
冻融前后姜曲海猪精子差异蛋白质组分析
2020-01-02 06:49:10蒋春茂武彩红邱树磊陈晓兰
上海农业学报 2019年6期
张 斌,蒋春茂,武彩红*,邱树磊,陈晓兰,姚 静,李 玲,刘 莉
(1江苏农牧科技职业学院,泰州225300;2无锡市农业委员会农业信息服务中心,无锡214000)
姜曲海猪具有适应力强、耐粗饲、肉品好和产仔多等特点[1],已被列为我国优良的地方猪种。但随着市场化深入发展,我国大量引入外来猪种,加之对姜曲海猪缺乏有效保护,导致其品种面临退化,数量急剧下降,因此,姜曲海猪种质资源保护逐渐受到关注。精子冷冻保存是优良猪种种质资源保护的有效方法之一,但猪精子对低温尤其敏感,冷冻-解冻后精子存在活力差、受胎低、产仔少等问题[2-3]。目前,许多学者的研究集中于猪精液冷冻保存方案的筛选,但因冷冻损伤机理不明,致使猪精液冷冻保存方案仍未取得显著进展。
双向凝胶电泳(2-DE)是蛋白质组学研究的一个重要手段,是根据等电点和相对分子量的不同而分离蛋白质的技术[4-5]。本研究运用2-DE技术对冻融前后姜曲海猪精子的差异表达蛋白进行分离,同时,对其进行质谱鉴定和功能分析,从蛋白水平角度揭示猪精子冷冻损伤机理,为优化姜曲海猪精液超低温冷冻保存方案奠定基础。
1 材料与方法
1.1 精液采集
利用假阴道法于无菌条件下分别采集4只健康、性成熟的姜曲海公猪精液,将采集的精液充分混匀,加入1.5倍预温稀释液(德国米尼图MII稀释剂按说明书配制)混匀,置于保温桶中于1 h内带回实验室。评定精子形态完整率和活率,分别达到80%和70%以上时可用于试验。
1.2 精液冷冻与解冻
1.2.1 精液冷冻
稀释后精液于17℃过夜、离心(800g,12 min),弃上清后加入冷冻I液(海藻糖375 mmol/L,葡萄糖1.1 g,柠檬酸钠1.48 g,Tris 2.42 g,青霉素0.06 g,链霉素0.1 g,20%卵黄,蒸馏水定容至100 mL),4℃平衡1.5 h后加入等体积4℃预冷的冷冻II液(冷冻I液中添加甘油,使其体积终浓度为3%),4℃继续平衡45 min,分装于0.25 mL细管内,置于距液氮面5 cm高度处熏蒸预冷10 min,然后投入液氮保存至少一周。
1.2.2 精子解冻
从液氮中迅速取出细管,投入37℃水浴中并轻轻充分晃动以使其迅速解冻。解冻后的精液收集于10 mL试管内,用9倍体积的PBS液对其进行清洗。
1.3 蛋白质提取及含量测定
采用改进的热Trizol法提取猪精子蛋白[6]。精液经核酸酶处理后,加入预冷的三氯醋酸-丙酮溶液作用2 h,离心、弃上清,再加入预冷的丙酮作用30 min洗涤沉淀,反复洗涤3次;去除丙酮,加入裂解液500μL(2 mol/L硫脲,7 mol/L尿素,20 mmol/L Tris-HCL,2%(M/V)丙基硫酸盐(CHAPS),2 mmol/L乙二胺四乙酸(EDTA),1 mmol/L苯甲磺酰氟(PMSF),10 mmol/L二硫苏糖醇(DTT)),冰浴超声 5 min;离心(15℃,20 000g,30 min),收集上清,利用2-D Quant-kit试剂盒(GE公司,美国)进行精子蛋白含量测定。
1.4 双向电泳
1.4.1 第一向等点聚焦
采用18 cm长、pH 3—10的干性胶条,取200μg蛋白提取液与水化缓冲液[2 mol/L硫脲,7 mol/L尿素,50 mmol/L DTT,4%CHAPS,0.8%胶条缓冲液(IPG Buffer,痕量溴酚蓝)]相混合至总体积为300μL,覆盖3 mL矿物油后进行等电聚焦。
等电聚焦结束后取出胶条,分别置于平衡A液[2%SDS,6 mol/L尿素,20%甘油,0.375 mol/L Tri-HCl(pH 8.8),1%DTT]、B液[以4%碘乙酰胺(IAM)代替 1%DTT]中,于低速摇床上各平衡 15 min。
1.4.2 第二向SDS-PAG电泳
经平衡后的胶条移至12%聚丙烯酰胺凝胶处,排除气泡使二者接触紧密,在胶条一侧加蛋白质Marker,用0.5%琼脂糖封固胶条,待溴酚蓝指示剂迁移至胶底,结束电泳。
1.4.3 凝胶染色及胶图分析
凝胶参照Zhao等[7]方法进行硝酸银染色。银染后的凝胶图经软件(ImageMaster 2D platinum 5.0)分析,再将同一样品的3张胶图拟合成1张虚拟胶图进行定量分析。
1.5 质谱鉴定和分析
采用MALDITof/Tof质谱仪检测3倍以上的蛋白差异点,进行质谱鉴定分析,获得肽质量指纹图谱(PMF),再利用Mascot软件在线搜索NCBInr数据库鉴定差异蛋白。
1.6 生物信息学分析
应用blast2 go软件进行搜索,并进行蛋白质GO注释分析和GO注释富集分析。其中,“test”为不同精子差异蛋白质,“reference”为鉴定所获得的全部蛋白质,相应的P值则利用超几何概率分布计算获得,最后选择GO层次汇总。分别对3个生物学功能进行分析,包括BP(Biological process,生物过程)、CC(Cellular component,细胞组分)和 MF(Molecular function,分子功能)。
2 结果与分析
2.1 冻融前后猪精子差异蛋白表达分析
精子蛋白浓度与吸光度值呈线性关系,y=-215.5x+121.3(R2=0.995),符合样品蛋白质量控制标准,说明提取的精子蛋白浓度稳定,可用作双向电泳分离。姜曲海公猪新鲜精子蛋白含量为18.3μg/μL,冻融后其蛋白含量为15.6μg/μL。冻融前后姜曲海猪精子双向电泳图谱见图1和图2。比较分析蛋白图谱发现,有42个蛋白点存在差异,其中3倍以上差异点31个。

图1 新鲜姜曲海猪精子蛋白2-DE图谱Fig.1 2-DE map of sperm proteins fromfresh Jiangquhai boar semen

图2 冻融姜曲海猪精子蛋白2-DE图谱Fig.2 2-DE map of sperm proteins from frozenthawed Jiangquhai boar semen
2.2 差异蛋白点的鉴定及功能分类
蛋白差异点经MALDI Tof/Tof质谱仪鉴定分析,所获结果提交至Mascot软件进行在线搜索,在NCBInr蛋白数据库中搜索,分析获得有意义差异蛋白7个(表1),均下调表达,P<0.05;7个差异蛋白中,包括能量代谢相关蛋白3个,结构相关蛋白2个,受精相关蛋白和酶相关蛋白各1个。

表1 MALDITof/Tof鉴定姜曲海猪精子冻融前后差异表达蛋白Table 1 MALDITof/Tof identification of differential sperm proteins between fresh and frozen-thawed Jiangquhai boar semen
采用Blast2 go软件进行GO功能富集分析,结果表明,冻融前后姜曲海猪精子差异表达蛋白质所处的细胞组分为细胞、细胞器、超分子纤维、膜和细胞连接等;其执行的分子功能比较广泛,包括装配、催化、结构分子活性、调节分子功能、抗氧化和转运等;参与的生物学过程包括生长、发育、繁殖、免疫、解毒、运输、代谢等(图3)。
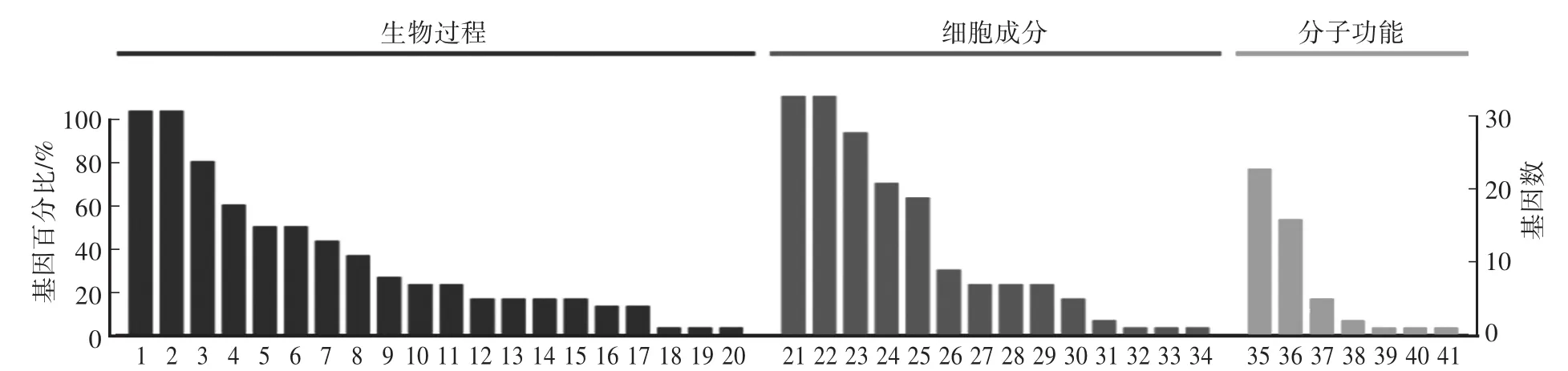
图3 冻融前后姜曲海猪精子差异表达蛋白GO功能注释Fig.3 Gene ontology analysis of differentially expressed Jiangquhai boar sperm proteins before and after freezing
3 讨论
3.1 冻融前后猪精子能量代谢蛋白差异表达
在鉴定获得有意义的差异表达蛋白中,能量代谢蛋白差异表达比例最高,包括电压依赖性阴离子通道蛋白2(VDAC2)、烯醇酶(ENO)和二氢硫辛酸脱氢酶(PDC3)。
VDAC存在于真核生物的线粒体外膜。哺乳动物VDAC包括VDAC1、VDAC2和VDAC3等3种不同的蛋白亚型[8]。其中,VDAC2存在于精子头部顶体区域[9],参与精子发生能量代谢调控[10]。王增军等[11]研究表明,人精子膜表面镶嵌的VDAC蛋白质调控精子膜内外的离子转运和信号转导,对精子的运动和功能具有重要作用。Sampson等[12]研究表明,位于线粒体外膜上的VDAC2,参与调节线粒体Ca2+的内流与外流,揭示了VDAC2在维持精子正常活动能力中的作用。烯醇酶(ENO)在催化糖酵解反应、参与基因转录、调控细胞凋亡及细胞分化等过程中发挥积极作用[13],但作用机制尚未清楚。Force等[14]研究发现,成熟精子的烯醇酶含量丰富。李鸿岩等[15]研究表明,冻融后绵羊精子ENO表达下调,且ENO可能与冷冻造成的精子活力的改变有关。PDC3是组成丙酮酸脱氢酶复合体(PDC)的3个亚单位之一。而PDC是一组限速酶,是线粒体内将丙酮酸转换成乙酰辅酶A的重要酶,且将糖酵解与三羧酸循环以及ATP的生产紧密联系在一起,在细胞线粒体呼吸链能量代谢中起着重要的作用[16]。本研究中,冻融姜曲海猪精子VDAC2、ENO和PDC3表达均下调,推测其不能为精子提供足够的运动所需能量,从而导致精子的活力和受精能力降低。
3.2 冻融前后猪精子结构蛋白差异表达
在鉴定获得有意义的差异表达蛋白中,第二大类别为精子结构蛋白,分别为β-微管蛋白和外致密纤维蛋白 2(Outer dense fiber protein 2,ODF2)。
β-微管蛋白与α-微管蛋白是构成细胞骨架的重要成分,发挥固定和支撑细胞形态的功能,维持细胞基本结构,负责轴丝迁移和鞭毛运动,参与物质运输[17]。本研究发现,冻融后姜曲海猪精子β-微管蛋白下调表达,可能是冻融过程诱发微管蛋白发生解聚,导致精子结构损伤,精子存活率降低。张欣宗等[18]研究发现,低温冷冻使人精子的微管蛋白上调表达,与本研究结果不一致,其原因有待进一步研究。
ODF2存在于精子尾部,具有保持精子鞭毛的弹性、参与精子鞭毛构成及能量代谢的功能,进而保证精子的正常运动与受精[19]。Mariappa等[20]研究表明,ODF2与精子形态、受精及受精卵的发育密切相关。ODF2也是精子中心体、体细胞中心体和纺锤体的组成部分[21]。因此,本研究中,冻融后姜曲海猪精子ODF2表达下调,一方面可能导致精子活力降低或结构畸形而造成受精失败,另一方面也可能影响受精卵纺锤体形成而使卵裂失败。
3.3 冻融前后猪精子受精相关差异表达蛋白
顶体素结合蛋白(ACRBP)是一种高度保守的哺乳动物蛋白,位于精子头部,具有与精子前顶体结合和包装组合顶体素酶原的功能,与顶体反应有关[22]。Arcelay等[23]研究发现,ACRBP均匀分布于未获能精子头部顶体内,而精子获能后,ACRBP分布在精子顶体膜上或以颗粒的形式散在顶体内。可见,精子头部ACRBP的分布与精子是否获能有关。研究表明,低温冷冻精子发生了获能反应,顶体蛋白酶原ACRBP发生了酪氨酸磷酸化[24]。本研究表明,冻融后姜曲海猪精子ACRBP表达下调,可能是冷冻导致精子提前获能所致。
3.4 冻融前后猪精子酶相关蛋白差异表达
谷胱甘肽过氧化物酶(GSH-Px)特异催化还原型谷胱甘肽(GSH)对过氧化氢的还原反应,具有清除脂质过氧化物、保护细胞膜结构和功能完整的作用[25]。GSH-Px同超氧化物歧化酶(SOD)和过氧化氢酶(CAT)构成精子防御活性氧(ROS)的主要酶系,其浓度与 ROS清除效率有关[26]。王艳华等[27]研究表明,山羊精液冷冻过程中抗氧化酶活性降低与精子细胞膜完整性损伤密切相关。本研究中,冻融精子GSH-Px表达量降低,可能导致其抗氧化活性降低,清除ROS效率降低,精子质膜的损伤程度增加。
4 结论
本研究表明,冻融后精子蛋白质总含量降低,鉴定获得的有意义差异蛋白涉及能量代谢、细胞结构装配、受精和抗氧化等生物过程。冻融影响精子能量代谢、细胞骨架、抗氧化活性和受精能力,但差异蛋白质下调如何影响精子功能和受精胚胎发育,还需进一步深入研究。
猜你喜欢
橡胶科技(2023年2期)2023-04-06 03:46:14
中国设备工程(2021年22期)2021-12-20 06:39:46
中国建筑金属结构(2021年8期)2021-09-05 01:19:14
现代畜牧科技(2021年3期)2021-07-21 08:41:58
现代畜牧科技(2021年6期)2021-07-16 05:50:06
商品与质量(2021年7期)2021-04-02 12:40:34
中国计划生育学杂志(2017年3期)2017-06-01 12:10:12
水利科技与经济(2017年2期)2017-04-22 02:34:14
兽医导刊(2016年6期)2016-05-17 03:50:17
中国现代医学杂志(2015年26期)2015-12-23 11:04:22
