
双相体系中一种诱变株细胞催化制备(S) 奥美拉唑
2020-01-03 10:13王志勇李德鹏王繁业
青岛大学学报(工程技术版) 2020年4期
王志勇 李德鹏 王繁业
摘要: 针对传统方法制备(S) 奥美拉唑过程中存在的催化剂活性弱及底物浓度过低等问题,本文以氯仿 水双相体系为反应介质,采用R. rhodochrous诱变株细胞催化奥美拉唑硫醚的生物氧化制备(S) 奥美拉唑。使得底物及细胞浓度分别达到180 mmol/L和100 g/L,且得到较高的产率(99.29%)和对映体过量百分率(enantiomeric excess,ee%)。同时,采用响应面法(response surface methodology,RSM)优化反应条件,建立了(S) 奥美拉唑收率的二次多项式模型,该模型的相关系数(correlation coefficient,R2)达到0.9990。研究结果表明,R. rhodochrous诱变株细胞表现出高对映选择性,催化活性以及底物和产物耐受性,可以用于高效制备手性亚砜。该研究对提高(S) 奥美拉唑单位体积产率及降低生产成本具有重要意义。
关键词: 生物氧化; 氯仿 水双相系统; (S) 奥美拉唑; 静息细胞; 响应面法
中图分类号: TQ463 文献标识码: A
抗溃疡药物质子泵抑制剂(proton-pump inhibitor,PPI)属于手性亚砜,奥美拉唑是其代表性药物之一,其立体构型与药物的药理活性及代谢速率等密切相关[1] ,采用单一对映体给药,可以大幅度提高其药理活性,(S) 奥美拉唑的高效制备一直是现在面临的问题。目前,手性亚砜主要由金属配合物催化剂[2] 催化硫化物的不对称氧化制得,但存在环境不友好等许多缺陷;也有许多研究者采用酶催化制备手性亚砜,如单加氧酶[3] ,辣根过氧化物酶[4] ,肌红蛋白和细胞色素等,利用酶催化制备手性亚砜存在许多不足,例如需要辅助因子循环系统,酶的价格昂贵等,而采用全细胞催化,不需要昂贵的辅因子再生。近年来,单水相系统中硫化物的全细胞生物氧化引起了人们的广泛关注[5 7] 。R.Holt等人[8] 在单水相体系中对15株菌株进行了生物氧化奥美拉唑硫醚实验,获得ee%为17%~99%的(R) 奥美拉唑;T.Yoshida等人[9] 在单水相体系中利用真菌CunninghamellaechinulataMK40制备了ee%>99%的(S) 雷贝拉唑。但在单水相体系中,由于底物和产物对细胞有较强的抑制作用,因此底物的质量浓度维持在较低水平(0.08~1.5 g/L),而且转化率随着底物质量浓度的增加而显著降低[7] 。相较于单水相体系,采用有机溶剂 水双相体系可以有效提高底物和产物的浓度。底物和产物的高浓度对提高单位体积产率,降低生产成本具有重要意义。因此,许多科学家致力于提高底物和产物的质量浓度,以提高反应器的操作强度,降低生产成本。Liu Z Q等人[10] 采用有机溶剂 水双相系统制备依折麦布手性中间体,可以显著增加底物的质量浓度。目前,在有机溶剂 水双相系统中利用细胞催化奥美拉唑硫醚生物氧化制备(S) 奧美拉唑尚未有报道。因此,为了显著提高底物浓度和(S) 奥美拉唑的产率,本研究在有机溶剂 水双相体系中利用R. rhodochrous诱变株细胞催化奥美拉唑硫醚的生物氧化制备(S) 奥美拉唑,并采用响应面法(response surface methodology,RSM)优化不对称细胞催化生物氧化的反应条件,建立了(S) 奥美拉唑收率的二次多项式模型,以大幅度提高(S) 奥美拉唑的产率。该研究对提高(S) 奥美拉唑单位体积产率及降低生产成本具有重要意义。
1 实验部分
1.1 实验材料和仪器
(S) 奥美拉唑,苏州维他化学有限公司;奥美拉唑硫醚,济南沃尔德化工有限公司;奥美拉唑,山东寿光富康制药有限公司。其他化学制品均为分析纯。本研究使用RhodococcusrhodochrousATCC 4276诱变株细胞,该诱变株具有比野生菌株更好的生物氧化活性和底物耐受性。
UV 1000型紫外可见分光光度计,翱艺仪器(上海)有限公司;Agilent 1200 LC型高效液相色谱仪,安捷伦科技有限公司;PHS 25C PH计,杭州奥立龙仪器有限公司。
1.2 静息细胞在有机 水双相体系中催化奥美拉唑硫醚氧化形成(S) 奥美拉唑
在反应瓶中进行生物氧化反应,反应温度为34~40 ℃,时间为35~45 h,转速150 rad/min,反应瓶通入洁净空气。反应有机相为氯仿,水相为磷酸盐缓冲液(pH=6.3~8.3,葡萄糖2 g/L)。将奥美拉唑硫醚溶解在氯仿中,使最终浓度为100~200 mmol/L(32.9~65.8 g/L,以氯仿体积计)。如果以水和氯仿总体积计,浓度为10~20 mmol/L(3.29~6.58 g/L)。将溶解奥美拉唑硫醚后的氯仿溶液、磷酸盐缓冲液和R. rhodochrou诱变株的静息细胞以反应瓶体积的30%~40%加入到反应瓶中,使湿细胞质量浓度为60~100 g/L,氯仿与磷酸盐缓冲液的体积比为1∶9。取样,用离心机(6 000 rad/min,6 min)分离静息细胞、氯仿和水,所得有机相用于测定对映体纯度和转化率。
1.3 高效液相色谱分析
采用Agilent 1200 LC型高效液相色谱仪进行高效液相色谱(high-performance liquid chromatography,HPLC)分析,在波长为302 nm下,测定奥美拉唑硫醚的转化率、ee%和(S) 奥美拉唑的产率。手性柱型号为Amylose-SA(规格为250 mm×4.6 mm,填料粒径为5 μ m,厂家为日本维美希公司)。流动相为乙腈和磷酸盐缓冲液(pH=6.0),体积比为15∶85,流速为0.6 mL/min。(S) 奥美拉唑和(R) 奥美拉唑的保留时间分别为5.8 min和6.9 min。
1.4 实验设计与统计分析
根据单因素预备实验,选择3个独立变量:即反应温度(A),磷酸盐缓冲液pH值(B)和反应时间(C);选择(S) 奥美拉唑的收率(Y)为响应变量。采用Design Expert 8.0.5设计了三因素三水平试验,共计17个试验点,这些试验点中有5个为中心试验点。Box-Behnken试验设计的变量和水平如表1所示。
2 结果与讨论
2.1 R. rhodochrous诱变株的静息细胞催化奥美拉唑硫醚的生物氧化制备(S) 奥美拉唑Box-Behnken设计和RSM产量结果如表2所示。由表2可以看出,(S) 奥美拉唑的最高收率为92.13%(编号10),ee%>99%。在奥美拉唑硫醚的生物氧化过程中,R. rhodochrous诱变株的静息细胞催化反应没有检测到砜的形成。以氯仿和磷酸盐缓冲液总体积计,底物的质量浓度为5.92 g/L,该结果优于文献[9,11 14]中底物的质量浓度0.5~1.5 g/L。
本研究之所以获得了较高的产量、ee%和底物浓度,其主要原因如下:
1) R. rhodochrousATCC 4276诱变株具有较高的对映选择性、催化活性和对底物与产物的耐受性,并且没有进一步氧化亚砜产生砜,说明该菌株是制备手性亚砜的很好生物催化剂,优于文献[15]报道的许多菌株,包括Trametes species H. specie,M. isabellina,Rhizopus specie,Trichaptum specie,B. cinerea,T. virideand和E. lata。
2) 为反应瓶提供洁净空气,支持细胞催化奥美拉唑硫醚的生物氧化,实现了高质量浓度的静息细胞反应。否则,细胞的质量浓度通常不会太高,如湿细胞20~40 g/L[7] ,湿细胞70~100 g/L[16] ,冻干酵母6.6 g/L[13] 。
3) 采用氯仿磷酸盐缓冲液双相体系是显著提高底物质量浓度的最重要因素,而采用有机 水双相体系可以提高细胞对底物的耐受性和对映体选择性[9 10] 。对于有机 水双相系统,水与有机溶剂的比例不仅影响微生物细胞与底物的反应界面面积,还会影响微生物细胞,因此会对微生物的催化活性产生显著影响[7,16 17] 。通常认为有机溶剂可以显著影响酶的活性[17] ,酶的生物氧化活性与溶剂的油水分配系数(oil-water partition coefficient,P呈正相关,一般用logP表示)即随着log P的增加,生物氧化活性增加,例如异辛烷(logP=4.5)[16] 的相对活性为161%,乙酸乙酯(logP=0.68)的相对活性为33.5%,通常认为logP低的溶剂(当logP<1时),会破坏细胞膜,导致细胞活性降低[9] 。
通过单变量实验,确定磷酸盐缓冲液与氯仿的最佳体积比为9∶1。在氯仿 磷酸盐缓冲液双相系统中,奥美拉唑硫醚和产物(S) 奥美拉唑几乎全部分布在氯仿中;相反,细胞则分布在磷酸盐缓冲液中。由此可以看出,氯仿中的奥美拉唑硫醚和(S) 奥美拉唑对细胞的损害很小,因此对细胞的生物催化活性几乎没有抑制作用。在單水相系统中,可能发生细胞对底物的吸附,造成底物富集,使得只有一小部分被吸附的底物参与了生物氧化反应,而其他被吸附的底物则可能会损伤细胞,抑制其催化活性。与单水相体系相比,在有机 水双相体系中,底物几乎完全分布在有机溶剂中,因此,细胞吸附的底物显著减少,吸附的底物主要参与生物氧化,而不会损害细胞和抑制其生物催化活性。
2.2 RSM优化R. rhodochrous诱变株细胞催化的奥美拉唑硫醚的不对称生物氧化
通过RSM设计和分析自变量对实验结果的影响,确定最佳实验条件,探索实验变量之间的相互作用并建立最佳拟合模型。给出(S) 奥美拉唑的产率回归模型为
Y=91.76+0.57A+0.96B+4.91C+0.63AB-1.23AC-1.09BC-10.15A 2 -10.49B 2 -4.15C 2 (1)
式中,A为反应温度的水平;B为磷酸盐缓冲液pH值的水平;C为反应时间的水平;AB为反应温度的水平和磷酸盐缓冲液pH值的水平的乘积;AC为反应温度的水平和反应时间的水平的乘积;BC为磷酸盐缓冲液pH值的水平和反应时间的水平的乘积。当式(1)中变量系数为正时,(S) 奥美拉唑的产量与该变量成正相关,线性项A,B,C和交互作用项AB具有正系数,表明这些变量的增加将提高(S) 奥美拉唑的产率。
对拟合模型进行方差分析,计算了拟合模型和模型中含有A,B,C等各单项式(如:0.57A,-1.09BC)在表2中的17个试验点水平下的平方和、自由度和均方根等。拟合模型的方差分析结果如表3所示。由表3可以看出,(S) 奥美拉唑的产率基于R2=0.999 0,说明该模型很好的拟合了实验数据。校正相关系数(adjusted correlation coefficient)R2Adj 为0.997 6,与R2仅相差0.001 4,进一步证明该模型很好的拟合了实验数据。该模型的P(P value)<0.000 1,证明了该模型的可靠性。
由式(1)和表3可知,除了相互作用项AB(P=0.022 9)外,所有线性,相互作用项的P<0.000 1~0.007 3,表明它们对(S) 奥美拉唑产率有较大影响。值得注意的是,反应温度(A),缓冲液pH和反应时间(相互作用项BC)对(S) 奥美拉唑产率的影响更大。
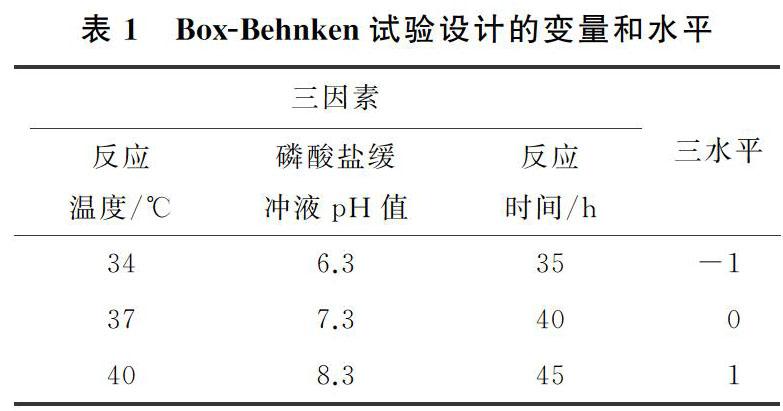
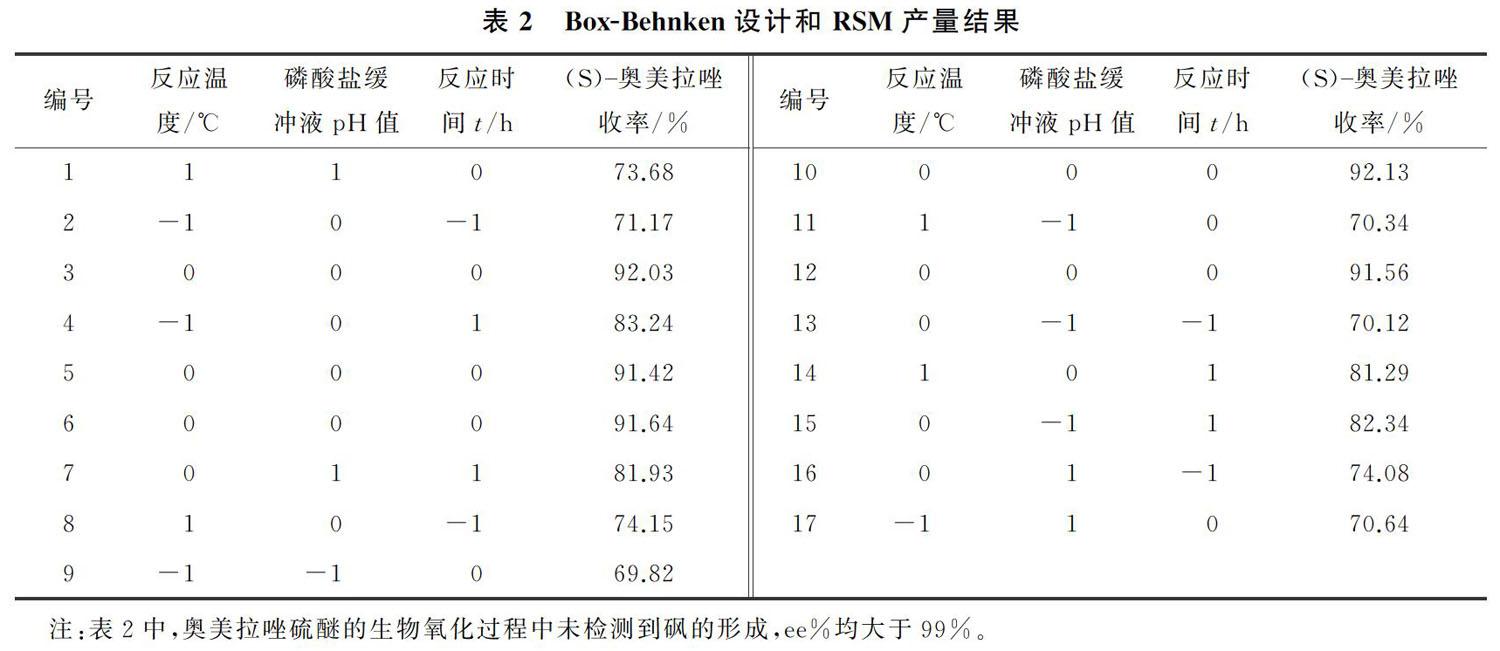
反应温度和磷酸盐缓冲液pH值对(S) 奥美拉唑产率的影响的响应面图如图1所示;反应温度和反应时间对(S) 奥美拉唑产率的影响的响应面图如图2所示。由图1和图2可以看出,当反应温度从34 ℃升至约37 ℃,(S) 奥美拉唑的产率随着反应温度的增加而增加;而当反应温度从37 ℃进一步升高,(S) 奥美拉唑的产率随着的温度的升高而降低,表明R. rhodochrous诱变株细胞催化反应的最佳温度为37 ℃,当反应温度偏离该最佳点时,R. rhodochrous诱变株细胞的生物催化活性将降低。通常情况下,生物催化剂对温度非常敏感,因此反应温度对酶、细胞的生物催化活性和稳定性具有显著影响[18 20] 。
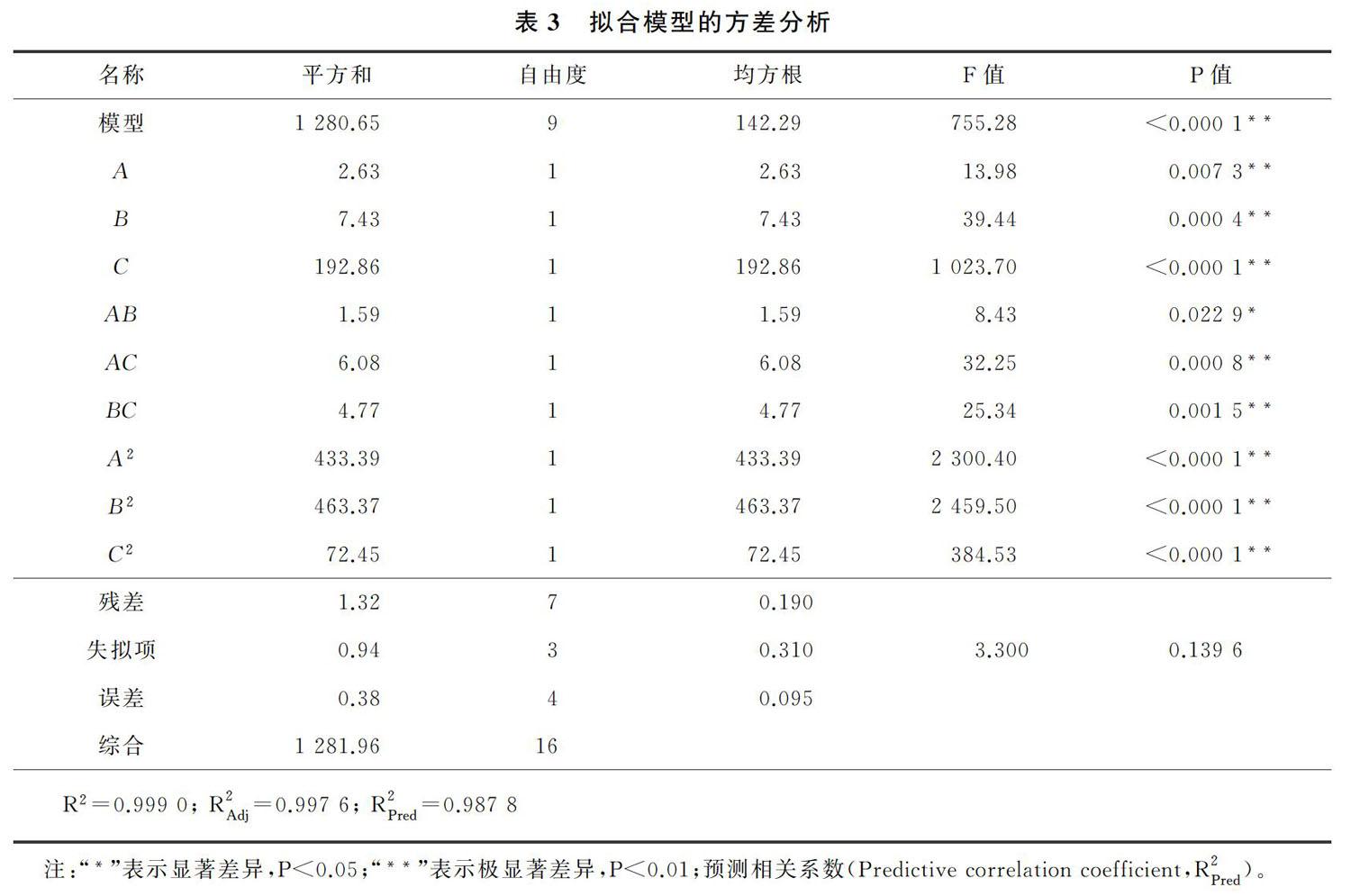
磷酸鹽缓冲液pH值和反应时间对(S) 奥美拉唑产率的影响的响应面图如图3所示。由图1和图3可以看出,随着缓冲液pH值从6.3升高至7.3,(S) 奥美拉唑的产率也随之升高;但是随着缓冲液pH值从 7.3进一步升高至8.3,(S) 奥美拉唑的产率降低。pH会影响底物和酶的离子状态,从而影响酶和整个细胞催化产物的产量和ee%[14] ,因此缓冲液的pH会影响R. rhodochrous细胞的生物催化活性;由图2和图3可以看出,随着反应时间从35 h延长至约43 h,(S) 奥美拉唑的产率迅速增加;而当反应时间从43 h延长至45 h,收率几乎没有提高。因此,最佳反应时间约为43 h,如果反应时间进一步延长,收率增加很小,而时间成本会大幅增加。
2.3 验证R.rhodochrous诱变株的静息细胞催化奥美拉唑硫醚的生物氧化的最优条件
用模型方程(1)进行优化计算,确定不对称生物氧化的最佳条件,并进行实验验证。确定的最佳参数如下:反应温度为37 ℃,磷酸盐缓冲液的pH值为7.3,反应时间为43 h。在静息细胞质量浓度为100 g/L和奥美拉唑硫醚的物质的量浓度为180 mmol/L的条件下,进行3次重复验证实验,所得(S) 奥美拉唑的平均收率为92.9%,该结果与模型计算结果(93.2%)非常吻合,相应的ee%>99%。实验结果表明,在异辛烷 水双相系统中,R. rhodochrousis诱变株细胞能够高效催化奥美拉唑硫醚的不对称生物氧化,可以用于高效制备手性亚砜。
3 结束语
本文通过对氯仿 水双相系统中诱变株细胞催化奥美拉唑硫醚生物氧化制备(S) 奥美拉唑进行研究。在高底物物质的量浓度(180 mmol/L)和细胞质量浓度(100 g/L)下,采用RSM对实验条件进行优化,获得的(S) 奥美拉唑的最佳收率为92.9%,ee%>99%;最佳条件:反应温度为37 ℃,磷酸盐缓冲液的pH值为7.3,反应时间为43 h;没有检测到砜产物形成。同时,建立了(S) 奥美拉唑收率的二次多项式模型,该模型的R2 =0.9990,可以很好的预测实验数据。实验表明氯仿 水双相系统可以显著提高细胞对底物的耐受性,因为几乎所有底物都分布在有机相中,不会损坏和抑制细胞的催化作用。R. rhodochrousATCC 4276诱变株显示出高对映选择性、催化活性以及底物和产物的耐受性,可以用于高效制备手性亚砜。该研究对提高(S) 奥美拉唑单位体积产率及降低生产成本具有重要意义。
参考文献:
[1] 刘文强, 李莉. 手性药物及其中间体拆分方法的研究进展[J]. 药学学报, 2018, 53(1): 37 46.
[2] Zhang J, Jiang T, Mai Y, et al. Selective catalytic oxidation of sulfides to sulfoxides or sulfones over amorphous Nb2O5/AC catalysts in aqueous phase at room temperature[J]. Catalysis Communications, 2019, 127(1): 10 14.
[3] 李仁结, 张龑, 许建和, 等. CYP116B家族单加氧酶的发现、表征及分子改造研究进展[J]. 生物加工过程, 2019, 17(1): 29 35, 43.
[4] 姜标, 黄浩, 罗军, 等. 硫化物的生物氧化成手性亚砜[J]. 有机化学, 2005, 25(12): 1542 1547.
[5] Li A T, Yu H L, Pan J, et al. Resolution of racemic sulfoxides with high productivity and enantioselectivity by a Rhodococcus sp. strain as an alternative to biooxidation of prochiral sulfides for efficient production of enantiopure sulfoxides[J]. Bioresour Technol, 2011, 102(2): 1537 1542.
[6] Elkin A A, Kylosova T I, Grishko V V, et al. Enantioselective oxidation of sulfides to sulfoxides by gordonia terrae IEGM 136 and rhodococcus rhodochrous IEGM 66[J]. Journal of Molecular Catalysis B: Enzymatic, 2013, 89(1): 82 85.
[7] Li A T, Zhang J D, Yu H L, et al. Significantly improved asymmetric oxidation of sulfide with resting cells of rhodococcus sp. in a biphasic system[J]. Process Biochemistry, 2011, 46(3): 689 694.
[8] Holt R, Lindberg P, Reeve C, et al. Preparation of pharmaceutically active compounds by biooxidation: U. S. 5, 840, 552[P]. 1998 11 24.
[9] Yoshida T, Kito M, Tsujii M, et al. Microbial synthesis of a proton pump inhibitor by enantioselective oxidation of a sulfide into its corresponding sulfoxide by cunninghamella echinulata MK40[J]. Biotechnology Letters, 2001, 23(15): 1217 1222.
[10] Liu Z Q, Dong S C, Yin H H, et al. Enzymatic synthesis of an ezetimibe intermediate using carbonyl reductase coupled with glucose dehydrogenase in an aqueous-organic solvent system[J]. Bioresource Technology, 2017, 229(1): 26 32.
[11] El′Kin A A, Grishko V V, Ivshina I B. Oxidative biotransformation of thioanisole by rhodococcus rhodochrous IEGM 66 cells[J]. Applied Biochemistry & Microbiology, 2010, 46(6): 586 591.
[12] Tarasova E V, Grishko V V, Ivshina I B, et al. Cell adaptations of rhodococcus rhodochrous IEGM 66 to betulinbiotransformation[J]. Process Biochemistry, 2017, 52: 1 9.
[13] Aguirre-Pranzoni C, Bisogno F R, Orden A A, et al. Lyophilized rhodotorula yeast as all-in-one redox biocatalyst: Access to enantiopure building blocks by simple chemoenzymatic one-pot procedures[J]. Journal of Molecular Catalysis B Enzymatic, 2015, 114(1): 19 24.
[14] Grishko V V, Tarasova E V, Ivshina I B. Biotransformation of betulin to betulone by growing and resting cells of the actinobacterium rhodococcus rhodochrous IEGM 66[J]. Process Biochemistry, 2013, 48(11): 1640 1644.
[15] Gonzalo G D, Mascotti M L, Orden A A, et al. Aspergillus genus as a source of new catalysts for sulfide oxidation[J]. Journal of Molecular Catalysis B: Enzymatic, 2012, 82(1): 32 36.
[16] He Y C, Ma C L, Yang Z X, et al. Highly enantioselective oxidation of phenyl methyl sulfide and its derivatives into optically pure (S)-sulfoxides with rhodococcus sp. CCZU10 1 in an n-octane-water biphasic system[J]. Applied Microbiology & Biotechnology, 2013, 97(24): 10329 10337.
[17] He Y C, Zhou Q, Ma C L, et al. Biosynthesis of benzoylformic acid from benzoyl cyanide by a newly isolated Rhodococcus sp. CCZU10 1 in toluene-water biphasic system[J]. Bioresource Technology, 2012, 115(1): 88 95.
[18] Waghmare G V, Chatterji A, Rathod V K. Kinetics of enzymatic synthesis of cinnamyl butyrate by immobilized lipase[J]. Applied Biochemistry and Biotechnology, 2017, 183(3): 792 806.
[19] De Miranda A S, Miranda L S M, De Souza R O M A. Lipases: Valuable catalysts for dynamic kinetic resolutions[J]. Biotechnology Advances, 2015, 33(5): 372 393.
[20] Mathpati A C, Badgujar K C, Bhanage B M. Kinetic modeling and docking study of immobilized lipase catalyzed synthesis of furfurylacetate[J]. Enzyme & Microbial Technology, 2016, 84(1): 1 10.
Preparation of (S)-Omeprazole by Biological Oxidation of Mutagenic Strains in Biphasic System
WANG Zhiyong, LI Depeng, WANG Fanye
(School of Chemical Engineering, Qingdao University of Science and Technology, Qingdao 266042, China)
Abstract: Aiming at the problems of weak catalyst activity and low substrate concentration in the process of preparing (S)-omeprazole by traditional methods, this paper uses the chloroform-water two-phase system as the reaction medium and uses the R. rhodochrous mutant strain Cells catalyze the biological oxidation of omeprazole sulfide to prepare (S)-omeprazole. The substrate and cell concentration reached 180 mmol/L and 100 g/L, respectively, and a higher yield (99.29%) and enantiomeric excess (ee%) were obtained. Response surface methodology (RSM) was used to optimize the reaction conditions, and a quadratic polynomial model of (S)-omeprazole yield was established. The correlation coefficient (R2) of the model reached 0.9990. The results of the study showed that R. rhodochrousmutagen strain cells exhibited high enantioselectivity, catalytic activity, substrate and product tolerance, and could be used to efficiently prepare chiral sulfoxides. This research is of great significance for increasing the yield per unit volume of (S)-omeprazole and reducing production costs.
Key words: biological oxidation; chloroform-water two-phase system; (S)-omeprazole; resting cells; response surface methodology
猜你喜欢
健康体检与管理(2022年4期)2022-05-13
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年7期)2022-04-22
中国典型病例大全(2022年7期)2022-04-22
中国药学药品知识仓库(2021年18期)2021-02-28
科学与财富(2017年2期)2017-03-15
湖北农业科学(2016年20期)2017-02-15
湖北农业科学(2016年20期)2017-02-15
现代农业研究(2016年11期)2017-01-10
科技视界(2016年21期)2016-10-17
