而导致冰点下降。换言之,由于AFPs吸附在冰晶生长通路上,粘滞在冰晶表面的AFPs导致冰晶表面像曲面一样弯曲。从热力学角度来说,溶剂更倾向于在平滑的冰晶表面生长,但由于冰晶表面曲率的增加使得溶剂不易附着在曲冰面上,因而冰晶的生长受抑制。其中,AFPs会一直吸附在冰晶表面,直到冰晶受到外界作用融化,方可脱离冰晶表面,且AFPs浓度越高,其作用效果越好。吸附抑制作用机制在目前AFPs研究领域中较为合理,利用吸附抑制作用机制可以计算出AFPs吸附在冰晶表面导致的冰点下降值[25]。
2.1.3 热滞现象动力学作用机制
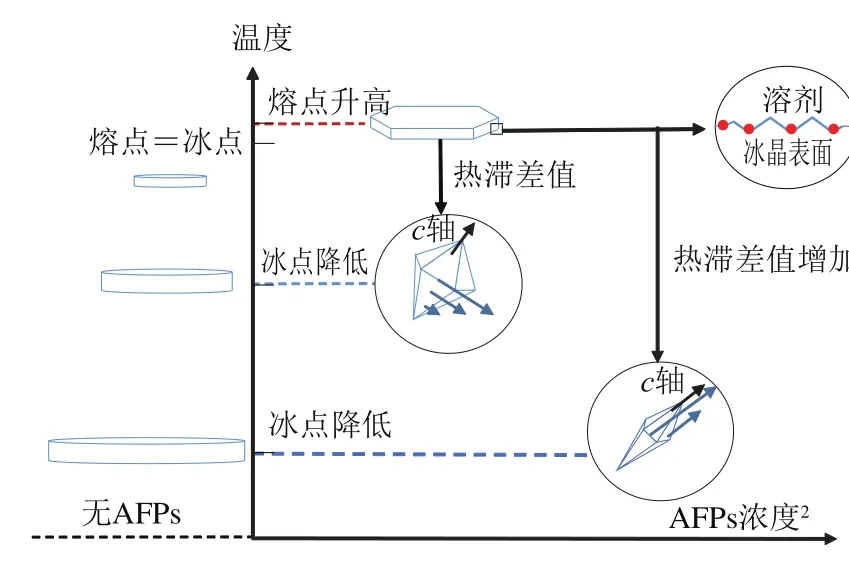
图3 热滞活性和冰晶形状和结构随浓度变化情况[28]Fig. 3 Changes in ice crystal shape and structure with thermal hysteresis activity concentration[28]
目前,许多研究已经证实热滞活性具有浓度依赖性[26-27],如图3所示,AFPs其可以与冰晶平面相互作用,吸附在冰晶表面降低冰晶表面的冰点并提高冰晶内部熔点,使冰晶产生热滞差值,而且热滞差值的增加与蛋白质浓度的平方成正比,不同类型的AFPs在等浓度下可产生的热滞差值也不同。AFPs还具有塑造冰晶表面晶体形状的能力,AFPs存在时会影响溶液中冰晶的生长结构,在AFPs作用活跃的情况下晶体延c轴方向生长;溶液中不存在AFPs的情况下,冰晶会随着温度的降低而变大,类似于扁平型圆盘生长,且冰点与熔点相等。
2.2 氢键结合作用
在溶液中AFPs与冰晶之间通过氢键进行结合,但氢键形成方式不同,主要包括“偶极子-偶极子”、氢键结合、冰核抑制、表面互补这4 种作用机制。
2.2.1 “偶极子-偶极子”冰核抑制机理
在水分子中,由于氢原子与氧原子的电荷中心并不重合,在正电荷中心与负电荷中心之间存在一段微小的距离,在电场中的原子相当于一个微观的电偶极子受到电场的作用。AFPs双螺旋结构中具有平行于其螺旋轴的偶极子[29],AFPs偶极子与冰核表面水分子偶极子的相互作用,使得水分子的无序性降低,在合向量方向上作用力最大。在低浓度的AFPs溶液中,由于冰晶表面AFPs浓度太低,导致无法进行显著的“偶极子-偶极子”相互作用,“偶极子-水”相互作用占主导地位,在冰核的棱镜面上,AFPs的螺旋轴平行于冰晶偶极子合向量方向。AFPs与冰晶表面的相互作用会诱导棱镜外层水分子局部有序排列。根据熵增原理,从c轴方向冰晶表面添加水分子比从冰晶表面添加水分子更加有利,所以冰核在c轴方向进一步增长。偶极子的相互作用改变了冰核的生长习惯,形成了双锥形冰晶体(图4A),并抑制冰核在基面的生长。随着AFPs浓度的增加,棱镜表面的“偶极子-偶极子”相互作用增强,导致棱镜表面水分子的偶极子与AFPs螺旋轴的偶极子反向平行。在不同棱镜面周围的“偶极子-偶极子”相互作用下,AFPs的螺旋轴逐渐平行于冰晶的c轴,这样改变了双锥形晶体生长方式的螺距,冰晶延c轴方向进一步生长(图4B)。当AFPs浓度足够高时,所有的AFPs螺旋轴都平行于c轴,双锥形状近乎于针状生长(图4C)。

图4 冰晶朝c轴方向生长模式[29]Fig. 4 Growth models of ice crystals in the c-axis direction[29]
“偶极子-偶极子”作用机制中AFPs与冰晶并不是以固定的方式结合,而是依靠冰核表面的水分子与AFPs的协同作用。由于AFPs螺旋轴表面散布着正电荷与负电荷[30],这些暴露的电荷对冰核表面的水分子偶极子产生了影响,这就意味着可以存在许多氢键模式,AFPs螺旋的偶极特性可能是这一结合形式的关键性因素。
2.2.2 氢原子结合作用机制
氢原子结合作用机制也称为“晶格匹配”或“氢键匹配作用机制”,在AFPs结构中存在β-链、α-转角和跨主链链接的氢键结构,该结构赋予AFPs刚性球形褶皱(图5A),AFPs中5 种含羟基氨基酸——谷氨酰胺(Gin)9、苏氨酸(Thr)18、苏氨酸(Thr)15、谷氨酰胺(Gin)44和天冬酰胺(Asn)14的羟基在主链和侧链上形成平坦的表面[31]。对溶剂结构分析表明,AFPs没有增强与水相互作用的能力,而是与冰产生线性结构的相互作用[32]。AFPs刚性平坦表面与平坦的冰晶表面之间形状互补,使AFPs表面亲水基团与冰晶表面氧原子形成氢键,多个氢键的精确匹配提供了很强的结合性和特异性,紧密连接的冰结合位点通过范德华力直接排斥干扰冰晶与水分子的进一步结合。
AFPs覆盖在冰晶的基面并与冰晶的棱镜面结合(图5B),AFPs冰结合位点不仅降低了冰晶的冻结活性,而且对冰晶的形态产生了影响,在AFPs存在下形成的冰晶为短六角形双锥体,随着时间的延长冰晶延c轴生长形成双锥体结构。

图5 氢原子结合作用机制中AFPs刚性结构与结合方式[31]Fig. 5 AFPs rigid structure and bonding mode in hydrogen atom-binding mechanism[31]
2.2.3 冰核抑制作用机制
冰核是形成冰晶的基础,没有冰核的存在,冰晶也就不能生长。AFPs具有“冰结合面”和“非冰结合面”两种不同面。其中,“冰结合面”上羟基和甲基有序间隔排列使得冰结合面上形成类冰水合层,促进冰核生成;而“非冰结合面”上存在的带电荷侧链及疏水性侧链,使得非冰结合面上的界面水无序,抑制冰核形成[33]。该作用机制在分子层面揭示了AFPs抑制冰核形成的原理。
2.2.4 “AFPs-冰晶”表面互补作用机制
冰晶与AFPs表面互补结合受多种作用力影响[34]。在“AFPs-冰晶”结合表面,冰如同“配体”与AFPs“受体”结合,因此该作用机制也被称为“受体-配体”作用机制[35]。Kuiper等[36]发现黑麦草的AFPs一级结构中含有许多重复性序列,在118 个氨基酸的多肽中,每14~15 个氨基酸组成一个环,8 个这样的环组成一个β-筒状结构,β-筒状结构的一端是保守的缬氨酸疏水核,另一端是由内部天冬酰氨构成的梯状结构。缬氨酸是疏水性氨基酸,天冬酰氨是亲水性氨基酸;β-筒状结构的亲水端与冰晶表面吻合互补,疏水端则有效地防止了水与冰晶的结合,阻止了冰晶继续生长,假定与冰晶结合区域AFPs表面平行(图6A)。在实际运用中,冰晶有许多不同的表面,每一个表面与氧原子间距都不同,冰晶就像配体一样,通过表面互补选择性地与不同受体的AFPs结合(图6B),这种表面互补方式可适用于所有AFPs[37]。AFPs的“表面互补”作用机制在解释多种冰晶表面与AFPs结合方式上提供了最合理的说明,但目前结合表面各种作用力的方式和大小还需进一步研究验证[38]。
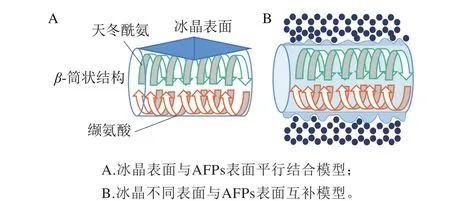
图6 AFPs与冰晶表面互补示意图[36]Fig. 6 Schematic diagram of complementation between AFPs and ice crystal surface[36]
综上所述,AFPs作用的共同特征如下:1)AFPs与冰晶表面结合取决于两个因素:一是由于热动力学活性的平衡状态使AFPs吸附在冰晶表面保持能量最低;二是由于AFPs的蛋白质结构与冰晶表面产生相互作用;2)AFPs在热动力学环境中相当于一个杂质粒子,在不存在相互作用的前提下不偏向于水或者是冰晶,由开尔文定律分析可知AFPs必须吸附在冰晶表面;3)AFPs氢键作用原理是依靠某种作用力与冰晶表面或冰核周围水分子发生作用,阻止冰晶的生长,进而降低冰点,引起热滞活性效应,同时依靠该作用力可抑制再结晶,也可以通过测定其热滞差值和冰晶结构验证该种AFPs是否存在;4)AFPs的作用能力与其浓度有关,且浓度越大作用效果越强,AFPs也存在一个渐近的活性饱和浓度,超过此浓度后,其抗冻能力不再增加,类似于酶类作用机制;5)在多种作用机制中提出AFPs有亲水作用基团和疏水作用基团,作用基团的存在与AFPs的氨基酸组成和二级结构有关。
3 植物源AFPs的分离纯化方法
3.1 渗透离心法
渗透离心法利用缓冲溶液以一定的固液比对植物进行均浆、渗透离心、过滤、离心、冻干,最终得到AFPs粉末。此外,绿色叶蛋白还需经过脱色等前处理,该种方法是提取植物源AFPs最常用的方法。尉姗姗等[39]利用纤维素DE-52离子柱层析提取分离出了冬季新疆沙冬青叶片中的AFPs,经差示扫描热量法测定分离出的蛋白组分热滞活性,结果显示当AFPs质量分数达到0.36%时热滞差值为0.46 ℃。提取后脱色采用有机溶剂或超临界CO2流体萃取,可获得高品质叶蛋白[40]。Jarząbek等[41]从木本植物针叶云杉中利用抗坏血酸真空渗透、离心收回渗透液获得AFPs,根据实验十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)显示,在这些云杉中积累了5~9 个多肽带,此植物源AFPs的分子质量为7~80 kDa。Simpson等[42]使用液氮研磨、浸提、离心的方法提取连翘树皮中的蛋白质,后利用阴离子交换、羟基磷灰石层析和凝胶溶液溶解从蛋白质中分离纯化出分子质量为20 kDa的AFPs。
3.2 冰特异性吸附分离法
冰特异性吸附分离法是利用AFPs可特异性吸附在冰晶表面的特点,利用新型的冰晶吸附设备或冰块从粗提物中纯化与冰晶结合的AFPs,该种方法可明显提高AFPs的浓度,操作简单方便。刘尚等[43]根据AFPs与冰晶的特异性结合,将碎冰放入女贞叶提取液中,通过碎冰吸附、凝胶过滤和离子交换层析,获得了4 个组分的蛋白质,其中的1 个经鉴定具有热滞活性,该蛋白亲水性氨基酸含量较高,确定为AFPs。Kuiper等[44]设计“冷手指”设备,该设备中的“冷手指”通过内部绝缘塑料管连接可编程水浴温度控制装置,让AFPs生长在“冰手指”上,结束时将“冷手指”上的冰块融化后可用于第二轮吸附,冰馏分中的AFPs可以通过冻干或超滤回收。Zhang Chao等[45]参考Kuiper等[44]的实验装置并进行了改进,利用三段提取法从小麦麸皮中提取并纯化植物源AFPs,结合冰特异性吸附原理提取纯化系数比传统非冰特异性吸附法高270 倍。
3.3 浊点萃取法
浊点萃取法利用非离子表面活化剂吸附水中悬浮的固体颗粒,加速悬浮液中粒子的沉降,实现有机物质的分离。非离子表面活性剂在水溶液中的溶解度随着温度的升高而降低,在升至一定温度时出现浑浊,可在放置或离心后得到两个液相,该温度叫浊点温度。浊点萃取法是分离科学领域一种新型的提取方法,利用表面活性材料在加热条件下,由于非离子表面活性物质在浊点温度分离成界面清晰的两相,最终实现蛋白质组分分离[46]。浊点萃取技术大多用于亲水蛋白与疏水蛋白的分离,并且扩展到植物化学物质的分离和与预浓缩[47]。Xu Huaneng等[48]将浊点萃取法用于冬小麦AFPs的分离纯化,最后得到的蛋白产品用SDS-PAGE测定其相对分子质量范围为15~30 kDa。
4 植物源AFPs在食品领域的应用
冷冻是食品冷链运输环节最重要的一个阶段,且冷藏是食品领域常用的贮藏方式,在食品的冷链运输中极易发生温度波动的现象,使食品发生冷冻-解冻循环。因此不可避免地发生冰晶再结晶现象,进而对细胞组织造成破坏,导致食品理化性质、组织形态发生变化[49]。同时,水的再结晶现象也会导致细胞内部组分发生改变,比如产生蛋白质变性、淀粉回生等现象[50]。植物源AFPs作为冷冻食品添加剂,可以有效提高冷冻食品的品质。
4.1 在冰激凌中的应用
冷冻甜乳品冰晶的形成和再结晶对于产品的品质有着至关重要的作用,冰晶的形成过程决定冷冻乳品硬化和贮存过程中冰晶的生长情况,影响冰激凌的乳脂状态、粗糙和水润程度、硬度、冰爽口感等感官品质;而再结晶决定了整个冷冻操作后冰激凌质地和结构稳定性。通过添加AFPs可从机械的角度控制冰晶冰核的大小,抑制蔗糖溶液中的冰晶生长,改善静态贮存条件下的稳定性[51]。
Lillford等[51]从冬季小麦草中提取AFPs添加到冰激凌中,通过实验研究观察到,AFPs质量分数达到0.05%~0.10%时即可诱导冰晶平均尺寸减小。Regand等[52]将冬麦草AFPs添加到冰激凌中,通过亮视野显微技术观察到,当粗蛋白添加量达到0.003%时,冬麦草AFPs表现出明显的抗冻性。对低温贮藏(-18 ℃)1 个月之后的冰激凌进行感官评价发现,温度在-18 ℃和波动条件(-20~-10 ℃)下空白组的冰激凌非常粗糙,冰激凌的冰晶成片状,而添加冬麦草AFPs的冰激凌口感滑腻、冰晶细小且均匀,证实了添加冬麦草AFPs能够修饰冰激凌冰晶的生长形态,且抑制再结晶;此外,通过热激处理进行验证,在巴氏杀菌条件下冬麦草AFPs活性没有受到影响,在添加量达到0.13%时冬麦草AFPs活性效果最好。Zhang Yanjie等[53]从冷驯化的燕麦中提取AFPs,在冰激凌中添加AFPs(添加量为0.1%)即可使玻璃化转变温度从-29.14 ℃上升到-27.74 ℃,提高了冰激凌抗融性并有效抑制了其再结晶。通过添加植物源AFPs减轻了冷冻和温度波动对冰激淋的损伤,从而获得了品质与口感优质的冷冻食品。
4.2 在冷冻面团中的应用
淀粉是多种食品加工过程中必不可少的加工原料,植物源AFPs在冷冻面团中的应用也相对较多。植物源AFPs可调节面团中结晶水析出的量,使面团中的水和面筋保持冷冻前的状态,进而使得面团中的凝胶稳定性、发酵面团的孔洞大小和均匀度、熟制后面团的质构和香气等不发生劣变[54]。
贾春利等[55]将女贞叶AFPs添加到小麦淀粉中,研究了淀粉凝胶冻融后的稳定性,结果发现随着女贞叶AFPs浓度的增加,冻融淀粉析水率和可冻结含水量显著降低,凝胶超微结构被修饰使得淀粉孔洞增大受到抑制,其均匀性提升,延缓了凝胶硬度增加和弹性降低,改善了冻融后凝胶质构。Kontogiorgos等[56-57]从冬麦草中分离得到一种热稳定性AFPs,并将其加入到面团中,发现空白面团在温度波动条件下(-20~-10 ℃)贮藏30 d,其面筋结构发生改变,而加入0.1%冬麦草AFPs的面团中冰晶再结晶现象降低,面团面筋网孔结构较小且形状均匀。此外,Liu Mei等[58]在面团中加入胡萝卜AFPs,发现添加胡萝卜AFPs降低了面团在冷冻-解冻循环过程中冻融水含量的增加,减弱了冷冻-解冻循环对冷冻面团贮藏性能和超微结构的破坏,从而改善了面团熟制后的特征体积和质构特性。夏露等[59]以冬小麦麸皮中提取的AFPs作为添加剂,分别以相对于糯米粉1%、2%、2.5%、3%的添加量制备速冻汤圆,实验结果表明2.5%的AFPs添加量对汤圆的品质有明显提高,煮熟后的汤圆外观光滑,弹性好,浑汤清澈透明。
4.3 在新鲜水果中的应用
速冻水果含水量高,在冷冻冷藏过程中易出现汁液流失、果实软烂变形等现象[60]。将海藻糖与冬小麦AFPs混合注入新鲜草莓中,然后经液氮冷冻,在解冻之后测定其细胞活性和汁液损失率,结果表明AFPs显著提高了草莓的耐冻性,并且能够保留其原有的形态和水果本身的纹理形状,保持水果的感官与风味[61]。Cruz等[62]利用真空浸渍法将豆瓣菜叶片与AFPs在封闭储罐中进行反应,经过短时间的真空压力作用和常压恢复过程,AFPs与豆瓣菜叶片毛孔直接接触,避免其了对叶片组织的损伤,降低叶片汁液损失;通过微观结构分析发现,豆瓣菜叶片中冰晶较小。新鲜的果蔬抗冻保护重点在于冷冻后的细胞形态是否完整、冻伤组织的细胞活力是否存在,而且果蔬抗冻领域不同的AFPs对于果蔬类细胞食品的适用性也需要进一步研究[63]。
5 结 语
植物体内产生AFPs是一个复杂的过程,植物源AFPs的存在与植物的生长抗性密切相关,外界环境的改变会直接影响植物源AFPs浓度,同时化学诱导等因素也会刺激植物产生AFPs,其中食品领域多采用克隆或转基因的途径获取植物源AFPs。植物源AFPs可抑制食品在冻结和冻藏过程中再结晶现象,减少解冻时汁液流造成的营养及品质的降低。此外,植物源AFPs无毒性,在多种食品中添加不会产生负面影响,其不存在毒性蛋白的特性[64],因此研究植物源AFPs在未来食品领域的应用前景非常广阔。
目前,对AFPs作用机理的研究可为食品抗冻技术提供新的研究方向,同时也为生产实践提供了理论依据。然而,植物源AFPs在其规模化应用方面仍受到较大限制,这主要是由于从植物体内提取的AFPs量少,无法进行大规模生产;而人工合成成本高,可重复性差。因此,利用植物源AFPs的自身性质,如相对分子质量、等电点、热滞活性及其抗冻作用机制开发大规模投入使用的高效分离纯化方法,并找到植物源AFPs在各类食品中应用的最佳条件,可为冷链食品生产、运输、贮藏提供保障。同时,利用基因技术,将植物体内的抗冻基因转入到受体细胞中,以期获得大量的具有抗冻活性分子,也将是我们在植物源AFPs食品应用领域的一个新方向。
猜你喜欢
现代商业银行·财富生活(2023年12期)2024-01-20 00:02:24
科普童话·学霸日记(2021年12期)2021-05-30 10:48:04
化工管理(2021年7期)2021-05-13 00:46:12
成都信息工程大学学报(2019年5期)2019-05-21 00:46:14
现代营销(创富信息版)(2018年6期)2018-09-05 03:14:32
上海金属(2016年3期)2016-11-23 05:19:38
琴童(2016年5期)2016-05-14 15:43:58
大型铸锻件(2015年1期)2016-01-12 06:32:58
上海金属(2014年3期)2014-12-19 13:09:04
上海金属(2014年3期)2014-12-19 13:09:03
