
雌激素通过介导HOTAIR表达对子宫内膜癌细胞增生及裸鼠致瘤能力的影响
2019-12-23 08:00:16马旭兰王慧霄蒋子雯代荫梅
首都医科大学学报 2019年6期
马旭兰 夏 迪 王慧霄 蒋子雯 代荫梅
(首都医科大学附属北京妇产医院妇科,北京 100026)
子宫内膜癌是来源于子宫内膜的上皮性恶性肿瘤,近年来发病率在世界范围内明显上升,严重危害女性健康[1]。子宫内膜癌的病理类型分为雌激素依赖型(Ⅰ型,占80%)和雌激素非依赖型(Ⅱ型),无孕激素拮抗的高雌激素长期刺激与子宫内膜癌的发生密切相关[2],但雌激素在子宫内膜癌发生发展中的确切作用机制目前尚未阐明。Huang等[3]研究发现,长链非编码RNA HOTAIR在子宫内膜癌组织中存在高表达,与肿瘤分期、肌层浸润及淋巴结转移等密切相关;敲减HOTAIR表达可抑制子宫内膜癌HEC-1A细胞的增生、侵袭和迁移等恶性生物学行为进展,本课题组前期在子宫内膜癌Ishikawa细胞中也得到一致结论[4]。因此,本研究通过细胞及动物实验从基因水平探究雌激素是否通过介导HOTAIR表达对子宫内膜癌细胞增生及裸鼠致瘤能力产生影响,以期为子宫内膜癌的临床治疗提供新靶点。
1 材料与方法
1.1 材料
高分化人子宫内膜癌细胞系Ishikawa(雌激素受体阳性)为本实验室保存;RPMI-1640培养基(酚红+/-)和胎牛血清购自美国Gibco公司;碳吸附特级胎牛血清购自以色列BI公司;干扰慢病毒及引物合成购自生工生物工程(上海)股份有限公司;Prime Script反转录试剂盒和SYBR Premix Ex Taq购自美国Promega公司;雌二醇(17β-estradiol,E2)购自美国Sigma公司;E2缓释片(60 d缓释、0.72 mg/片)购自美国IRA公司;BALB/c-nu裸鼠28只(3~4周龄,雌性,SPF级)购自北京华阜康生物科技股份有限公司,实验动物许可证号:SCXK(京)2011-0012。本动物实验已通过首都医科大学动物福利协会伦理审查,伦理编号:AEEI-2018-196,于中国医学科学院医学实验动物研究所完成实验。
1.2 细胞培养与处理
Ishikawa细胞用含10%(体积分数)胎牛血清、100 U/L青霉素和100 U/L链霉素的RPMI-1640培养液,置于37 ℃、5%(体积分数)CO2细胞培养箱中培养。每1~2 d更换新鲜培养基,当细胞融合度达到80%~90%时进行传代。用E2(10 nmol/L)处理细胞4 h后提取总RNA进行后续实验。实验细胞分为4组:对照组(Ishikawa细胞)、E2组(E2+Ishikawa细胞)、E2+shNC组(E2+shNC细胞)、E2+shHOTAIR组(E2+shHOTAIR细胞)。进行雌激素实验前需更换含10%(体积分数)碳吸附胎牛血清的无酚红RPMI-1640完全培养液培养至少3 d以耗竭细胞中的雌激素。
1.3 构建慢病毒介导干扰HOTAIR表达的shHOTAIR及阴性对照shNC稳转细胞株
设计3条HOTAIR低表达的干扰序列:shHOTAIR-2379(5′-CCGGGAACGGGAGTACAGAGAGAAT CTCGAGATTCTCTCTGTACTCCCGTTCTTTTTTG-3′)、shHOTAIR-2380(5′-CCGGGCAGCACAGAGCAACTCTATACTCGAGTATAGAGTTGCTCTGTGCTGCTTTTTTG-3′)、shHOTAIR-2381(5′-CCGGGGACTGGGAGGCGCTA ATTAACTCGAGTTAATTAGCGCCTCCCAGTCCTTT TTTG-3′)及阴性对照shNC序列进行慢病毒包装(包装后表达绿色荧光蛋白及嘌呤霉素抗性),经测序证实构建成功。将构建好的干扰慢病毒分别感染Ishikawa细胞,用含嘌呤霉素的完全培养基筛选感染成功的细胞,并于荧光显微镜下筛选表达绿色荧光大于90%的细胞株,qRT-PCR检测shHOTAIR细胞株中HOTAIR的干扰效率,选择干扰效果最好的细胞株进行后续细胞及动物实验。
1.4 qRT-PCR实验
提取各组细胞总RNA,根据试剂盒说明书反转录为cDNA,各基因反应条件按试剂盒说明书操作,进行qRT-PCR。计算△Ct,以2-△△Ct表示目的基因的相对表达量。HOTAIR引物:F1(5′-GGTAGAAAAAGCAACCACGAAGC-3′)、R1(5′-ACATAAACCTCTGTCTGTGAGTGCC-3′);内参基因GAPDH引物:F2(5′-GCTGAGAACGGGAAGCTTGT-3′)、R2(5′-GCCAGGGGTGCTAAGCAG-3′)。
1.5 裸鼠成瘤实验
本实验将28只BALB/c-nu裸鼠采用数字表法随机平均分为4组:对照组(接种Ishikawa细胞)、E2组(接种E2+Ishikawa细胞)、E2+shNC组(接种E2+shNC细胞)、E2+shHOTAIR组(接种E2+shHOTAIR细胞),每组7只,组内各裸鼠进行打耳孔标记。饲养1周后,对28只裸鼠进行双侧卵巢切除术。待裸鼠状态恢复后,对需E2处理的3组裸鼠颈部皮下植入E2缓释剂。于各裸鼠右侧背部皮下对应接种处于对数生长期的4组细胞,每只裸鼠接种细胞量为5×106个/100 μL。接种细胞后每3日监测裸鼠生长情况及称质量,记录每只裸鼠的成瘤时间,计算成瘤率。细胞成瘤后开始监测移植瘤生长情况,测量移植瘤的长径a及短径b计算移植瘤体积并绘制生长曲线(移植瘤体积V=a×b2/2)。于接种第28天(移植瘤体积达到500 mm3时)终止实验,全身麻醉后颈椎脱位法处死裸鼠,完整剥离肿瘤,测量质量及长短径并拍照记录。
1.6 移植瘤组织病理学检测
将剥离的四组移植瘤各取部分于10%(体积分数)甲醛溶液固定、石蜡包埋、病理切片,进行苏木精-伊红(hematoxylin and eosin,HE)染色,镜下(200×、400×)观察移植瘤的病理形态改变。
1.7 统计学方法
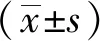
2 结果
2.1 成功构建慢病毒介导干扰HOTAIR表达的shHOTAIR及阴性对照shNC稳定转染细胞株
已成功包装HOTAIR低表达的3组慢病毒shHOTAIR-2379、shHOTAIR-2380、shHOTAIR-2381及阴性对照慢病毒shNC,经测序证实构建成功。将构建成功的慢病毒转染至Ishikawa细胞,成功构建相应4株稳转细胞,qRT-PCR结果显示(图1):shHOTAIR-2379细胞株中HOTAIR相对表达量(0.340±0.048)是shNC细胞株(0.931±0.032)的0.365倍(P<0.001);shHOTAIR-2380细胞株中HOTAIR相对表达量(0.325±0.041)是shNC细胞株的0.349倍(P<0.001);shHOTAIR-2381细胞株中HOTAIR相对表达量(0.379±0.034)是shNC细胞株的0.407倍(P<0.001)。由此可见,shHOTAIR稳转细胞株中HOTAIR表达水平均明显低于shNC细胞株,差异均有统计学意义。慢病毒介导的shHOTAIR细胞株中HOTAIR表达敲减有效,shHOTAIR-2380细胞株敲减效果最好。

图1 qRT-PCR法检测稳定转染细胞株shHOTAIR及shNC中HOTAIR的表达Fig.1 Expression of HOTAIR in stable transfected cell lines shHOTAIR and shNC was detected by qRT-PCR
2.2 qRT-PCR法检测E2对HOTAIR表达不同的4组Ishikawa细胞中HOTAIR表达的影响
qRT-PCR法检测4组细胞中HOTAIR的相对表达量发现(图2):E2组(2.084±0.052)HOTAIR的相对表达量明显高于对照组(1.000±0.046),约其2.084倍,差异有统计学意义(P<0.001);E2+shHOTAIR组(0.930±0.013)HOTAIR的相对表达量明显低于E2+shNC组(1.980±0.064),约其0.470倍,差异有统计学意义(P<0.001)。

图2 qRT-PCR法检测E2对HOTAIR表达不同的4组Ishikawa细胞中HOTAIR表达的影响Fig.2 Effect of E2 on the expression of HOTAIR in four groups of Ishikawa cells with different HOTAIR expression by qRT-PCR
2.3 成功构建E2作用于HOTAIR表达不同的4组Ishikawa细胞的裸鼠移植瘤模型
各组荷瘤裸鼠及相应移植瘤对比如下(图3A、B),本实验成功构建了雌激素E2作用于HOTAIR表达不同的4组Ishikawa细胞的裸鼠移植瘤模型。
2.4 E2对HOTAIR表达不同的4组Ishikawa细胞裸鼠致瘤能力的影响
E2组和E2+shNC组裸鼠于接种细胞第8天时全部成瘤,成瘤率均为100%;对照组裸鼠第8天时只有5只成瘤,余下2只终未成瘤,成瘤率为71.4%;E2+shHOTAIR组裸鼠于第8天时仅有2只成瘤,第11天时增加至5只,余下2只终未成瘤,成瘤率为71.4%(表1)。其中,E2组于接种细胞第15天时死亡一只荷瘤裸鼠,剩余6只。通过对以上成瘤率的比较发现,各组差异尚无统计学意义。裸鼠成瘤后通过测量并计算移植瘤体积,绘制移植瘤生长曲线(图3 C),发现在裸鼠接种细胞第26日及第28日,移植瘤体积对比:E2组>对照组,E2+shNC组>E2+shHOTAIR组,差异有统计学意义。综合比较4组裸鼠成瘤时间、成瘤率及生长曲线可得,裸鼠致瘤能力对比:E2组>对照组,E2+shNC组>E2+shHOTAIR组。

表1 4组裸鼠成瘤时间和成瘤率的比较Tab.1 Comparison of tumor formation time and tumor formation rate in four groups of nude mice
2.5 E2对HOTAIR表达不同的4组Ishikawa细胞裸鼠移植瘤终体积及终质量的影响
处死各组裸鼠后,剥离移植瘤,测量长短径及称质量,根据公式计算4组移植瘤终体积对比发现(图4A):E2组移植瘤的平均体积(488.936±130.021)mm3明显高于对照组(251.975±63.767)mm3,约1.940倍,差异有统计学意义(P<0.001);E2+shHOTAIR组移植瘤的平均体积(130.517±22.273)mm3明显低于E2+shNC组(335.271±70.768)mm3,约0.389倍,差异有统计学意义(P<0.01)。同时,4组移植瘤的终质量对比发现(图4B):E2组移植瘤的平均质量(0.377±0.035)g,明显高于对照组(0.221±0.034) g,约1.706倍,差异有统计学意义(P<0.001);E2+shHOTAIR组移植瘤的平均质量(0.130±0.031)g,明显低于E2+shNC组(0.328±0.018)g,约0.396倍,差异有统计学意义(P<0.001)。

图4 E2对HOTAIR表达不同的4组Ishikawa细胞裸鼠移植瘤终体积(A)及终质量(B)的影响Fig.4 Effects of E2 on the final volume (A) and the final weight (B) of xenograft tumors in four groups nude mice with different expression of HOTAIR
2.6 裸鼠移植瘤组织病理学检测
4组裸鼠移植瘤组织切片HE染色,镜下(200×、400×)观察移植瘤的病理形态改变发现(图5),组织内细胞量大,异形明显,核大、深染、不规则,病理核分裂活跃,并可见局部细胞出现空泡变性及坏死灶,符合子宫内膜癌细胞特征。

图5 4组裸鼠移植瘤HE染色Fig.5 HE staining of xenograft tumors in four groups nude mice
3 讨论
在人类基因组中,稳定转录的蛋白质编码基因仅占比2%,高达90%的基因转录产物为非编码RNA[5]。长链非编码RNA可在表观遗传学、转录水平及转录后水平等多层面广泛参与基因组的表达调控过程[6]。其中,HOTAIR是首个被发现的反义转录的长链非编码RNA,位于第12号染色体的HOXD基因座上,长度为2.2 kb[7]。研究显示,HOTAIR在子宫内膜癌[3]、乳腺癌[8]、宫颈癌[9]和肝细胞癌等[10]多种肿瘤及椎间盘退变[11]、软骨损伤[12]等多种非肿瘤疾病中均存在高表达水平,且与疾病的发生发展关系密切。HOTAIR的经典作用机制是发挥支架作用,招募PRC2和LSD1复合体并与之结合,定位至特定位点,导致相关抑癌基因甲基化而发生表观遗传学沉默,从而促进疾病的发生发展[13]。此外,HOTAIR还可作为竞争性内源RNA[14]、激活信号传导通路[9]、参与上皮-间质转化[15]、调控自噬发生[16]及促进蛋白质泛素化[17]等多种途径促进疾病的进展。
近年来,关于HOTAIR在多种疾病中的作用机制已取得很大进展,但HOTAIR的转录调控机制的相关研究却为数不多。Bhan等[18]发现,HOTAIR在雌激素受体阳性的人乳腺癌MCF7细胞中转录受雌激素正向调控。该研究表明,HOTAIR的启动子上含有雌激素受体反应元件序列,而雌激素可以通过与雌激素受体结合形成二聚体,进而结合至HOTAIR启动子上的雌激素反应元件,从而诱导HOTAIR转录。同样,子宫内膜癌(Ⅰ型)起源于腺上皮,发病原因与雌激素密切相关,但雌激素促进子宫内膜癌发生发展的具体作用机制目前尚不明确。因此,本研究通过细胞和动物实验两部分进行探讨雌激素是否通过介导HOTAIR表达对子宫内膜癌细胞增生及裸鼠致瘤能力产生影响。
本研究采用慢病毒携带shHOTAIR干扰基因稳定转染至Ishikawa细胞基因组的方法敲减HOTAIR表达水平[19],使HOTAIR以敲减状态在细胞中稳定传代,并通过qRT-PCR实验证实shHOTAIR稳转细胞中HOTAIR表达水平被有效敲减。本研究在体外运用雌激素E2处理HOTAIR表达不同的4组细胞,qRT-PCR法检测HOTAIR的表达水平,结果显示雌激素可上调子宫内膜癌细胞中HOTAIR的表达,而抑制HOTAIR表达可抑制该效应的发生。
本研究进一步选择3~4周龄的先天性胸腺缺陷的BALB/c-nu裸鼠,自然杀伤细胞活性低,接种癌细胞易于成瘤[20]。待裸鼠适应一周后对裸鼠进行双侧卵巢去势切除手术建立了雌激素缺乏的绝经裸鼠模型,符合子宫内膜癌多发生于绝经后女性的发病特点,也可排除雌性裸鼠内源性雌激素的干扰,更好地观察外源性雌激素对子宫内膜癌细胞裸鼠成瘤的影响。之后本研究在接种细胞前进行裸鼠颈部皮下包埋雌激素缓释剂,保证实验裸鼠每日都能定量接受雌激素刺激,相对于皮下注射给药方式[21]可以更好地模拟体内雌激素释放,药效更稳定,对裸鼠影响更小。接种细胞后8 d裸鼠出现肉眼可见的瘤结节,E2组和E2+shNC组7只全部成瘤,对照组只有5只成瘤,而E2+shHOTAIR组裸鼠在第8天时仅有2只成瘤,在第11天时,成瘤裸鼠增加至5只。对比4组裸鼠移植瘤生长曲线发现,在裸鼠接种细胞第26日及第28日,移植瘤体积E2组>对照组,E2+shNC>E2+shHOTAIR,差异有统计学意义。综合比较4组裸鼠成瘤时间、成瘤率及生长曲线可得,雌激素可以促进子宫内膜癌细胞的裸鼠致瘤能力,而抑制HOTAIR表达可抑制子宫内膜癌细胞的裸鼠致瘤能力。E2组在接种细胞第15天时死亡一只荷瘤裸鼠,分析原因可能是裸鼠免疫系统差,而雌激素可以促进移植瘤生长,可能影响该鼠正常生理机能导致死亡。通过对4组裸鼠移植瘤生长曲线、终体积及终质量对比发现,雌激素可促进子宫内膜移植瘤细胞的增生能力,抑制HOTAIR表达可抑制雌激素的促瘤增生作用。动物实验是从体内实验基础上过渡到临床应用的重要环节,本动物实验成功构建了雌激素E2作用于HOTAIR表达不同的4组Ishikawa细胞的裸鼠移植瘤模型,可为雌激素依赖性肿瘤的动物实验模型构建提供理论基础。
综上所述,本研究通过体内外实验证明雌激素可通过上调HOTAIR表达促进子宫内膜癌细胞的增生及裸鼠致瘤能力。但肿瘤的发病机制异常复杂,雌激素调控HOTAIR促进子宫内膜癌发生发展的具体机制仍需进一步深入研究。明确雌激素调控HOTAIR作用于子宫内膜癌的具体机制,可能为子宫内膜癌患者提供新的治疗靶点。
猜你喜欢
特产研究(2022年6期)2023-01-17 05:05:32
植物保护(2021年5期)2021-10-12 14:52:03
奥秘(创新大赛)(2019年9期)2019-10-09 02:03:56
小哥白尼(趣味科学)(2019年1期)2019-04-12 00:23:56
奥秘(2017年5期)2017-07-05 11:09:30
郑州大学学报(医学版)(2016年4期)2016-08-11 04:36:07
癌症进展(2016年10期)2016-03-20 13:15:42
环境与生活(2016年6期)2016-02-27 13:47:10
医学研究杂志(2015年11期)2015-06-10 06:44:03
中国当代医药(2015年16期)2015-03-01 02:03:11
