
消化道巨噬细胞的功能与炎性肠病和肠道肿瘤的相关性研究进展
2019-12-23 08:00:22王田田高琛琛李利生徐敬东
首都医科大学学报 2019年6期
王田田 高琛琛 李利生 徐敬东*
(1.首都医科大学基础医学院生理学与病理生理学系,北京 100069;2.首都医科大学基础医学院机能学实验中心,北京 100069)
巨噬细胞(macrophages,Mφ)是由血液中的单核细胞(monocytes,Mo)分化而来,在人体防御功能中扮演了重要角色,消化道具有重要而复杂的免疫功能,其中Mφ在维持肠道免疫平衡中的作用引起了高度关注。此外,Mφ在不同的肠道环境下发挥多种功能,即宿主防御、伤口愈合和免疫调节。在不同的肠道微环境的作用下Mφ可分化为M1Mφ、M2Mφ和调节性Mφ(regulatory macrophages,Mregs),并且与炎性肠病(inflammatory bowel disease,IBD)、胃肠道肿瘤的关系密切。IBD包括两种类型的疾病:溃疡性结肠炎(ulcerative colitis, UC)和克罗恩病 (Crohn’s disease, CD)。IBD一直被认为是典型的自身免疫性疾病,有报道[1]证实多种因素可使肠道菌群紊乱并破坏黏液层,导致IBD的发生。目前有研究[2-3]表明肠道Mφ功能失衡可引起IBD和胃肠道肿瘤等多种肠道疾患发生。故本综述以Mφ在肠道免疫调节中的作用为主线,以肠道自身免疫疾病发生与Mφ的相关性为主要内容,阐述Mφ在肠道免疫平衡中的作用,为临床治疗肠道疾患提供新视角。
1 巨噬细胞与肠道微生物
肠道中寄居着种类繁多数量庞大的微生物,寄居部位不同,其种类和数量不同。 从口腔进入胃的细菌绝大多数被胃酸杀灭,剩下的主要是革兰氏阳性需氧菌;小肠菌群的构成介于胃和结肠之间,近端小肠的菌丛与胃内相近,但常能分离出大肠杆菌和厌氧菌;远段回肠厌氧菌的数量开始超过需氧菌,其中大肠杆菌恒定存在,厌氧菌如双歧杆菌属、类杆菌属、梭状芽胞杆菌属都有相当数量;在回盲瓣的远侧,细菌数量急剧上升,结肠细菌数量高达1011~1012CFU/mL,细菌总量几乎占粪便干重的1/3。其中厌氧菌达需氧菌的103~104倍。主要菌种为粪杆菌属、双歧杆菌属和真杆菌属,在十二指肠约为102CFU/mL,在空肠约为102CFU/mL,在近端回肠约为103CFU/mL,在末端回肠约为107~108CFU/mL,在结肠中数量最多约为1011~1012CFU/mL[3]。在生理条件下,肠道微生物在维持肠道免疫系统稳态中不可或缺,肠道微生物分为有益菌和有害菌,正常情况下它们保持动态平衡,当发生炎性反应时肠道菌群失调,刺激肠道Mφ迁徙与分化。Mφ是一种吞噬细胞,参与了固有免疫和获得性免疫,通过对细胞残片及病原体的识别、吞噬处理达到抗原呈递的效果。由于肠道是Mφ和共生菌的最大存储库,因此Mφ可以被视为肠道菌群的“监管者”而不是炎性反应催化剂。因为Mφ产生的免疫抑制细胞因子可抑制肠道细菌引起的炎性反应级联反应,由此保护肠免受菌群失衡引起的过度反应而导致严重的炎性损伤。
2 巨噬细胞的分化与生物学特征
2.1 巨噬细胞的起源和分化
单核-巨噬细胞系统(mononeuclear phagocyte system,MPS)来自于骨髓造血干细胞和胚胎中的卵黄囊祖细胞以及早期的胎儿肝脏。早在1980年,Johnson等[4]利用Chédiak-Higashi 标志物实验证实,肺的间质和肺泡内Mφ都来源于骨髓前体细胞。MPS由Mo、Mφ、破骨细胞和树突状细胞(dendritic cell,DC)等组成。
Mφ几乎存在于所有组织中。它们与循环外周血Mo(peripheral blood mononuclear cells,PBMcs)有区别,后者在稳定状态或响应炎性反应时迁移到组织中,PBMcs由骨髓中常见的祖细胞分化而来,这些祖细胞是许多不同细胞类型的前体细胞,包括中性粒细胞(neutrophil,NE)、嗜酸性粒细胞、嗜碱性粒细胞、Mφ、DC和肥大细胞(mast cell,MC)。在Mo发育期间,骨髓祖细胞又称粒细胞/巨噬细胞集落形成单位granulocyte macrophage colony forming unit,GM-CFUc)依次产生原始Mo、幼稚Mo和最终从骨髓释放到血液中的Mo。Mo从血液迁移到组织中以补充胃肠道的Mφ和肝脏的枯否细胞等[5]。
在血液中,关于Mo群体是否会分化为组织特定的Mφ存在很大争议。虽然Mo异质性尚未完全了解,但一种理论认为,Mo在血液中继续发育并成熟,在此过程中可迁徙至不同的组织中形成具有不同功能和结构的细胞群。依据它们从血液中迁移到组织所发挥的功能可分为“炎性”和“定居”Mo[6]。定居Mo存在于非炎性组织中且半衰期长,而其前体炎性Mo存在于炎性组织中且存活时间较短。这两种Mo群体可凭借细胞表面标志物趋化因子受体CX3CR1的表达来区分。炎性Mo被定义为CC-趋化因子受体2(CCR2+)CX3CR1low和GR1+(也称为Ly6C);而定居Mo定义为CCR2-CX3CR1hiGR1-。人的Mo也可根据细胞表面标志物CX3CR1的表达分为两大类[6-7],大多数Mo是CD14+CD16-CD64+CCR2+CX3CR1low,被称为“经典”Mo;或者是CD14lowCD16+CD64-CX3CR1hi,被称为“非经典”Mo。大约90%的人类Mo表达经典标记。有研究[6]显示这些定居Mo在血管受到损伤和感染的情况下迅速侵入组织,随后启动固有免疫反应并分化成Mφ。而炎性Mo的作用相反,它在感染后到达炎性反应部位并分化为炎性DC。也有研究[8]显示炎性Mo可分化为炎性Mφ,定居Mo的游走可能依赖于受损组织或内皮细胞发出的化学信号。
更重要的是,有研究[6]进一步的证明,人类骨髓中的Mo可通过Ly6C/Gr1、CCR2和CX3CR1的表达被分为Ly6ChiMo和Ly6ClowMo。Ly6ChiMo表达高水平的Ly6C/GR1、CCR2和CD62L,但CX3CR1水平低。CCR2是趋化因子受体,可诱导Ly6ChiMo进入血液循环。Ly6ClowMo表达低水平的Ly6C/GR1、CCR2和CD62L,但表达高水平的CX3CR1[9]。Ly6ClowMo被认为是定居Mφ的前体,进入血流的Mo依赖于CX3CR1表达,但也有实验[6]结果与此相反,认为肠道Mφ在炎性反应和健康状态下均依赖于循环血液Ly6ChiMo的持续快速补充。在正常结肠中,Mo通过失去Ly6C的表达,逐渐分化为组织定居Mφ,上调CX3CR1、F4/80、CD64、CD11c等Mφ标记的表达并分泌抗炎细胞因子白介素-10(interleukin-10,IL-10)[10]。缺乏CCR2或其趋化因子配体2[chemokine (C-C motif) ligand 2,CCL2]的小鼠肠道Mφ数量减少,这意味着Mφ前体在肠道固有层的招募可能依赖于CCR2-CCL2轴。在肠道炎性反应中,组织中的定居Mφ仍然来自血液循环的Mo,只是从抗炎Mφ转变为Toll样受体(Toll-like receptor,TLR)高表达的炎性Mφ。然而,研究[9]显示血液Ly6ChiMo也可转化为Ly6ClowMo并返回至骨髓来补充定居Mφ。Bain等[11]提出不仅血液Mo可分化为Mφ,且定居Mφ具有自我更新的特点,但其机制尚未完全阐明,有研究[12]表明定居Mφ来自于一个在原位自我更新的胚胎前体,但是可以被进入肠道的Ly6ChiMo“充盈”,这些Mo作为完全应答的细胞进入肠道,随后又受到肠道微环境的调控。这个解释可能有助于解释部分定居Mφ的起源。此外,从骨髓直接进入血液的Mo具有炎性反应表型,这些细胞迅速离开血流并分化成组织巨噬细胞(tissue macrophages,TMφ)或DC。Auffray等[13]推测,最初未募集到组织的Mo可能在血液中进一步成熟并最终成为血细胞的一部分,其被认为有助于维持血管内皮细胞的完整性,并保持组织中定居Mφ稳定。因此,这些Mo是炎性反应和创伤期间组织中Mφ的主要来源,它们从血流中的迁移增强。
肠道定居Mφ产生抗炎细胞因子,特别是IL-10和转化生长因子-β(transforming growth factor-β,TGF-β),而炎性Mφ在炎性反应部位起作用并具有强杀菌特性。在肠道中,IL-10可在Mφ、Th2细胞、B细胞、自然杀伤样T细胞(natural killer T-like cells,NKT)、调节性T细胞(regulatory cells,Tregs )中表达。
活化的Mo和Mφ是IL-10的主要来源,IL-10可抑制在脂多糖(lipopolysaccharides,LPS)[14]的刺激下Mφ产生的多种促炎因子,如一氧化氮(nitric oxide,NO)等,以免造成更严重的组织损伤,其机制是通过 LPS-IκBα-NF-κB信号轴发挥作用的[15]。实验[16]表明Mφ产生的IL-10对Foxp3+Tregs表达具有重要的调节作用,CD4+Foxp3+T细胞通过IL-10和TGF-β依赖性机制维持肠内稳态。最近,关于具有Mφ类型特异性缺失IL-10的柠檬酸杆菌感染小鼠的研究[17]证明,IL-10通过调节IL-23的产生以限制固有免疫防止急性细菌感染中的过度炎性反应,而IL-10的缺乏可导致肠道Mφ染色质稳定性改变[18],这些结果提示IL-10在抑制炎性反应中起关键作用。
同时肠道定居Mφ高表达TGF-β受体并参与活化的TGF-β信号的转导,在TGF-β作用下通过TLRs激活并可转移NF-κB[8]。而TGF-β可与Tregs表达的Foxp3结合,形成CD4+Foxp3+Tregs降低黏膜Mφ激活和转位NF-κB的能力。此外,Smad7可表达阻断TGF-β信号传导并激活炎性Mφ,这一发现在坏死性小肠结肠炎Mφ 实验中得到证实[12]。
粒细胞巨噬细胞集落刺激因子(granulocyte macrophage colony stimulating factor,GM-CSF)是促进造血干细胞分化成原始Mo、幼稚Mo和Mo的主要因子。血流中循环中有不同类型Mo,部分血中Mo经历成熟,适应其局部微环境变为各种定居Mφ,部分Mo则分化成游离Mφ,在不同组织中如阿米巴虫一样迁移(图1)。
2.2 巨噬细胞的生物学特征和功能
Mφ的体积是Mo的5~10倍,且有更多的细胞器(特别是溶酶体)、褶皱和伪足。而定居Mφ分布广泛,表型众多,例如胃肠道的Mφ、肺中的尘细胞、皮肤中的朗格汉斯细胞等。Mφ是以其吞噬作用命名的,其通过调理素(Fc受体和C3b受体)或非调理素受体[甘露糖受体(mannose receptor,MMR)、清道夫受体(scavenger receptor,SR)、模式识别受体(pattern recognition receptor,PRR)、特别是TLRs],使Mφ可以参与固有免疫和获得性免疫。Mφ具有强大的吞噬功能,每天可清除约2×1011个红细胞,这相当于每年接近3 kg的铁和血红蛋白被“回收”以供机体重复使用,此过程一旦受阻机体将无法生存。同时Mφ还参与清除组织重塑过程中产生的细胞碎片,并快速有效地清除已经凋亡的细胞。此过程不依赖于免疫细胞的信号传导而发生,使未被刺激的Mφ产生很少或不产生免疫介质,即介导吞噬作用的受体不能转导诱导细胞因子基因转录的信号,或产生抑制信号和细胞因子,使Mφ每天发生的大部分吞噬作用与其他免疫细胞无关。因此,Mφ的主要作用不是免疫效应细胞的作用,而是作为普通的“清洁细胞”,清除细胞外基质的间隙环境[19]。
Mφ可通过TLRs、PRR和IL-1R识别坏死细胞碎片中存在的内源性危险信号,其中大部分信号通过髓样分化因子(myeloid differentiation factor88,MyD88)呈递。该功能使Mφ成为宿主危险的主要传感器之一。内源性危险信号可引起组织中Mφ活化,使其在固有免疫和适应性免疫中分化成不同表型。除此之外,Mφ也可被IL-8激活并释放IL-1、IL-6、IL-12和肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α),其将NE募集至炎性反应部位并产生杀菌物质,导致组织液化并形成脓液以消除入侵的病原体。而NE通过分泌存储在颗粒中乳铁蛋白、脂笼蛋白、溶菌酶、IL-37、CX3C-趋化因子配体1(chemokine CX3C motif ligand 1,CX3CL1)活化Mφ[20],使Mφ可以识别进入上皮或黏膜屏障的微生物,并伸出伪足以吞噬这些微生物,随后通过溶酶体进行消化。自然杀伤细胞(natural killer cell,NK)通过释放γ干扰素(interferon-γ,IFN-γ)或通过细胞毒性来影响Mφ的活化,同时Mφ产生的IL-1β、IL-10和TGF-β可抑制固有免疫应答,而吞噬衰老的NE,脂氧素、保护素和分解素有助于保护固有免疫应答。
3 肠内Mφ特性与功能
3.1 肠道Mφ的一般特征
Spoettl 等[21]通过体外三维共培养模型证实肠道Mφ的分化依赖于肠上皮细胞。Mφ广泛分布于所有哺乳动物的肠道中,包括黏膜层和黏膜下层。研究[22]显示肠道Mφ数量变化与肠道细菌数量相关。使用成熟小鼠Mφ标志物F4/80发现,小肠中F4/80总量为1.3×107,大肠中为1.4×107,在无菌小鼠的肠道中,Mφ的数量减少,F4/80明显降低[23],这提示Mφ具有肠道病原防御功能。
肠道中Mφ与其他组织存在差异。在生理状态下[24],Mφ不表达共刺激分子,如CD40、CD80和CD86,或诱导呼吸爆发来消灭微生物,且肠道定居Mφ对TLRs配体的反应与其他组织中有所不同[8]。TLRs是位于细胞表面或内部的膜糖蛋白,具有结合配体的胞外区和触发胞内信号传导级联的外质域。可以形成异源或同源二聚体,或与其他受体结合以识别各种微生物。Mφ可通过MyD88、TLRs结构域衔接蛋白(TIR-domain-containing adaptor inducing interferon-β,TRIF)和NF-κB介导的多种途径被激活[25],而TLRs是最具特色的PRR。然而,肠道定居Mφ对TLRs配体无反应且不产生促炎细胞因子或趋化因子,如IL-1、IL-6、IL-12、IL-23、TNF-α、趋化因子CXC配体(chemokine C-X-C motif ligand,CXCL)10(IP-10)[8,24]。推测可能是肠道的Mφ通过各种机制阻断了TLRs的下游信号,包括缺乏关键的适配器信号分子,如MyD88、白细胞介素-1受体相关激酶(interleukin-1 receptor-associated kinase,IRAK) 和TNF受体相关因子6(TNF receptor associated factor 6,TRAF6),或存在抑制分子,如IRAK-M。总的来说,这些结果导致NF-κB不能转移至核内,或不能激活基因转录[26]。当然,这并不意味着TLRs在肠道Mφ中表达或不表达不重要。事实上,Mφ中的TLRs信号通路对于保护肠道上皮免受机械和毒性损伤必不可少[27]。
3.2 肠Mφ可塑性特征
Mφ可根据环境因素改变其生理功能,而表现出一定的可塑性。Mφ可以对受伤或感染后迅速产生的内源性刺激作出反应,这些早期刺激通常由固有免疫细胞产生,并对Mφ的功能产生快速调节。同时Mφ可对由抗原特异性免疫细胞产生的信号作出反应。这些信号比固有免疫刺激局限但持续时间长,通常会引起Mφ结构和功能的改变。目前,Mφ可根据宿主防御、伤口愈合和免疫调节的自我平衡不同可分化为M1Mφ、M2Mφ、TAM和Mregs[19](图2)。
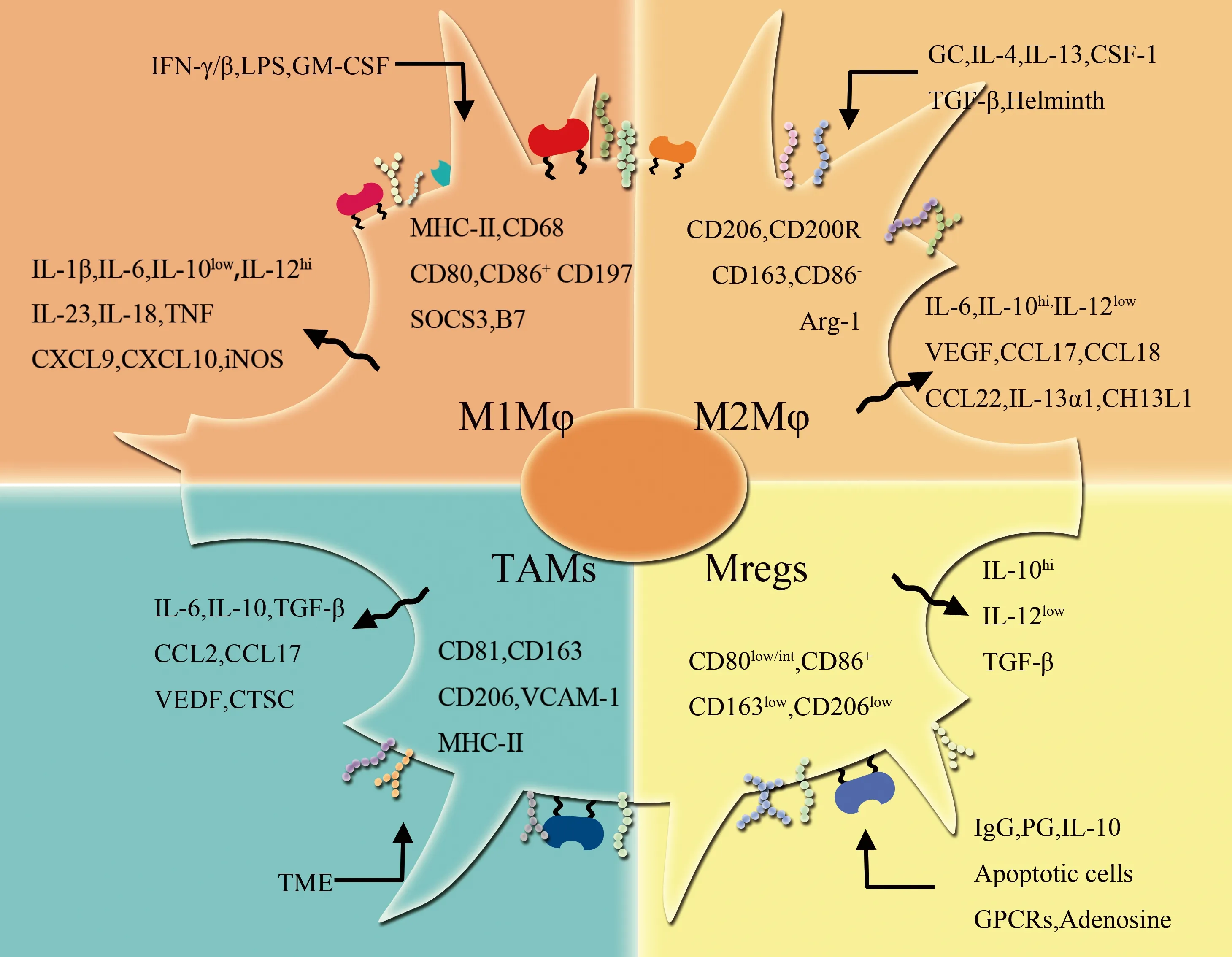
图2 Mφ的分型和各自的刺激因素、表面标记和细胞因子关联性Fig.2 Classification of Mφ and its correlation with stimulating factors, surface markers and cytokines
Black arrows represent stimulation; blue arrows represent release; green arrows represent expression;IFN-γ/β:interferon-γ/β;LPS:lipopolysaccharide;GM-CSF:granulocytemacrophage colony stimulating factor;MHC-II:major histocompatility complex class II;SOCS3: suppressor of cytokine signaling 3;TNF:tumor necrosis factor;iNOS:inducible nitric oxide synthase;GC:glucocorticoid;IL:interleukin;CSF-1:macrophage colony-stimulating factor 1;TGF-β:transforming growth factor-β;Helminth;Arg-1:arginine-1;VEGF:vascular endothelial growth factor;CCL:the chemokine (C-C motif) Ligand;CH13L1:chitinase-3-like protein-1;TME:tumor microenvironment;VCAM-1:vascular cell adhesion molecule-1;CTSC:cathepsin C;apoptotic cells;GPCRS:g-protein-coupled receptors.
3.2.1 经典活化的Mφ(classically activated macrophages,M1Mφ)
M1Mφ特指在细胞介导的免疫应答过程中产生效应的Mφ,可产生促炎细胞因子,具有很强的杀死微生物特性,但是这些特性也容易引起组织破坏。IFN-γ、LPS、GM-CSF等可激活M1Mφ[28],在M1Mφ激活过程中触发的基因是由转录因子组合诱导的,包括在IFN-γ受体后激活的信号转导和转录激活(signal transducer and activator of transcription,STAT)分子,以及作用于TLRs或TNF受体活化的NF-κB和丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)。其特征在于分泌IL-1β、TNF、IL-12和IL-18和趋化因子CXCL9、CXCL10的能力增强,可高表达主要组织相容性复合体Ⅱ类分子(major histocompatility complex class Ⅱ,MHC-Ⅱ)、CD68、CD80、CD86共刺激分子。M1Mφ细胞内的细胞因子信号传导抑制因子3(suppressor of cytokine signaling 3,SOCS3)的表达增加,激活诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)产生NO,加剧了炎性反应[29]。 由此可见,M1Mφ具有吞噬大量病原体的能力,并杀死吞噬的细菌。
在M1Mφ的激活中,IFN-γ和TNF两种信号介导下可使Mφ具有强杀伤能力并分泌促炎细胞因子和炎性介质。NK细胞通过产生IFN-γ,这可使Mφ分泌促炎细胞因子,产生更多的·O2-和NO-以增加其杀伤能力。因此,免疫介质使M1Mφ发挥抗感染能力,而IFN-γ作用于M1Mφ的效应短暂[30]。所以,通常需要适应性免疫应答来维持M1Mφ稳定的宿主防御。TNF以MyD88依赖性方式发挥作用,TLRs配体诱导TNF mRNA的转录,TNF-α与IFN-γ共同作用活化Mφ群。除了MyD88,TLRs配体还可以激活TRIF依赖性途径,通过IFN调节因子3(interferon regulatory factor 3,IRF3)调节IFN-β的产生[31]。这种内源性产生的IFN-β可以代替由NK细胞和T细胞产生的IFN-γ并激活M1Mφ。由M1Mφ产生的促炎细胞因子不仅参与宿主防御,也会造成宿主损伤。M1Mφ产生的IL-1、IL-6和IL-23与TH17细胞的发育和增生有关[32],而TH17细胞分泌的IL-17与多形核白细胞(polymorphonuclear leukocyte,PMN)募集有关,这可导致炎性反应性自身免疫病变[33]。同时M1Mφ不仅通过分泌TGF-β清除凋亡的PMNs抑制炎性反应的发生,还可激活T细胞产生的IFN-γ参与 Th1 型免疫应答,由此可见,M1Mφ发挥细胞介导的免疫应答。这提示M1Mφ是宿主防御的重要组成部分,但M1Mφ活化是一把双刃剑。M1Mφ是几种自身免疫性疾病的关键介质,包括类风湿关节炎、IBD[9]、多发性硬化症、系统性红斑狼疮、肥胖、1型糖尿病[34]。这一特性引起了科学家的高度重视。
3.2.2 选择性活化Mφ(alternative activated macrophages,M2Mφ)
又名替代性活化Mφ,在促进伤口愈合,寄生虫感染、组织重塑、血管生成和过敏性疾病的反应中发挥重要作用。巨噬细胞集落刺激因子-1(macrophage colony-stimulating factor-1,CSF-1)、IL-4、IL-13、TGF-β、糖皮质激素(glucocorticoid,GC)、真菌和蠕虫感染等可使M2Mφ激活,M2Mφ使MMR上调,诱导产生精氨酸酶-1(arginine-1,Arg-1),抑制合成NO,降低杀伤细胞内病原体的能力[35]。M2Mφ可分泌CCL17、CCL18、CCL22[36]。而IL-10 作用于Mφ可下调MHCⅡ分子、MMR和SR,降低Mφ对抗原的吞噬能力[37]。且特征性表达MMR(CD206)。然而,在2013年Jaguin等[38]观察到M1Mφ和M2Mφ之间CD206表达没有差异,并提出M2Mφ的特异性是CD200R膜糖蛋白的上调。CD163被认为是M2Mφ标志物,但最近在人体组织中检测发现,CD163必须与转录因子CMAF共同标记阳性才能作为M2Mφ标志物,单纯一个CD163不能认为是M2Mφ标志物[39]。
与M1Mφ类似,M2Mφ也可对固有免疫或适应性免疫发生反应。在组织损伤时释放IL-4,其可将定居Mφ转变为M2Mφ促进伤口愈合。同时嗜碱性粒细胞和MC是IL-4产生的早期重要来源,其他粒细胞也能产生[40]。IL-4刺激Mφ中的Arg-1活性,使其能将精氨酸转化为鸟氨酸,促进细胞外基质的产生,在IL-4存在下产生的M2Mφ上调了MMR的表达,并且产生·O2-和NO-的水平比M1Mφ低。可见M2Mφ主要与伤口愈合有关,M1Mφ高表达IL-12和低表达IL-10,而M2Mφ则变化正好相反,M2Mφ可表达IL-13α1,而M1Mφ则不表达[41],在此过程中,两者相互协调。
3.2.3 调节性Mφ(regulatory macrophages,Mregs)
类似于上述的两个Mφ群,Mregs也可在固有免疫或适应性免疫应答后产生,其主要作用是与细胞增生和免疫调节有关[42]。Mregs具有明显抑制T细胞功能。Mregs通过IFN-γ诱导吲哚胺2,3-加双氧酶活动来抑制有丝分裂原刺激T细胞增生,同时通过iNOS依赖性机制抑制多克隆T细胞的增生,并可使活化的T细胞失活。用IgG免疫复合物(immune complex,IC)和TLRs激动剂共同刺激后会导致免疫抑制性细胞因子IL-10+Mφ群发育[43-44]。除了IC之外,其他因子包括前列腺素(prostaglandin,PG)、凋亡细胞(apoptotic cell)、腺苷(adenosine)、IL-10和一些G蛋白偶联受体(G-protein-coupled receptors,GPCRs)配体也可以为Mregs的产生提供信号[45]。Mregs表型为CD80low/intermediate、CD86+、CD163low和CD206low[42],M1Mφ、Mregs的CD86均呈高度阳性,而人Mregs与小鼠Mregs表型并不完全相同。Mregs也可能在适应性免疫反应的后期阶段产生,其主要作用是抑制免疫反应并限制炎性反应[46]。人类骨髓间充质干细胞(bone marrow-derived mesenchymal stromal cells,MSCs)和间充质干细胞来源的囊泡(MSC-derived extracellular vesicles,MSC-EVs)影响人Mregs的功能[42]。MSC和MSC-EVs可下调IL-23和IL-22的生成,同时上调免疫抑制因子PGE2的生成,增强了Mregs的抗炎表型,使用PGE2与MSCs共培养,MSC-EVs可通过PGE2介导Mregs表型变化。Lo等[47]报道,MSCs能够在C57体内、外触发M1Mφ到M2Mφ表型的变化。有研究[48]显示MSCs通过增加IL-10的产生、SR的表达和吞噬能力等途径,使Mo和Mφ向M2Mφ表型分化。
虽然不同刺激产生的Mregs亚群之间可能存在细微的差异,但有些特征都是共同的。大多数Mregs所共有的一个特征是需要两种刺激来诱导其抗炎活性。单一刺激(例如,IC、PG等)本身基本无激活功能。然而,当与第二个刺激物如TLRs结合时,这两种信号共同作用可使Mregs产生IL-10和TGF-β,且Mregs产生的IL-10水平明显高于M1Mφ和M2Mφ[49],IL-10的产生是Mregs重要特征之一。除了IL-10的产生,Mregs可下调IL-12的产生[44],由于IL-10可以抑制各种促炎细胞因子的产生和活性,所以Mregs可有效抑制炎性反应。与M2Mφ不同,Mregs不参与细胞外基质的产生,但表达共刺激分子CD80和CD86,可以将抗原呈递给T细胞,是有效的抗原呈递细胞[48]。因此,Mregs和M2Mφ之间存在明确的功能以及生化差异。由于Mregs的不同亚群之间存在细微的差异,这对Mregs亚群开发可靠的生物标志物是非常重要,可用个别亚群特异性治疗药物进行靶向治疗。
由此可见,Mφ的结构和功能非常复杂。一方面,它们是体内病原体防御的先驱,并且是控制免疫反应的监管机构之一;另一方面,它们可以被认为是固有免疫和适应性免疫之间的桥梁。
4 Mφ和IBD
根据肠道Mφ维持体内平衡的机制,任何抗炎系统缺陷都可能导致免疫耐受的降低进而导致IBD。1998年,发现在小鼠结肠炎中Mφ MHC-Ⅱ分子的低表达[50]。M1Mφ产生促炎因子如IL-1、IL-12、IL-23和TNF-α,而M2Mφ释放抗炎细胞因子,包括IL-10和血管内皮生成因子(vascular endothelial growth factor,VEGF)。因此,当发生炎性反应时,Mφ可能存在适应性免疫反应的障碍。GM-CSF通过产生Mregs对CD和急性结肠炎具有保护作用[51]。经典CD14+CD16Mo在CD患者的炎性反应黏膜中分化为大量炎性Mφ[9]。IBD患者黏膜中的CD14+Mo促进TNF-α、IL-1β和IL-6的产生[52]。此外,IL-10-/-的小鼠发生自发性IBD[8]。CD患者通过TGF-β1-Smad-Snail 通路诱导肠上皮间质转化,从而导致肠道狭窄[53]。临床研究[54]显示,通过肠内营养补充TGF-β可治疗儿童的IBD,CD4+Foxp3+T细胞因无IL-10和TGF-β依赖性机制故无法保护肠道免受慢性炎性反应侵害[29]。研究[55]已证实M2Mφ 可激活Wnt信号通路,该通路与UC相关,并可促进IBD中黏膜的修复。而Hippo通路分子Yes相关蛋白(Yes-associated protein,YAP)可以通过调控Mφ极化,以及肠道菌群稳态失衡,进而加重IBD的发生[35]。这些研究[35,55]显示肠道Mφ与IBD发生与发展有很大关系。此外,核副斑点包装转录本1 (nuclear paraspeckle assembly transcript 1,NEAT1)在IBD中表达较高,通过调节肠上皮屏障和外泌体介导的Mφ极化参与炎性反应,揭示了靶向NEAT1治疗IBD的潜在策略[56]。进一步研究[57]显示,炎性反应性结肠中脂肪因子的上调可通过抑制M2Mφ极化而加重DSS诱导的IBD的发生。当然也有研究[56-57]结果存在一些疑惑。一项研究[58]显示RoRy+ILCs促使Mφ响应微生物信号并产生促炎因子IL-1β。相反,另有研究[59]证实,Mφ通过调节RoRy+ILCs激活IL-22促进负反馈调节,抑制炎性反应发生与发展。
5 Mφ和胃肠道肿瘤
Mφ与肿瘤的发生、发展密切相关,肿瘤中的Mφ的数量的减少与肿瘤的转移率显著降低有关[60]。Mφ受到不同的信号、细胞因子刺激其表型会发生变化,反映了Mφ的可塑性。M1Mφ、M2Mφ和Mregs是Mφ极化的不同状态。M1Mφ、M2Mφ两者均已在肿瘤中观察到[61]。一般认为这两种类型的Mφ在功能上是拮抗的。M1Mφ特异性表达诱导型iNOS、CD197、MHC和B7分子,抑制肿瘤生长;而M2Mφ 特异性表达CD206、CD163、Arg-1,主要参与调节炎性反应,促进血管生成、组织重塑、以及肿瘤的进展和转移。高水平M1Mφ浸润与结肠癌生存率提高有关。然而,肿瘤中通常存在大量M2Mφ浸润[62],大肠癌组织中蛋白激酶Cα(protein kinase Cα,PKCα)的高表达与M1Mφ含量增高有关,在肠道中,PKCα抑制结肠癌细胞的增生[63]。推测PKCα能通过诱导肿瘤相关巨噬细胞(tumor-associated macrophages,TAMs)在肿瘤微环境(tumor microenvironment,TME)中向M1Mφ样表型极化而发挥抗癌作用,进而抑制结肠癌细胞的增生。这表明,PKCα信号通路的调控可能是结肠癌治疗的一种新策略。
与其他Mφ类似,TAMs来源于骨髓中的造血干细胞和胚胎卵黄囊中的祖细胞[64]。多达70%的结肠癌患者在肿瘤组织中表现出较高的TAMs浸润。有关 TAMs研究[65]证实,TAMs通过释放各种细胞因子和炎性介质,如IL-6、IL-10、TGF-β、CCL2、CCL17、VEGF和组织蛋白酶C(cathepsin C,CTSC)促进免疫抑制、免疫逃避、肿瘤发生、肿瘤转移和血管生成以及入侵,TAMs产生的IL-10已经被证明可以促进肿瘤的发展[66]。TAMs可在TEM中被多种细胞因子激活,肿瘤细胞可分泌多种细胞因子影响TAMs并表达CD163、CD206、CD81、血管内皮细胞黏附分子-1(vascular cell adhesion molecule-1,VCAM-1)、MHC-Ⅱ等。TAMs在肿瘤发展和抗肿瘤中的机制复杂,临床研究表明在大多数癌症中,TAMs的丰度与肿瘤预后不良有明显的相关性,但TAMs与临床结果之间既有正相关,也有负相关。目前找到精确抗肿瘤靶点有困难,但这是未来抗肿瘤研究的重大挑战[67]。
另外,M2Mφ分泌的VEGF与肿瘤血管生成、侵袭和转移密切相关。M2Mφ通过与TNF协同作用募集到肿瘤[68]。大分子造影剂PG-Gd-NIR813显示肿瘤相关M2Mφ的双磁光成像探针,并且一些新的因素已被评估为胃肠肿瘤发展的介质,如M2Mφ分泌的CHI3L1通过激活癌细胞的IL-13受体促进胃癌和乳腺癌转移[69],Mo的MCP-1被认为是将Mφ招募到TEM中最重要的趋化因子[70]。所有这些都可能成为抗肿瘤治疗的新方法。
6 小结与展望
综上所述,Mφ可根据环境变化分化成不同的表型,充当固有免疫和适应性免疫的哨兵,为共生细菌及其宿主创造和谐环境。在肠黏膜中,起到了抑制炎性反应维持动态平衡的重要作用,这种平衡一旦打破就会降低机体免疫耐受性,引起肠道功能紊乱,从而可见它们与急性组织损伤或慢性自身免疫性疾病的密切关联。有关肠道Mφ和IBD以及肿瘤的新发现可能对研究和疾病治疗非常有帮助。
猜你喜欢
现代临床医学(2021年4期)2021-07-31 07:55:54
天津医科大学学报(2021年3期)2021-07-21 09:03:48
中华养生保健(2020年9期)2021-01-18 03:12:32
中外医疗(2016年15期)2016-12-01 04:25:44
中华老年多器官疾病杂志(2016年9期)2016-04-28 08:52:49
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:37
医学研究杂志(2015年9期)2015-07-01 17:28:19
医学研究杂志(2015年12期)2015-06-10 06:57:46
中国中医药现代远程教育(2014年15期)2014-03-01 04:27:57
现代检验医学杂志(2014年1期)2014-02-06 01:29:19
