
干旱胁迫对玉米物候及产量组成的影响及模拟研究*
2019-12-11 07:42张玉书纪瑞鹏张淑杰陈妮娜
中国生态农业学报(中英文) 2019年12期
米 娜, 张玉书, 蔡 福, 纪瑞鹏**, 方 缘, 张淑杰, 陈妮娜
干旱胁迫对玉米物候及产量组成的影响及模拟研究*
米 娜1, 张玉书1, 蔡 福1, 纪瑞鹏1**, 方 缘2, 张淑杰1, 陈妮娜1
(1. 中国气象局沈阳大气环境研究所/辽宁省农业气象灾害重点实验室 沈阳 110166; 2. 中国气象局气象干部培训学院辽宁分院 沈阳 110166)
为了研究产量关键期干旱胁迫对玉米物候及产量和产量组成的影响, 评估作物生长模型对干旱胁迫下玉米物候和产量模拟的效果, 基于锦州农业气象试验站2011—2015年分期播种试验玉米产量和产量组成观测资料, 尤其是2014年和2015年天然干旱胁迫试验数据和2015年玉米开花、吐丝物候加密观测资料, 分析了产量关键期干旱胁迫对玉米物候及产量和产量组成的影响, 评估了CERES-Maize模型对不同降水年型玉米产量和产量组成的模拟效果, 提出了模型改进的方向。结果表明, 2014年和2015年辽宁省西部地区在玉米开花期前后经历了较严重的干旱胁迫过程, 干旱胁迫导致玉米吐丝延迟程度大于开花, 90%以上的植株能到达开花阶段, 但仅有45%~88%的植株能到达吐丝阶段, 直接影响株籽粒数(不同播期下的玉米株籽粒数相差32%)及最终产量(产量下降33%~78%)。CERES-Maize模型对正常年玉米产量及产量组成的模拟效果较好, 对干旱年的模拟效果较差, 部分原因在于模型在模拟玉米物候时不对开花和吐丝期加以区别, 只考虑了温度对花期的影响, 而没有考虑干旱胁迫下玉米因开花–吐丝间隔增大、雌穗发育异常、物候期推迟而造成的减产作用。因此, 玉米产量关键期干旱胁迫直接影响玉米物候(开花—吐丝期), 进而影响玉米穗粒数及最终产量; 为提高干旱胁迫下作物模型的模拟评估能力, 亟待开展干旱胁迫下基于冠层吐丝动态的玉米产量模拟研究。
玉米; 产量关键期; 干旱胁迫; 开花—吐丝间隔; 产量; 产量组成; CERES-Maize模型
我国北方大部分农区位于半干旱半湿润气候区内, 频繁出现的干旱是制约农业生产的主要气象灾害之一, 其中跨季节的长时间连续干旱对农业影响最大。对东北地区而言, 吉林省西部和辽宁省西部在玉米()生长季内为干旱高风险区[1]。研究表明, 21世纪中期东北春玉米缺水率将呈增加的趋势, 意味着干旱对当地玉米生产影响将进一步加剧[2-3]。如何及时有效地识别干旱并评估其影响, 进而提前采取抗旱减灾措施来降低可能造成的损失受到越来越多的关注[4], 干旱影响评估研究同时可为保险业参与干旱灾害事件管理奠定一定的研究基础[5]。
玉米是雌雄同株作物, 一般来说, 在适宜环境条件下, 多数玉米品种抽雄后2~4 d雄穗开花散粉, 开花散粉后2~4 d雌穗吐丝, 授粉后10~15 d籽粒体积迅速增大, 进入籽粒形成期[6]。玉米开花—吐丝间隔(anthesis-silking interval, ASI)是指玉米雄穗开花散粉和雌穗吐丝之间的时间间隔。大量研究表明, 玉米最终产量与ASI呈负相关关系[7-9]。玉米开花期前后发生干旱胁迫, 会引起玉米植株生殖生长出现问题, 延缓雌雄穗发育进程、雄穗抽出困难、雌穗吐丝延迟, 由于雄穗的延迟程度要小于雌穗吐丝的延迟, 导致ASI变大, 影响玉米授粉、受精过程, 最终导致秃尖形成, 穗粒数降低; 干旱严重时会导致雌穗发育异常(不能吐丝), 不能正常受精结实, 形成空秆。玉米苗期(三叶—拔节期)遭受干旱胁迫, 会导致生育期推迟, 株高和叶面积降低, 但对果穗性状和最终产量影响较小。吐丝—乳熟期遭受干旱胁迫, 会缩短玉米灌浆时间, 全生育期长度缩短, 籽粒产量下降[10-13]。
玉米的籽粒产量与收获时的单株籽粒数(KNP)关系密切, 相比单个籽粒重而言, 玉米单株籽粒数是最终产量的决定因素[14-17]。玉米单株籽粒数与开花期前后4~5周同化物的生产有关, 因此玉米开花期前后(通常指开花前10 d至开花后15 d)这段时间为“产量关键期”[18]。产量关键期遭受干旱胁迫会造成植株生长速率和干物质向果穗分配比例下降, 导致雌穗吐丝时间延迟、ASI变大[18,9], 对株籽粒数影响较大。调亏灌溉试验结果表明, 当在抽雄和籽粒形成阶段不进行灌溉补水时, 籽粒数下降20%~ 35%[19]。干旱胁迫控制试验结果表明, 拔节至吐丝期遭受干旱胁迫后穗粒数下降19%[13], 拔节至开花期长时间干旱或开花期前后干旱均会导致单株籽粒数显著下降(分别下降12%~20%和34%~ 41%)[20,16]。籽粒重(KW)是除了籽粒数之外的另一个产量组成要素, 基因型、作物管理措施、环境等因素是造成籽粒重发生变化的主要原因[21]。针对干旱胁迫对玉米籽粒重的影响已开展了较为广泛的研究, 不同生育阶段的干旱胁迫对玉米籽粒重的影响是较为复杂的, 这种复杂性表现在籽粒重在很大程度上受到有效灌浆期源汇比(SSR)的影响, SSR是有效灌浆期(指开花后15 d至生理成熟)生物量累积(PBG)和株籽粒数(KNP)的比值。因为PBG和KNP均会受到干旱胁迫的影响, 干旱胁迫对他们的各自影响使得KW对干旱胁迫的响应变得较为复杂[22]。
自20世纪末 CERES-Maize模型在我国开始应用以来, 许多学者对该模型进行了校准和验证, 大多研究剔除了受重大灾害影响的试验数据[23]。在干旱胁迫条件下, CERES-Maize模型用两个胁迫因子模拟作物受到的影响, 总的原则是, 比较潜在蒸散(或蒸散需水)和潜在根系吸水(或植物所吸收的土壤水)的大小, 以此判断作物生长是否受到影响并且定量化影响的大小。CERES-Maize模型中以热量时间表示物候发展进程[24], 由生长度日(growing degree-days, GDD)表示, 模型未考虑其他因素(水分、N)对物候进程的影响。在CERES-Maize模型基础上改进发展而成的IXIM模型也不区分对开花和吐丝期的模拟, 即模型只能模拟开花期(吐丝期), 并且干旱胁迫对开花期(吐丝期)物候不产生影响[25]。Lizaso等[26]针对高温对玉米物候的影响, 改进了IXIM模型对物候的模拟, 将玉米散粉看成一个连续的动态过程, 模拟每天到达开花散粉阶段的植株数量, 并把50%的植株到达开花散粉阶段的时间定为玉米的开花日期, 同时在模型中加入两个参数, 即ASNS和ASEN, 分别代表非胁迫条件下该品种的ASI和ASI对环境胁迫的响应参数, 使模型能够分别模拟开花和吐丝日期。然而在干旱胁迫条件下, 玉米的吐丝日期会延后, 且对整个冠层而言, 玉米吐丝的持续时间(从第1株玉米吐丝至最后1株玉米吐丝完成)也会延长, 有部分植株可能受干旱影响不能吐丝, 目前作物生长模型还难以实现对玉米吐丝动态及吐丝百分率的模拟。CERES-Maize模型的最初版本[27]中使用1个非线性函数来预测KNP, 即KNP是吐丝后植株生长速率(PGR)的函数。同时也定义了1个由品种决定的单株最大籽粒数(即潜在籽粒数)。后来的CERES-Maize模型版本又使用线性函数来预测KNP, 同时定义了该线性函数的最大值(即单株最大籽粒数), 函数的参数和单株最大籽粒数由基因型决定。CSM-IXIM模型对KNP的模拟与CERES-Maize模型的不同在于使用开花前后平均植株生长速度函数(而不是吐丝后的PGR函数)来预测KNP[25]。函数形式也由单一函数变成了双函数, 即下面的函数曲线预测顶穗的籽粒数, 上面的曲线预测顶穗和下侧穗的籽粒数之和。作物生长模型中对玉米KNP的模拟是为了模拟目的而采取的一种简化方法, 没有考虑极端状况下的情况, 难以代表玉米生长的真实状况。比如, 如果玉米因受到环境胁迫在开花前后那段时间里生长速率很低, 玉米可能不能形成籽粒[28]。尽管Lizaso等[26]对CSM-IXIM模型中物候的模拟进行了改进, 考虑了干旱胁迫下的ASI, 但对吐丝物候的模拟仍是建立在经验函数的基础上, 没有建立基于玉米生长发育过程的物候模拟机制, 同时没有考虑干旱胁迫对玉米植株吐丝百分率的影响, 也没有建立ASI和吐丝百分率与单位面积上玉米籽粒数之间的关系, 因此干旱胁迫下玉米株籽粒数的模拟仍需进一步完善。最近Durand等[29]研究也指出, 作物生长模型中籽粒数的模拟模块仍然需要给予特别的关注, 需要使用更加可靠的函数和更好的输入数据来对模型进行调整。本研究在分析产量关键期干旱胁迫对玉米开花、吐丝物候以及产量和产量组成影响的基础上, 检验CERES-Maize模型对不同年型玉米产量和产量组成的模拟效果, 提出了模型改进的方向, 以期为提高干旱影响评估准确性提供借鉴。
1 研究地区概况与研究方法
1.1 试验设置及测定
玉米分期播种试验于2011—2015年在辽宁省锦州农业气象试验站(41°08′N, 121°10′E, 海拔27.4 m)开展, 该试验站为中国气象局设立的东北地区生态与农业气象野外科学试验基地之一。该研究区属温带季风型大陆性气候, 四季分明, 多年(1981—2010年)平均气温为9.9 ℃, 平均降水量为568 mm。试验区的土壤为典型棕壤, pH 6.3, 有机质含量为18.0 g·kg-1; 0~100 cm土层田间持水量22.3%, 凋萎湿度6.5%, 容重1.6 g·cm-3。2011—2015年各设置5个玉米播期(处理), 分别为4月20日(处理A)、4月30日(处理B)、5月10日(处理C)、5月20日(处理D)和5月30日(处理E)。试验玉米品种为‘丹玉39’, 该品种为稀植大穗型晚熟品种, 适宜在辽宁省南部及沿海地区种植。试验小区长15 m, 宽 10 m, 设3个重复, 分期播种试验播种行距为60 cm, 株距为40 cm, 播种时施底肥750 kg·hm-2, 底肥为复合肥料, N、P2O5、K2O含量分别为12%、18%和15%, 总养分≥45%。各个播期处理仅在播种当日灌溉1次, 以保证玉米出苗。
试验过程中记录所有播期的玉米发育期(出苗、三叶、七叶、拔节、抽雄、开花、吐丝、乳熟、成熟), 以处理内50%的植株达到某一发育期为该处理达到这一发育期的日期。对于干旱较严重的2015年, 对各处理产量关键期内的开花和吐丝期进行了加密观测, 测定每个处理达到开花和吐丝阶段的植株数量, 并计算各处理达到开花和吐丝阶段的植株百分率。成熟期, 每个处理收获20株玉米: 将玉米齐地面割下, 风干, 测定每株玉米的籽粒重; 将玉米脱粒, 充分混合, 数出8个百粒, 测定平均百粒重; 株籽粒数利用株籽粒重与百粒重相除计算得出, 最终产量利用株籽粒重与单位面积上的有效株数的乘积计算。
1.2 降水及土壤水分状况
2014年和2015年降水数据来源于锦州气象观测站, 两年4—9月的降水累积量较多年(1981—2010年)平均值偏少约40%, 降水偏少时段集中于7—9月(图1)。
玉米生育期内每10 d利用人工土钻取土法测定一次0~50 cm土壤含水量, 每次取样重复4次, 用土壤相对湿度(土壤质量含水量与田间持水量的比值)表示。2014年和2015年的4—9月土壤相对湿度变化情况及其与玉米各播期发育期的关系如图2所示, 总体来看, 2014年土壤水分状况好于2015年。
1.3 CERES-Maize模型校正及评价
米娜等[30]曾利用2011—2013年(2011年和2013年降水较多年平均值偏少20%, 2012年较多年平均值偏多48%)锦州分期播种试验数据对CERES-Maize模型进行了校正和评价, CERES-Maize模型能够较好地模拟不同播期玉米的物候期和产量, 其归一化均方根误差(NRMSE)小于10.3%。本研究拟开展模型对正常年(2012—2013年)和降水偏少年(2014—2015年)产量及产量组成的模拟效果评估, 利用误差百分比、均方根误差(RMSE)、归一化均方根误差(NRMSE)和决定系数(2)等指标来评价模型的模拟效果。其中NRMSE可表征观测值与模拟值的相对差异, NRMSE小于20%表示模拟效果好, 20%~30%表示模拟效果较好, 超过30%表示模拟效果较差[31]。模型使用的遗传参数值见表1。
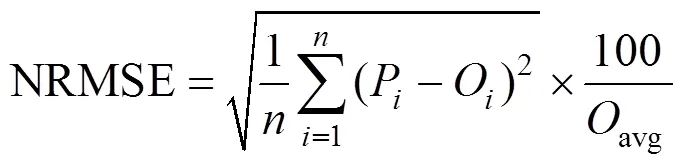

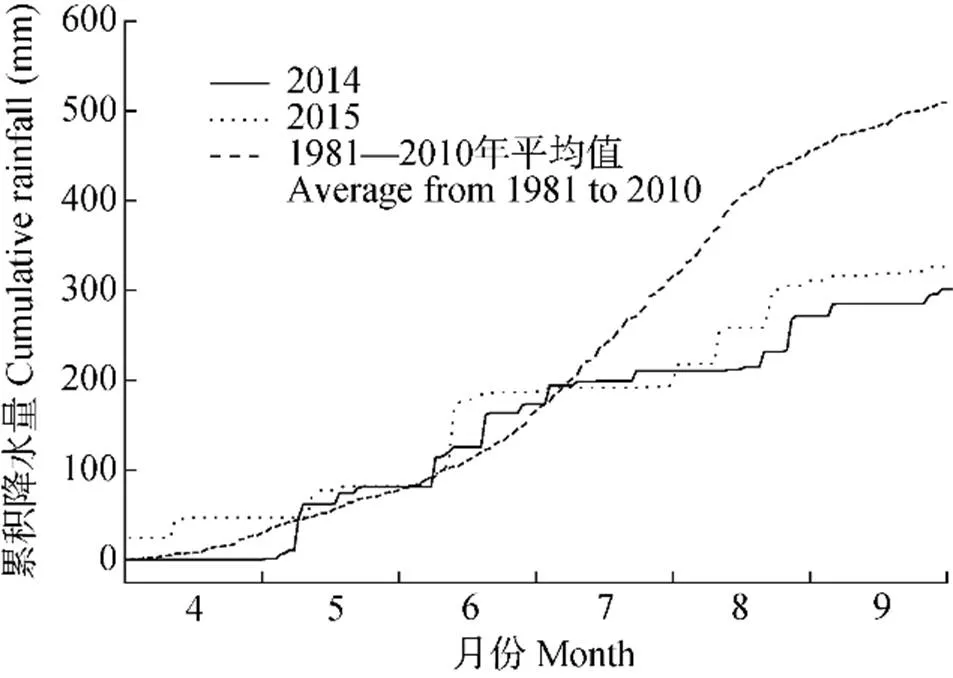
图1 锦州农业气象试验站2014年和2015年的4—9月累积降水量与多年(1981—2010年)平均值的比较
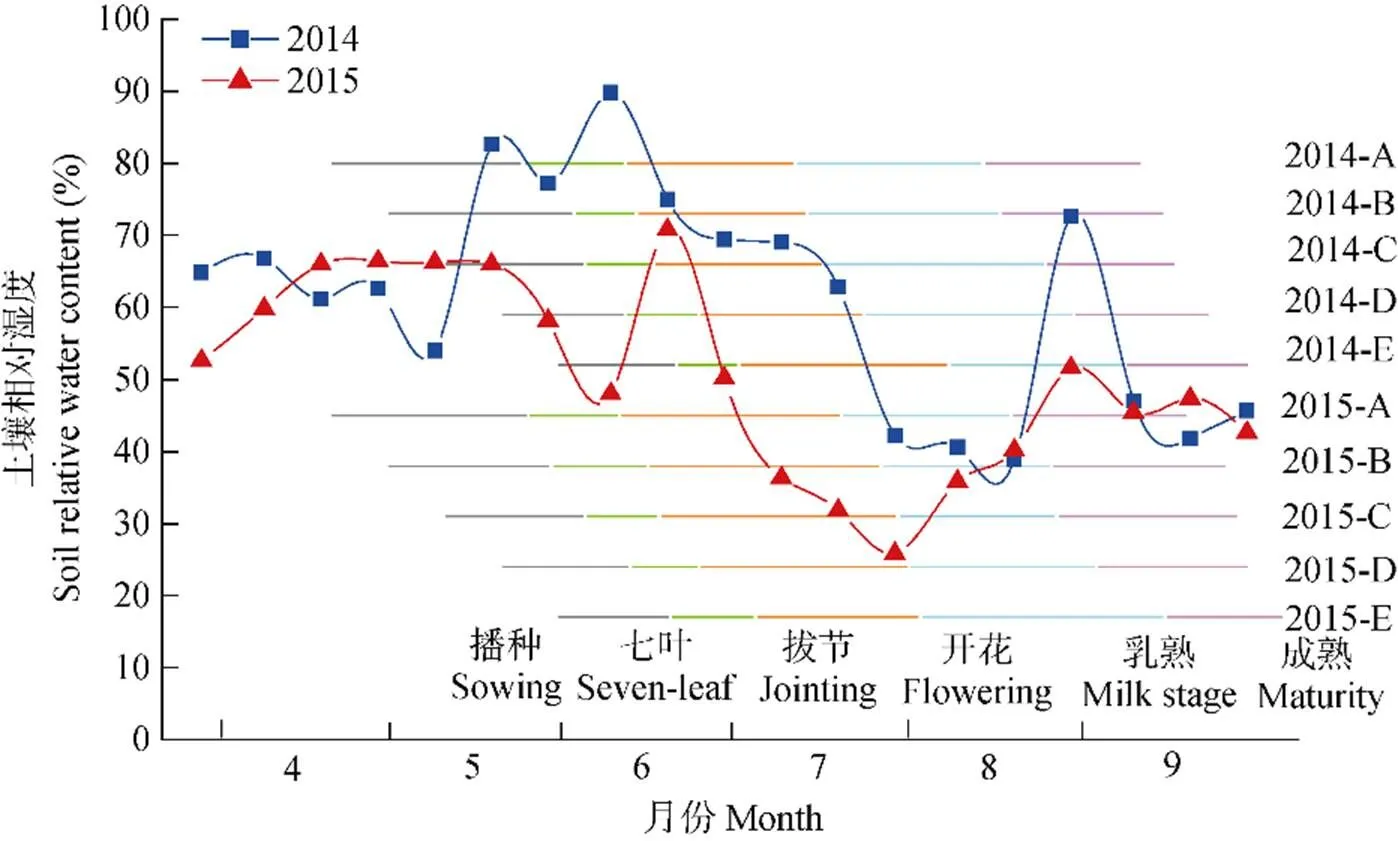
图2 锦州农业气象试验站玉米生长季土壤相对含水量(0~50 cm,每10 d测定1次)与2014年和2015年4月20日(A)、4月30日(B)、5月10日(C)、5月20日(D)和5月30日(E)播种的玉米各发育期时间

表1 CERES-Maize模型中玉米品种‘丹玉39’遗传参数取值
式中:P为第个模拟值,O为第个观测值,avg和avg分别为观测平均值和模拟平均值,为观测值或模拟值的个数。
2 结果与分析
2.1 产量关键期干旱对开花、吐丝及雌雄穗发育的影响
2015年产量关键期干旱导致各播期开花、吐丝日期不同程度延迟, 总体来看, 吐丝的延迟程度大于开花, 由此导致开花—吐丝间隔加大。受干旱影响, 2015年各播期玉米植株的开花日期比常年推迟1~14 d, 吐丝日期推迟5~22 d, 开花—吐丝间隔日数为3~15 d。从图3可见, 前3个播期(即4月20日—5月10日播种)的开花日期和吐丝日期延迟天数较多, 分别延迟10~14 d和12~22 d, 原因在于干旱对这3个播期的影响开始较早(从拔节后10 d左右开始), 且持续时间较长, 有的甚至持续到了灌浆初期。干旱对后2个播期(即5月20—30日播种)开花、吐丝期的延迟程度较前3个播期轻, 原因在于随着播期的推迟, 后2个播期生育期内气温逐渐升高, 缩短了生育期持续时间。

图3 2015年不同播期下玉米开花日期(月-日)、开花—吐丝间隔(方框内数据)及吐丝日期(月-日)
括号内数字代表与常年比较发育期的推迟天数。Numerals within brackets show the delay days compared to a normal year.
产量关键期干旱不仅延迟玉米开花、吐丝日期, 还会导致雄穗和雌穗发育异常。4月20日播种处理中有12%的植株未能抽雄开花, 其余达到抽雄开花期的植株中仅有一半植株到达了吐丝阶段。随着播期的推迟, 到达抽雄开花期和吐丝期的植株比例逐渐升高。总体来看, 产量关键期干旱对吐丝的影响重于抽雄开花, 如尽管在5月20日播种和5月30日播种处理下, 雄穗都能够正常发育, 但有12%左右的雌穗发育异常, 不能正常吐丝(图4)。
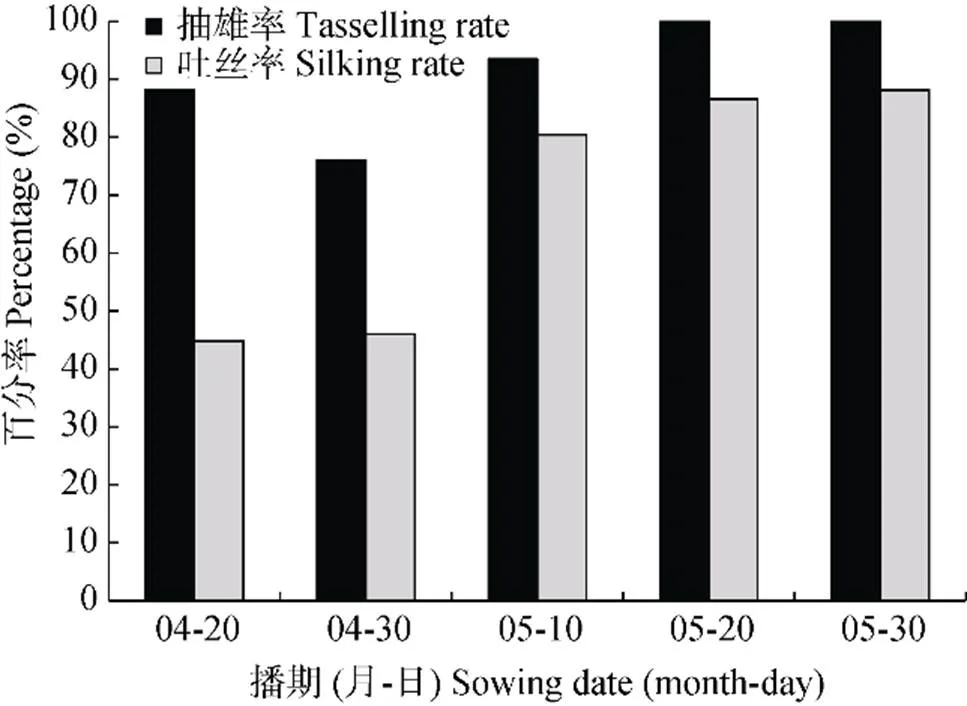
图4 2015年不同播期下玉米抽雄和吐丝植株的百分比
2.2 产量关键期干旱对玉米产量及产量组成的影响
由于2015年土壤干旱状况重于2014年, 因此2015年5个播期的籽粒产量均小于2014年各个播期。2015年随着播期的推迟, 籽粒产量逐渐增加, 较晚播期(5月20日和5月30日播种)的籽粒产量较其他播期显著增加13.6%~34.3%。与2015年情况相反, 2014年各播期籽粒产量随着播期的推迟而减少, 最高产量(4月20日播种)比最低产量(5月20日播种)高71.0%。
株籽粒重由株籽粒数和籽粒重决定, 均受到播期的影响。2014年, 随着播期的推迟株籽粒数逐渐减少, 2015年的情况则正好相反, 最后一个播期取得了最大的株籽粒数(表2)。2014年和2015年的数据均表明, 籽粒产量与株籽粒数呈显著的正相关关系(图略,=0.96,<0.001), 株籽粒数的变化能够解释93%的籽粒产量的变化。两年中各播期籽粒重的变化均呈现出先增加后减少的趋势, 较晚播种的玉米受到开花吐丝期和灌浆期干旱的影响, 或由于灌浆时长缩短, 或由于灌浆期干物质不足, 导致籽粒重减小。
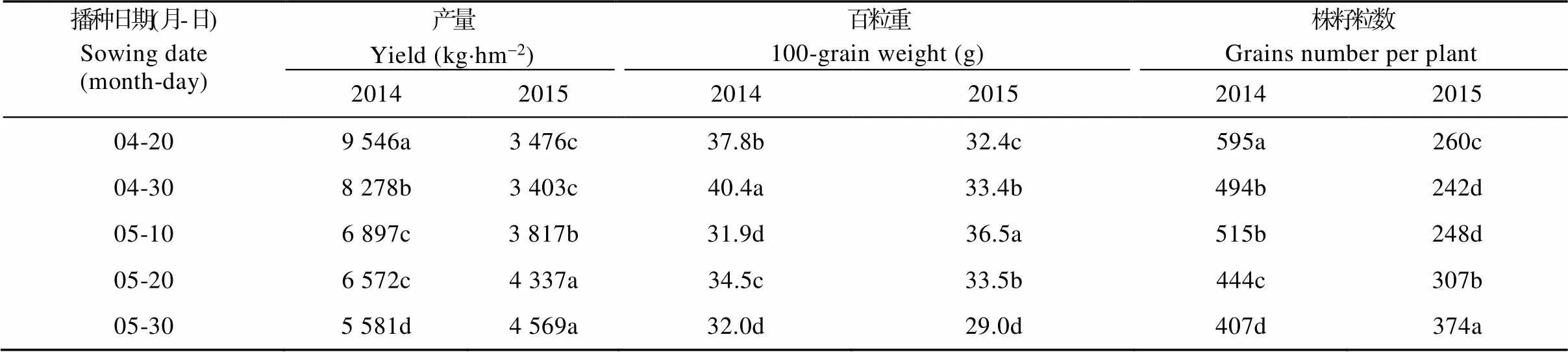
表2 播期对玉米籽粒产量、百粒重和株籽粒数的影响
同列不同小写字母表示不同播期间在0.05水平差异显著。Differing lowercase letters within a column mean significant differencesat 0.05 probability level (LSD).
2.3 作物生长模型对玉米产量及产量组成模拟效果评估
2.3.1 对正常年分期播种试验产量及产量组成的模拟
应用作物生长模型CERES-Maize对2012年和2013年玉米籽粒产量、株籽粒数、籽粒重进行模拟。结果表明, 2012年和2013年产量模拟的误差百分率分别为5.3%和9.9%, RMSE分别为593 kg×hm-2和928 kg×hm-2, NRMSE分别为6.5%和10.8%; 株籽粒数模拟的误差百分率分别为12.3%和12.9%, RMSE分别为85粒×株-1和80粒×株-1, NRMSE分别为14.6%和14.1%; 百粒重模拟的误差百分率分别为9.0%和7.0%, RMSE分别为4.9 g和3.2 g, NRMSE分别为13.1%和9.2%。整体来看, 模型对正常年分期播种试验产量及产量组成的模拟效果较好(图5)。
2.3.2 对干旱年分期播种试验产量及产量组成的模拟
模型对2014年和2015年分期播种试验产量和株籽粒数的模拟效果较差, 对百粒重的模拟效果较好(图6)。产量模拟的误差百分率分别为23.7%和97.2%, RMSE分别为1 811 kg×hm-2和3 751 kg×hm-2, NRMSE分别为24.6%和95.7%; 株籽粒数模拟的误差百分率分别为18.3%和76.0.0%, RMSE分别为106粒×株-1和219粒×株-1, NRMSE分别为21.6%和76.5%。百粒重模拟的误差百分率分别为7.9%和6.1%, RMSE分别为3.4 g和4.3 g, NRMSE分别为10.4%和13.1%。2014年干旱主要发生在玉米生育后期, 且前两个播期播种较早受干旱影响较小, 模型对其产量和株籽粒数的模拟效果尚可(A、B播期产量模拟误差百分率9.3%, 株籽粒数模拟误差百分率为8.2%), 随着播期的推迟, 干旱对玉米生长发育的影响愈发凸显, 产量和株籽粒数均有所下降, 且模型模拟效果变差。2015年干旱影响开始较早, 且持续时间长, 导致玉米产量大幅下降, 模型对产量和株籽粒数的模拟效果很差。
3 讨论与结论
玉米开花的时间由基因型、温度和光周期决定, 不受植株生长速率(plant growth rate, PGR)的影响[32]。但也有研究表明, 玉米营养生长阶段受干旱胁迫会延迟开花散粉时间, 并且降低散粉的速率[33]。与开花不同的是, 吐丝的时间依赖于PGR, 是果穗干物质累积量的函数[18]。影响PGR的环境状况, 比如干旱胁迫或者氮素缺乏, 都会延迟吐丝的时间, 引起穗粒数的下降[34-35]。本研究通过对2015年锦州农田试验站不同播期开花、吐丝动态的观测, 发现干旱胁迫导致玉米吐丝延迟程度大于开花(ASI为3~15 d不等), 90%以上的植株能到达开花阶段, 但仅有45%~88%的植株能到达吐丝阶段, 直接影响株籽粒数及最终产量(产量下降33%~78%)。Yao等[32]的研究表明, 当在玉米开花期对其进行去叶处理, 以达到影响PGR的目的时, 雄穗的开花时间没有受到影响, 雌穗的吐丝时间受到了相应的影响, 即植株的叶面积指数越小(植株获得的光能资源越少), 则吐丝的延迟越严重。在极端环境下, 能够到达吐丝阶段的植株百分率甚至不足50%, 但雄穗基本能百分之百到达开花阶段。因此表明, 在群体水平上, PGR的下降对开花和吐丝的影响存在较大差别。
本研究结果表明, 不同播期下的玉米株籽粒数相差32%左右, 产量关键期遭受干旱胁迫导致株籽粒数下降, 原因是当PGR因受到干旱胁迫影响而降低时, 果穗的生长也会下降, 不仅是由于整个植株积累的生物量降低, 还由于总生物量向果穗的分配比例下降[9]。如Andrade等[36]的试验数据表明, 当某一个玉米品种开花期间日生长量超过3 g∙株-1时, 干物质向果穗的分配比例大约为1/6, 当日生长量为2 g∙株-1时, 该比例下降为1/18, 当日生长量不足1 g∙株-1时, 该比例下降为0。由此可见, 开花期前后干旱胁迫, 造成植株生长速率和干物质向果穗分配比例下降, 导致雌穗吐丝时间延迟、ASI变大[18], 对株籽粒数影响较大。针对干旱胁迫对籽粒重的影响分析已开展了较为广泛的研究。Ouattar等[37]研究发现吐丝后期和灌浆期以及灌浆后期至生理成熟期的短期干旱不会对籽粒重产生显著的影响。然而, 吐丝后期至生理成熟期期间的长期干旱会使籽粒重下降50%。Çakir[19]研究表明各生长阶段的水分亏缺都可能会导致籽粒重降低, 但在不同的年份影响程度不同。Aydinsakir等[38]研究表明玉米全生育期灌溉量减少50%会显著降低籽粒重。Hammad等[39]发现在玉米营养生长阶段和生殖生长阶段遭受水分亏缺时, 籽粒重均显著下降。调亏灌溉研究发现在营养生长和开花阶段, 充分灌溉和调亏灌溉下的籽粒重无显著差别, 在籽粒灌浆期无灌溉会显著降低籽粒重[22,27]。本研究结果表明, 由于干旱导致灌浆时长缩短, 而且灌浆期干物质积累不足, 致使籽粒重变小。

图5 正常年(2012—2013年)不同播期玉米产量(a, b)、株籽粒数(c, d)、百粒重(e, f)的模拟值与实测值比较
作物生长模型中模拟产量的形成主要有两种类型: 一是模拟植株生物量, 通过生物量向储藏器官的分配或收获指数模拟产量(如WOFOST、INTERCOM、Aquacrop和CropSyst模型); 二是模拟产量组成, 即分别模拟籽粒数和籽粒重(如STICS、Hybrid-Maize和CERES-Maize模型)[40]。CERES-Maize模型中作物产量=株籽粒数×粒重×单位面积株数(密度)。该模型的最初版本中使用一个非线性函数[7]来预测株籽粒数KNP, 即KNP是吐丝后(籽粒形成期)植株生长速率PGR的函数。同时也定义了一个由品种决定的单株最大籽粒数(即潜在籽粒数)。后来的CERES-Maize模型版本又使用线性函数来预测KNP, 同时定义了该线性函数的最大值(即单株最大籽粒数), 函数的参数和单株最大籽粒数由基因型决定。籽粒重是通过与温度有关的最优生长速率函数及灌浆时长模拟, 籽粒以一定的生长速率进行干物质积累, 当灌浆期结束后籽粒重不再增加[24]。干旱胁迫下, 模型对产量和产量组成的模拟效果较差, 部分原因在于CERES-Maize模型在模拟玉米物候时不对开花和吐丝期加以区别, 往往只考虑了温度对花期的影响, 而没有考虑干旱胁迫下玉米因开花–吐丝间隔增大、雌穗发育异常、物候期推迟而造成的减产作用。

图6 干旱年(2014—2015年)不同播期玉米产量(a, b)、株籽粒数(c, d)、百粒重(e, f)的模拟值与实测值比较
Borrás等[18]发展了一个基于植株生长和生物量向果穗分配模拟ASI的冠层吐丝动态定量理论模型(以下简称冠层吐丝动态模型), 从而建立了果穗生物量累积和ASI之间的直接关系。冠层吐丝动态模型考虑了冠层内植株个体间PGR的差异, 模型中有两个重要的概念, 即基本植物生长速率(PGRb, 决定果穗是否生长)和果穗生物量阈值(EBt, 决定是否吐丝)。冠层吐丝动态模型能够较准确地模拟冠层吐丝的时间及吐丝植株百分率, 这表明将吐丝过程与开花期植株生长相耦合, 是模拟冠层尺度ASI的一个有效的手段。在最终产量模拟时是否应将未吐丝的植株加以考虑, 将CERES-Maize模型原来的产量计算公式: 籽粒产量=株籽粒数×粒重×单位面积株数(密度), 修正为: 籽粒产量=株籽粒数×粒重×单位面积有效株数, 仍需要开展相关的试验来进一步研究。此外, 开花前后的环境状况不仅影响植株的生长, 也影响ASI(主要由于雌穗吐丝时间发生变化)[18], ASI与籽粒数呈负相关关系, 因此, 是否可以尝试基于不同ASI植株所占的百分比和不同ASI下籽粒结实状况模拟籽粒数。总之, 对于ASI和吐丝百分率的模拟为干旱胁迫下准确模拟KNP提供了新的途径, 非常有必要进一步开展研究, 改进模型, 提高模型对干旱胁迫影响的模拟准确率。
综上, 产量关键期干旱胁迫导致玉米吐丝延迟程度大于开花, ASI为3~15 d不等。干旱胁迫影响下, 90%以上的玉米植株能到达开花阶段, 但仅有45%~88%的植株能到达吐丝阶段, 直接影响株籽粒数及最终产量(产量下降33%~78%)。CERES-Maize模型对产量关键期干旱胁迫下玉米的产量及产量组成模拟较差, 部分原因在于模型在模拟玉米物候时不对开花和吐丝期加以区别, 往往只考虑了温度对花期的影响, 而没有考虑干旱胁迫下玉米因开花–吐丝间隔增大、雌穗发育异常、物候期推迟而造成的减产作用。冠层吐丝动态模型为干旱胁迫下准确模拟KNP提供了新的途径, 非常有必要进一步开展研究, 将其嵌入作物生长模型, 以提高作物模型对干旱影响模拟的准确率。
[1] 杨晓晨, 明博, 陶洪斌, 等. 中国东北春玉米区干旱时空分布特征及其对产量的影响[J]. 中国生态农业学报, 2015, 23(6): 758–767Yang X C, Ming B, Tao H B, et al. The spatial distribution characteristics of drought and impacts on yield of spring maize in Northeast China [J]. Chinese Journal of Eco-Agriculture, 2015, 23(6): 758–767
[2] 赵宗慈, 罗勇. 21世纪中国东北地区气候变化预估[J]. 气象与环境学报, 2007, 23(3): 1–4Zhao Z C, Luo Y. Projections of climate change over northeastern China for the 21st century [J]. Journal of Meteorology and Environment, 2007, 23(3): 1–4
[3] 卢洪健, 莫兴国, 孟德娟, 等. 气候变化背景下东北地区气象干旱的时空演变特征[J]. 地理科学, 2015, 35(8): 1051–1059Lu H J, Mo X G, Meng D J, et al. Analyzing spatiotemporal patterns of meteorological drought and its responses to climate change across Northeast China[J]. Scientia Geographica Sinica, 2015, 35(8): 1051–1059
[4] Hussain H A, Men S N, Hussain S, et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids[J]. Scientific Reports, 2019, 9: 3890
[5] 朱思颖和郭高平. 保险业参与干旱灾害事件管理研究[J].经济研究导刊, 2018, (30): 86–88Zhu S Y and Guo G P. Research on the relationship between insurance industry and drought events management[J]. Economic Research Guide, 2018, (30): 86–88
[6] 张国平, 周伟军. 作物栽培学[M]. 杭州: 浙江大学出版社, 2001Zhang G P, Zhou W J. Crop Cultivation Science[M]. Hangzhou: Press of Zhejiang University, 2001
[7] Edmeades G O, Daynard T B, The development of plant-to-plant variability in maize at different planting densities[J].Canada Journal of Plant Science, 1979, 59: 561–576
[8] Hall A J, Villela F, Trapani N, et al. The effects of water stress and genotype on the dynamics of pollen-shedding and silking in maize[J]. Field Crops Research, 1982, 5: 349–363
[9] Borrás L, Vitantonio-Mazzini L N. Maize reproductive development and kernel set under limited plant growth environments[J]. Journal of Experimental Botany, 2018, 69(13): 3235–3243
[10] Bolaños J, Edmeades G O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize[J]. Field Crops Research, 1996, 48: 65–80
[11] 李叶蓓, 陶洪斌,王若男, 等. 干旱对玉米穗发育及产量的影响[J]. 中国生态农业学报, 2015, 23(4): 383–391Li Y B, Tao H B, Wang R N, et al. Effect of drought on ear development and yield of maize[J]. Chinese Journal of Eco-Agriculture, 2015, 23(4): 383–391
[12] Turc O, Bouteillé M, Fuad-Hassan A, et al. The growth of vegetative and reproductive structures (leaves and silks) respond similarly to hydraulic cues in maize[J]. New Phytologist, 2016, 212(2): 377–388
[13] 纪瑞鹏, 车宇胜, 朱永宁, 等. 干旱对东北春玉米生长发育和产量的影响[J]. 应用生态学报, 2012, 23(11): 3021–3026Ji R P, Che Y S, Zhu Y N, et al. Impacts of drought stress on the growth and development and grain yield of spring maize in Northeast China[J]. Chinese Journal of Applied Ecology, 2012, 23(11): 3021–3026
[14] Otegui M E. Prolificacy and grain yield components in modern Argentinean maize hybrids[J]. Maydica, 1995, 40: 371–376
[15] Chapman S C, Edmeades G O. Selection improves drought tolerance in tropical maize populations II. Direct and correlated responses among secondary traits[J]. Crop Science, 1999, 39: 1315–1324
[16] Mi N, Cai F, Zhang Y S, et al. Differential responses of maize yield to drought at vegetative and reproductive stages[J]. Plant Soil Environment, 2018, 64(6): 260–267
[17] Mi N, Zhang Y S, Cai F, et al. Higher yields of rain-fed maize affected by drought and sowing date in a semi-humid region of North-eastern China[J]. International Journal of Agriculture and Biology, 2019, 22(1): 83–90
[18] Borrás L, Westgate M E, Astini J P, et al. Coupling time to silking with plant growth rate in maize[J]. Field Crops Research, 2007, 102: 73–85
[19] Çakir R. Effect of water stress at different development stages on vegetative and reproductive growth of corn[J]. Field Crop Research, 2004, 89: 1–16
[20] 米娜, 蔡福, 张玉书, 等. 不同生育期持续干旱对玉米的影响及其与减产率的定量关系[J]. 应用生态学报, 2017, 28(5): 1563–1570Mi N, Cai, F, Zhang Y S, et al. Effects of continuous drought during different growth stages on maize and its quantitative relationship with yield loss[J]. Chinese Journal of Applied Ecology, 2017, 28(5): 1563–1570
[21] Borrás L, Gambín B L. Trait dissection of maize kernel weight: Towards integrating hierarchical scales using a plant growth approach[J]. Field Crops Research, 2010, 118: 1–12
[22] Wang J T, Kang S Z, Du T S, et al. Estimating the upper and lower limits of kernel weight under different water regimes in hybrid maize seed production[J]. Agricultural Water Management, 2019, 213: 128–134
[23] 熊伟, 林而达. CERES-Maize区域应用效果分析[J]. 中国农业气象, 2009, 30(1): 3–7Xiong W, Lin E D. Performance of CERES-Maize in regional application[J]. Chinese Journal of Agrometeorology, 2009, 30(1): 3–7
[24] Jones J W, Hoogenboom G, Porter C H, et al. DSSAT cropping system model[J]. European Journal of Agronomy, 2003, 18: 235–265
[25] LIZASO J I, BOOTE K J, JONES J W, et al. CSM-IXIM: a new maize simulation model for DSSAT version 4.5[J]. Agronomy Journal, 2011, 103: 766–779
[26] LIZASO J I, RUIZ-RAMOS M, RODRIGUEZ L. Modeling the response of maize phenology, kernel set, and yield components to heat stress and heat shock with CSM-IXIM[J]. Field Crops Research, 2017, 214: 239–252
[27] JONES C A, KINIRY J R. CERES-Maize: A Simulation Model of Maize Growth and Development[M]. Texas A &M Univ. Press, 1986
[28] TOLLENAAR M, DWYER L M, STEWART D W. Ear and kernel formation in maize hybrids representing three decades of grain yield improvement in Ontario[J]. Crop Science. 1992, 32: 432–438
[29] DURAND J L, DELUSCA K, BOOTE K, et al. How accurately do maize crop models simulate the interactions of atmospheric CO2concentration levels with limited water supply on water use and yield[J]. European Journal of Agronomy 2017, doi:10.1016/j.eja.2017.01.002
[30] 米娜, 张玉书, 纪瑞鹏, 等. 基于作物模型与最佳季节法的锦州地区玉米最佳播种期分析[J]. 中国农业气象, 2016, 37(1): 68–76Mi N, Zhang Y S, Ji R P, et al. Analysis on optimum sowing date of maize in Jinzhou using crop growth model and optimum season method[J]. Chinese Journal of Agrometeorology, 2016, 37(1): 68–76
[31] Jamieson P D, Porter J R, Wilson D R. A test of the computer simulation model ARC-WHEAT on wheat crops grown in New Zealand[J]. Field Crops Research, 1991, 27: 337–350
[32] Yao N R, Yeboua K, Kafrouma A. Effects of intensity and timing of defoliation and growth, yield components and grain yield in maize[J]. Experimental Agriculture, 1991, 27: 137–144
[33] Wang J T, Kang S Z, Zhang X T, et al. Flowering characteristics and yield of maize inbreds grown for hybrid seed production under deficit irrigation[J]. Crop Science, 2017, 57: 2238–2250
[34] Hall A J, Lemcoff J H, Trápani N. Water stress before and during flowering in maize and its effects on yield, its components, and their determinants[J]. Maydica, 1981, 26: 19–38
[35] Edmeades G O, Bazinger M, Hernández M, et al. Causes for silk delay in a lowland tropical maize population[J]. Crop Science, 1993, 33: 1029–1035
[36] Andrade F H, Vega C, Uhart S, et al. Kernel number determination in maize[J].Crop Science, 1999, 39: 453–459
[37] Ouattar S, Jones R J, Crookston R K. Effect of water deficit during grain filling on the pattern of maize kernel growth and development[J]. Crop Science, 1987, 27: 726–730
[38] Aydinsakir K, Erdal S, Buyuktas D, et al. The influence of regular deficit irrigation applications on water use, yield, and quality components of two corn (L.) genotypes [J]. Agricultural Water Management, 2013, 128: 65–71
[39] Hammad H M, Ahmad A, Abbas F, et al. Water and nitrogen productivity of maize under semiarid environments[J]. Crop Science, 2015, 55: 877–888
[40] 王金涛. 制种玉米开花特性与籽粒数模拟及节水调质高效灌溉优化决策研究[D]. 北京: 中国农业大学, 2017Wang J T. Modeling flowering characteristics and kernel number and optimizing irrigation scheduling for improving water use efficiency and regulating seed quality in hybrid maize seed production[D]. Beijing: China Agricultural University, 2017
Effect of drought stress on maize phenology and yield components and its simulation*
MI Na1, ZHANG Yushu1, CAI Fu1, JI Ruipeng1**, FANG Yuan2, ZHANG Shujie1, CHEN Nina1
(1. Institute of Atmospheric Environment, China Meteorological Administration / Key Laboratory of Agrometeorological Disasters, Liaoning Province, Shenyang 110166, China; 2. Meteorological Cadre Training Institute, China Meteorological Administration (Liaoning), Shenyang 110166, China)
Drought is one of the main limiting factors for maize production in Northeast China, and drought stress is particularly severe during the seeding and flowering stages of the crop. The occurrence of drought around anthesis affects the temporal separation of male (anthesis) and female (silking) floral maturity (referred to as the anthesis-silking interval, ASI), which leads to a reduction in maize kernel numbers. To examine the effect of drought stress during the “yield critical period” on maize phenology, yield and yield components as well as to assess the modeling results of phenology and yield simulated by a crop growth model under drought stress, a field experiment was conducted. Sowing was performed on different dates at 10-day intervals beginning from April 20 over a 5-year period (2011–2015). Observation data of maize yield and yield components, particularly phonology and yield data in the drought years of 2014 and 2015 were used to analyze the effect of drought stress on maize phenology and yield components. The results of simulation using the CERES-Maize model were assessed in this study. Further improvements for the CERES-Maize model under drought conditions were proposed. The findings revealed that drought during the yield critical period delayed silking much greater than anthesis. More than 90% of the plants reached the anthesis stage, whereas only 45%–88% of the plants reached the silking stage. Drought in 2014 and 2015 reduced the kernel numbers of maize by 32% and maize yield by 33%-78%. Grain yield was significantly (< 0.001) and positively correlated with the number of kernels per ear. The CERES-Maize model showed good performance (normalized root mean square error (NRMSE) of yield simulation was 6.5% for 2012 and 10.8% for 2013) in the normal years, whereas its performance during the drought years was unsatisfactory (NRMSE of yield simulation was 21.6% for 2014 and 76.5% for 2015), which was attributed partly to the neglect of the longer ASI, failure to reach silking, or delayed phenology, causing a decrease in yield. Above all, drought stress during the yield critical period affected maize phonology (anthesis to silking stage) to some extent, affecting kernel number and grain yield. Thus, it is necessary to study the modeling of maize yield under drought stress based on ASI and silking dynamics by coupling the plant biomass framework.
Maize; Yield critical period; Drought stress; Anthesis to silking interval; Yield; Yield components; CERES-Maize model
P49
2096-6237(2019)12-1779-10

10.13930/j.cnki.cjea.190390
* 辽宁省自然科学基金指导计划(20180551169)、辽宁省科学技术厅农业攻关及成果产业化项目(2014210003)、国家自然科学基金面上项目(41975149, 41775110)和中央级公益性科研院所基本科研业务费项目(2018SYIAEHZ1, 2018SYIAEZD1)资助
纪瑞鹏, 研究方向为农业气象灾害评估、农业气候资源利用等。E-mail:jiruipeng@163.com
米娜, 主要研究方向为植物干旱响应与干旱影响评估。E-mail: mina7921@126.com
2019-05-23
2019-08-11
* This study was supported by the Natural Science Foundation of Liaoning (20180551169),the Agricultural Research and Achievements Industrialization Project of Liaoning Province (2014210003), the National Natural Science Foundation of China (41975149, 41775110) and the Central Public-Interest Scientific Institution Basal Research Fund of China (2018SYIAEHZ1, 2018SYIAEZD1).
, E-mail:jiruipeng@163.com
May 23, 2019;
Aug. 11, 2019
米娜, 张玉书, 蔡福, 纪瑞鹏, 方缘, 张淑杰, 陈妮娜. 干旱胁迫对玉米物候及产量组成的影响及模拟研究[J]. 中国生态农业学报(中英文), 2019, 27(12): 1779-1788
MI N, ZHANG Y S, CAI F, JI R P, FANG Y, ZHANG S J, CHEN N N. Effect of drought stress on maize phenology and yield components and its simulation[J]. Chinese Journal of Eco-Agriculture, 2019, 27(12): 1779-1788
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
中国农学通报(2022年13期)2022-05-31
现代畜牧科技(2021年4期)2021-12-05
商品与质量(2021年16期)2021-04-23
林业机械与木工设备(2021年2期)2021-03-11
——以长白山区为例
东北林业大学学报(2020年9期)2020-10-21
农业灾害研究(2020年8期)2020-05-11
意林(2018年18期)2018-09-30
作文周刊·小学一年级版(2017年35期)2017-10-18
