
夏秋季丹霞梧桐叶片光合特性日变化特征比较
2019-12-04 04:42:12黄华章戴文坛缪绅裕陶文琴陈健辉陈再雄陈志红
中国野生植物资源 2019年5期
黄华章,戴文坛,缪绅裕*,陶文琴,陈健辉,陈再雄,陈志红,胡 辉
(1. 广州大学 生命科学学院,广东 广州 510006;2. 韶关市丹霞山旅游投资经营有限公司,广东 仁化 512300;3. 广东南雄小流坑-青嶂山省级自然保护区管理处,广东 南雄 512400)
梧桐科梧桐属(FirmianaMarsili)的灌木或小乔木种类——丹霞梧桐(F.danxiaensisHuse et H.S.Kiu),迄今仅发现自然分布于广东省仁化县丹霞山和南雄市全安镇[1]的丹霞地貌山地,为中国特有种,已被列为国家Ⅱ级重点保护野生植物和野生极小种群保护计划[2]。目前对丹霞梧桐的研究侧重于其微卫星标记[3]和遗传多样性[4]、全新基因组序列组装[5]、空间分布的微地貌环境特征[6]及林冠层分布测定[7]、地理成分分析[8]及群落特征[9]、人工繁殖与野外回归实验[2]等,尚未涉及其生理生态学指标的测定及与生态因子的相关性研究。梧桐属中有许多种为地方特有种,为研究生态适应性和分化提供了材料,但其如何能适应岩石崖壁和土壤贫瘠胁迫下丹霞地貌山地的机制尚不明了[4]。为此,本研究拟通过测定丹霞梧桐夏秋季叶片的光合速率、蒸腾速率并分析其与相关生态因子的关系,探讨该植物对丹霞地貌中不同季节气温等生态因子的适应性,以期为该植物种群的生存和发展提供基础数据。
1 材料与方法
1.1 研究地自然环境概况
研究地点位于广东南雄全安镇苍石谷(114°12'14.112″E,25°07'30.738″,海拔201.3 m)。南雄市极端最高温37.4℃,极端最低温-1.4℃,年均温19.9℃,年降水量1 550 mm,年均日照时数1 825.7 h,土壤为紫红色厚层砂页岩、紫红色粉砂岩夹砾岩等[8]。
1.2 光合生理生态指标的测定
利用北京雅欣公司生产的Yaxin-1102型便携式光合蒸腾仪,分别于2018年夏季的7月23日(大暑日)和秋季的10月4日(晴到多云)中的8:00~17:00,每隔1 h 测定1次,选择健康成熟的丹霞梧桐植株中上部叶片3~6片,待系统稳定后,测定其净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、叶片温度(Tl)及主要环境因子,包括大气CO2浓度(Ca)、光合有效辐射(PAR)、气温(Ta)和大气相对湿度(RH)等参数。每个测定日,同步用日本产SPAD-502叶绿度测定仪随机测定3株丹霞梧桐共30枚成熟叶片的叶绿度。根据公式:叶片水分利用效率(Water Use Efficiency,WUE)=光合速率/蒸腾速率,计算其叶片的水分利用效率。
1.3 数据分析
用Excel 2007软件进行数据处理(如计算平均值和标准差等)并绘图,用SPSS 23.0软件进行数据统计与分析。
2 结果与分析
2.1 叶片SPAD值
夏季(7月23日)和秋季(10月4日)丹霞梧桐叶片SPAD测定值分别为38.58±3.69、36.72±3.62,经检验,二者之间不存在显著性差异。尽管丹霞梧桐为落叶小乔木,10月上旬已有不少植株许多叶片变黄(每年10月左右树叶变黄后脱落[6]),但测定日多数植株叶片依然维持绿色,所测定的SPAD值在秋季比夏季略有下降。
2.2 气温与大气相对湿度
夏季测定日(7月23日)平均气温(37.37±1.99℃)略高于秋季(10月4日)的33.78±3.43℃,大气相对湿度在夏季(65.66%±4.87%)显著高于秋季(41.97%±6.88%),即体现出丹霞梧桐生境在夏季具有高温高湿的特征。测定日中,7月最高气温出现在上午11:00(40.10℃),但秋季则出现在下午13:00(38.10℃)。测定日的夏季最高气温40.10℃和秋季最高气温38.10℃,均高于南雄市的极端最高温37.4℃[8]。
2.3 大气CO2浓度和胞间CO2浓度
测定日的平均大气CO2浓度在夏季(395.41±1.963 μmol CO2/mol)略高于秋季(383.72±14.22 μmol CO2/mol);胞间CO2浓度在夏季(268.50±55.88 μmol CO2/mol)也略高于秋季(244.80±72.75 μmol CO2/mol)。无论是夏季还是秋季,大气CO2浓度变化较平稳,且均显著高于胞间CO2浓度。
2.3 光合有效辐射
测定日的光合有效辐射日变化情况见图1。因晴到多云天气,致使7月23日的光合有效辐射在806.89~1339.22 μmol/m2·s之间,平均为1084.46±165.95 μmol/m2s,中午12:00测得1个相对低峰值859.89 μmol/m2·s。10月4日基本为晴天,测得的光合有效辐射在25.33~1233.56 μmol/m2·s之间,平均950.25±406.22 μmol/m2·s,在8:00~15:00之间数值较平稳,但在15:00之后因野外测定地在山谷,秋季日落早,导致山体遮蔽了太阳直射,光合有效辐射数值迅速下降,至17:00时已接近0,以致日平均值下降、变异幅度大增。
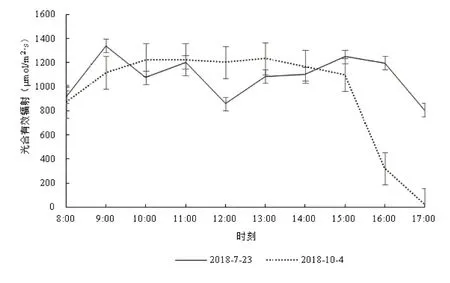
图1 不同测定日光合有效辐射的日变化Fig.1 Diurnal variation ofphotosynthetically active radiation in different determining days
2.4 叶片净光合速率
丹霞梧桐叶片净光合速率日变化的测定结果见图2。图2中可见,2个测定日的净光合速率曲线均呈现为多峰曲线,7月23日的净光合速率在9.71~24.18 μmol CO2/m2·s之间,平均为14.35±4.56 μmol CO2/m2·s,最大值24.18 μmol CO2/m2·s出现在下午14:00。10月4日测得的净光合速率在0.69~16.99 μmol CO2/m2·s之间,平均8.70±5.16 μmol/m2·s,最大值16.99 μmol CO2/m2·s出现在上午10:00。夏季和秋季2个测定日叶片净光合速率平均值比较,7月23日略大于10月4日。
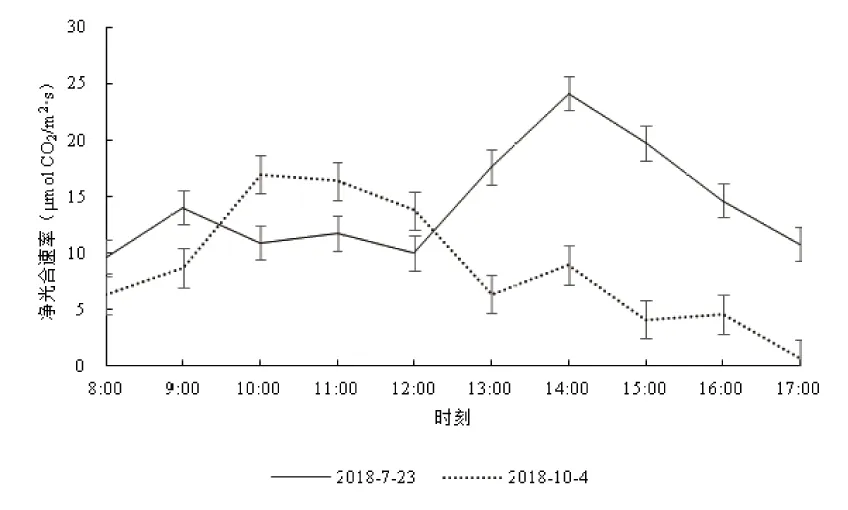
图2 不同测定日丹霞梧桐叶片净光合速率的日变化Fig.2 Diurnal variation of netphotosynthetic rates of F. danxiaensis leaf in different determining days
2.5 叶片蒸腾速率
丹霞梧桐叶片蒸腾速率日变化的测定结果见图3。图3中可见,2个测定日的蒸腾速率曲线均呈现为多峰曲线,7月23日的蒸腾速率在1.52~8.13 mmol H2O/m2·s之间,平均为4.28±1.97 mmol H2O/m2·s,最大值8.13 mmol H2O/m2·s出现在上午11:00;第2高峰7.10 mmol H2O/m2·s出现在下午14:00。10月4日测得的蒸腾速率在0.05~5.05 mmol H2O/m2·s之间,平均2.75±1.34 mmol H2O/m2·s,最大值5.05 mmol H2O/m2·s出现在上午10:00,第2高峰4.03 mmol H2O/m2·s出现在中午12:00。夏季和秋季2个测定日叶片蒸腾速率平均值比较,7月23日大于10月4日。
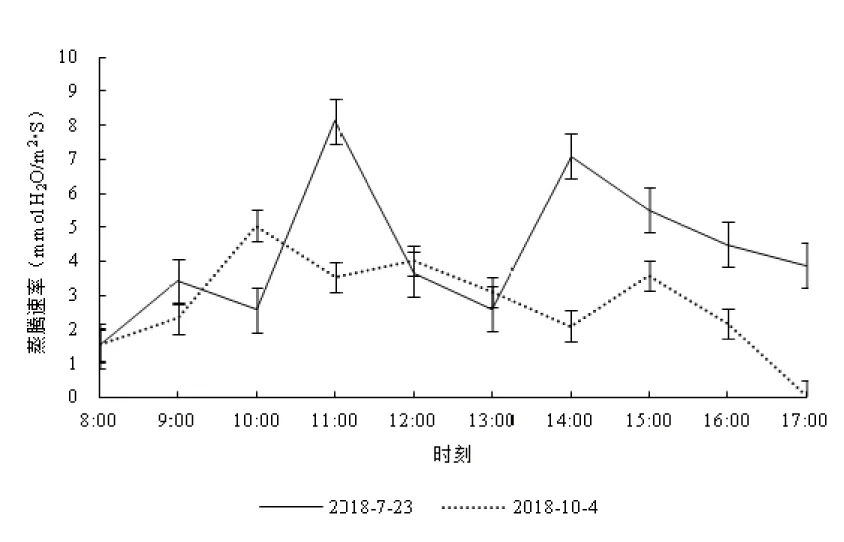
图3 不同测定日丹霞梧桐叶片蒸腾速率的日变化Fig.3 Diurnal variation oftranspiration rates of F. danxiaensis leaf in different determining days
根据叶片净光合速率和蒸腾速率计算出的水分利用效率,在夏季(7月23日)平均为3.88±1.63,比秋季(10月4日)的4.26±3.51略低。
2.6 叶片气孔导度
丹霞梧桐叶片气孔导度日变化的测定结果见图4。图4中可见,2个测定日的叶片气孔导度曲线均呈现为较典型的双峰曲线,尤其是在夏季。7月23日的气孔导度在67.13~348.47 mol H2O/m2·s之间,平均为184.33±86.00 mol H2O/m2·s,最大值348.47 mol H2O/m2·s出现在下午14:00;第2高峰322.77 mol H2O/m2·s出现在上午11:00;最小值67.13 mol H2O/m2·s出现在下午13:00,具有明显的午休现象。10月4日测得的气孔导度在4.80~128.50 mol H2O/m2·s之间,平均85.19±34.44 mol H2O/m2·s,最大值128.50 mol H2O/m2·s出现在上午10:00,第2高峰120.93 mol H2O/m2·s出现在下午15:00。夏季和秋季2个测定日叶片气孔导度平均值比较,7月23日显著大于10月4日。
2.7 光合特性及其与生境因子的相关性
对各生理生态指标与生境因子相关性进行分析,结果见表1。表1中可见,在夏季仅有叶片蒸腾速率(Tr)与气孔导度(Gs)达到极显著正线性相关,蒸腾速率与气温达到显著线性相关。而在秋季,净光合速率(Pn)与蒸腾速率达到极显著正相关,与光合有效辐射(PAR)和气孔导度均达到显著的正相关;蒸腾速率还与气孔导度、气温(Ta)达到极显著正相关,而与光合有效辐射达到显著正相关;光合有效辐射还与气温达极显著正相关,而与气孔导度、大气相对湿度达到显著正相关;气温和大气相对湿度(RH)则呈极显著的负相关。
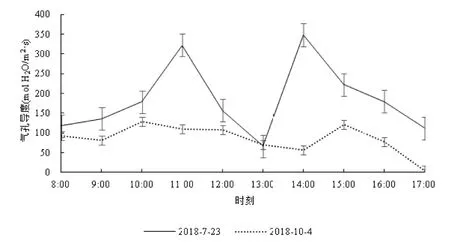
图4 不同测定日丹霞梧桐叶片气孔导度的日变化Fig.4 Diurnal variation of stomatal conductance ofF. danxiaensis leaf in different determining days
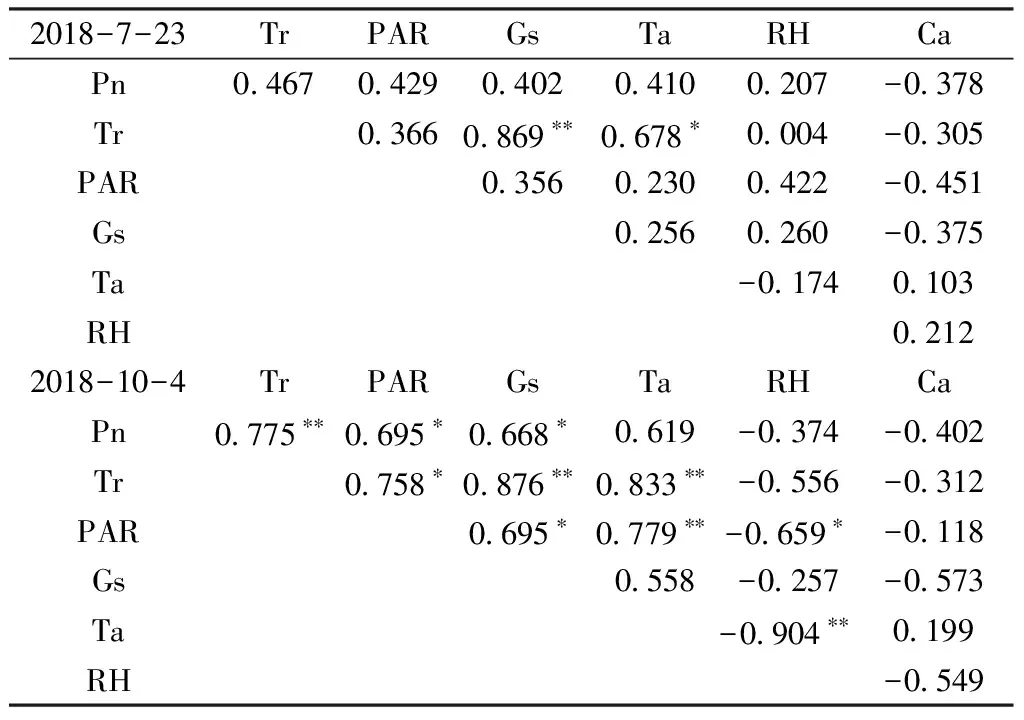
表1 丹霞梧桐叶片光合特性与生境因子相关系数
注:自由度df=8时,0.05水平显著相关的临界值是0.632;0.01水平极显著相关临界值是0.765
3 结论与讨论
光合作用的日变化是外界环境条件相互作用,以及环境条件与植物本身因素相互制约的综合表现,反映了生产过程中物质积累、生理代谢对环境因子的不同适应性[10]。植物光合作用的日变化一般分为“单峰”和“双峰”2种曲线类型[11],但本研究所测得的夏、秋季丹霞梧桐叶片净光合速率、蒸腾速率的日变化均呈不规则的多峰曲线,可能与测定日的生态因子(如光合有效辐射等)的不规则变化有关,尽管在夏季净光合速率与光合有效辐射的之间未达到显著相关,而在秋季它们之间则显著相关。秋季丹霞梧桐的净光合速率与蒸腾速率、光合有效辐射、气孔导度均呈显著或极显著的正相关,这与前人研究小叶女贞(LigustrumquihouiCarr.)的结果[12]较一致。本研究结果夏季(7月)比秋季(10月)的平均净光合速率要大,且净光合速率与其他生态生理因子的相关性呈现夏季相关性不大,秋季则显著相关的规律,与白及[Bletillastriata(Thunb. ex A. Murray) Rchb. f.]的净光合速率6月>8月>10月;6月、10月光合速率与光合有效辐射、气孔导度呈正相关,而8月不呈显著相关的变化规律[13]相似。丹霞梧桐夏季的净光合速率大于秋季,这与同为国家保护植物的耐荫树种四药门花(TetrathyriumsubcordatumBenth.)最大净光合速率在夏季的6.10 μmol CO2/m2·s,高于冬季的5.55 μmol CO2/m2·s[14]的变化情况一致。
净光合速率的多峰曲线也会出现在其他植物中,例如不同种源的山杏[Armeniacasibirica(L.) Lam.]光合速率、气孔导度和蒸腾速率日变化都具有单峰、双峰和多峰3种类型[15]。国家保护植物喜树(CamptothecaacuminataDecne.)的净光合速率日变化为不对称的双峰曲线,最大峰值出现在上午8:00左右(11.31 μmol CO2/m2·s)[16]。丹霞梧桐净光合速率的峰值在7月为24.18 μmol CO2/m2·s,10月为16.99 μmol CO2/m2·s,7月的峰值略高于阳生的云杉(PiceaasperataMast.)的21.58 μmol CO2/m2·s和高山松(PinusdensataMast.)的21.57 μmol CO2/m2·s[17];夏、秋季的峰值均高于阳生的小叶女贞(14.14 μmol CO2/m2·s)和广东含笑(MicheliaguangdongensisY. H.)(13.46 μmol CO2/m2·s)[18]。丹霞梧桐蒸腾速率在7月8.13 mmol H2O/m2·s,10月5.05 mmol H2O/m2·s,均高于小叶女贞蒸腾速率的峰值4.91 mmol H2O/m2·s,可见丹霞梧桐的净光合速率、蒸腾速率等均比一般的阳生植物高,体现出对丹霞地貌山谷特殊生境的适应性较强;且夏季各光合特性均高于落叶前的秋季,也体现出落叶树种比常绿树种因生长周期相对短而充分利用生长季中的高温高湿的夏季。丹霞山的沟谷地带小气候相对封闭,与开阔区域相比保持了高湿环境,但林内气温则比空旷地略低[18]。丹霞梧桐较耐旱,为适应在土壤较贫瘠的崖壁上生存,其树干挺拔,根系特别发达,能最大限度地吸收水分存活下来[6]。本研究结果提示,尽管丹霞梧桐对自然生境有较好的适应性,但因其经济价值(优良的园林绿化树种[8]和树干可作钢琴用材)而受到一定的生存威胁[2],低的遗传多样性可能源自其受限的自然分布区域和强烈的丹霞地貌土壤(低的土壤肥力和强酸性)的选择压力[2,4],因而保护丹霞梧桐的生境是维护该极小种群生存和发展的最好方法,减少其被人为破坏,在就地保护的同时要加强对其进行迁地和离体保护。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
东方少年·布老虎画刊(2023年1期)2023-08-02 08:34:24
热带亚热带植物学报(2022年6期)2022-12-17 07:12:04
节水灌溉(2021年5期)2021-05-26 06:49:38
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
阅读(低年级)(2021年10期)2021-03-08 12:49:39
节水灌溉(2020年9期)2020-09-16 00:32:12
中国农业科技导报(2020年12期)2020-03-15 10:50:08
乡村地理(2018年4期)2018-03-23 01:53:40
