
从江香猪SIM1 基因SNP 位点遗传效应分析
2019-12-04 08:30毛世明陈大芳龙清孟冯文武李明勇尹杨莎唐明宗
养猪 2019年6期
李 俊,毛世明,陈大芳,龙清孟,周 迪,李 平,张 勇,张 雄,冯文武,李明勇,尹杨莎,唐明宗
(1.贵州省种畜禽种质测定中心,贵州 贵阳 550021;2 贵州大学动物科学学院,贵州 贵阳 550025;
3.贵州省畜牧兽医研究所,贵州 贵阳 550021)
SIM1(Single-minded 1)基因是一种编码与神经发生有关的转录因子,是碱性-螺旋-环-螺旋(basic helix-loop-helix, bHLH)转录因子超家族中新发现的亚家族成员之一[1],通过影响下丘脑视上核和室旁核神经内分泌细胞在哺乳动物能量平衡调节中的作用,从而影响动物的采食行为,可导致食欲过盛、肥胖和体重增加[2]。Holder 等[3]研究发现SIM1基因平衡易位会导致儿童肥胖。Tolson 等[4]发现SIM1+/-和SIM1-/-小鼠食量增加,比同窝正常小鼠肥胖,且催产素(Oxytocin)和黑素皮质素激素(MC4R)在下丘脑内的表达量显著降低;由此,Traurig 等[5]推测SIM1基因可能参与瘦素—黑素皮质激素—催产素这一调控途径,进而导致肥胖,并通过DNA 测序发现SIM1基因一处非编码区突变位点与印第安人的体脂指数显著相关。Zhao 等[6]应用辐射杂交细胞系(RH)将SIM1基因定位到猪染色体1p13,并且在SIM1基因内含子七的607 bp(A/G)和SIM1外显子八的780 bp(C/T)处检测到两个多态位点,发现该突变和生长率、背膘厚、眼肌面积、腿部脂肪重、日增重以及肌内脂肪含量显著相关。因此推测,SIM1基因可能是影响猪胴体性状、生长性状和肉质性状的潜在功能基因。本文采用PCR 产物直接测序法,对已筛选到的SIM1基因3 个SNP 位点进行验证和基因分型,分析该基因对从江香猪胴体性状的遗传效应,为进一步利用分子标记选育提纯从江香猪奠定理论基础。
1 试验与方法
1.1 试验时间与地点
试验于2018 年3 月至2019 年6 月在贵州省贵阳市乌当区羊昌镇绿生源香猪生态福利养殖场和中心实验室进行。
1.2 试验材料
1.2.1 试验动物 试验样品为74 头从江香猪耳组织,采自贵阳市乌当区绿生源香猪生态福利养殖场。用耳号钳取耳组织1.5 g,标记后分别放入装有1 mL 70%乙醇的2 mL 离心管中,带回实验室-20 ℃保存,用于DNA 提取。
1.2.2 主要试剂及仪器 组织基因组DNA 提取试剂盒、2×Taq Master Mix、核酸染料、琼脂糖、DL 2 000 DNA Marker 等试剂均购自上海生工生物工程技术服务有限公司;去离子水、无水乙醇、TAE 缓冲液等试剂实验室自备。PCR 扩增仪(C1000 Touch),凝胶成相系统(G:BOX F3),紫外分光光度计(NanoDrop-ONE),去离子纯水仪(Milli-Q Intergral 3)等。
1.3 试验方法
1.3.1 DNA 提取 用基因组DNA 提取试剂盒提取DNA,1%琼脂糖凝胶电泳检测,凝胶成像系统照相保存,并用紫外分光光度计测量每个DNA 样品浓度3 次,取其平均值,用ddH2O 将所有DNA 样品调至100ng/μL,-20℃保存备用。
1.3.2 引物设计与PCR 扩增 参照GenBank(登录号为NC_010443.5)提供的猪SIM1基因序列,以及李俊等[7]利用DNA 池在从江香猪群体中筛查到SIM1基因的12 个SNP 位点,运用NCBI 中的Primer-BLAST 设计引物,委托赛默飞世尔科技(中国)有限公司合成,引物序列及扩增条件见表1。

表1 SIM1 基因引物序列、退火温度、扩增长度及区域
1.3.3 PCR 反应体系及条件 采用25 μL PCR 反应体系,即DNA 模板2.5 μL,2×Taq Master Mix 12.5 μL,上、下游引物各1.5 μL,ddH2O 7 μL。PCR 反应条件为:95 ℃预变性4 min;94 ℃变性30 s,退火30 s,72 ℃延伸1 min,共34 个循环;72 ℃再延伸10 min;最后得到的PCR 产物4 ℃保存,1%琼脂糖凝胶电泳检测PCR 产物,凝胶成像系统照相保存,将检测合格样品送英潍捷基(上海)贸易有限公司测序。
1.3.4 统计分析 测序得到的DNA 序列用DNAStar等软件进行序列对比,筛选SNP 位点,计算不同位点在受试群体中的基因型频率、等位基因频率、有效等位基因数(Ne)、群体遗传纯合度(Ho)、杂合度(He)和多态信息含量(PIC);根据最小二乘线性模型,分析不同基因型与样本胴体性状的关联效应。
2 结果与分析
2.1 PCR 产物检测
PCR 扩增产物经1%琼脂糖凝胶电泳检测,均显示为一条特异性条带,其片段长度P1、P2、P3 分别为193 bp、194 bp、173 bp,与目的片段相符,条带清晰、整齐无拖尾,可用于测序(图1)。
2.2 测序结果分析
运用SeqMan 软件对所有样品的测序结果分析发现(图2),C77T 位点分型为MM、MN、NN 基因型;C225T 位点分型为CC、CT、TT 基因型,A426G 位点分型为AA、AG、GG 基因型。通过序列对比发现,MM 基因型、CC 基因型、AA 基因型个体的序列与GenBank上提交的序列相一致,定义为野生型,NN 基因型、TT 基因型、GG 基因型定义为突变型,MN 基因型、CT 基因型、AG 基因型为杂合型。

图1 部分PCR 产物电泳检测结果

图2 SIM1 基因3 个SNP 位点测序结果
2.3 SNP 位点的基因型频率及遗传特异性分析
从群体遗传学角度分析从江香猪SIM1基因SNP 位点的基因型和等位基因频率(表2)。从表2可看出,对于C77T 位点,MM 基因型频率最高,表现为优势基因型;M、N 等位基因频率分别为87.8%、12.2%,且M 为优势等位基因。对于C225T 位点,CC基因型频率最高,表现为优势基因型;C、T 等位基因频率分别为76.3%、23.7%,且C 为优势等位基因。在A426G 位点上,AA 基因型和AG 基因型频率一样,均为35.1%,A、G 等位基因频率分别为52.6%、47.4%,且A 为优势等位基因。
通过对C77T、C225T、A426G 位点的遗传结构进行分析,均发现这3 个SNP 位点在试验群体中纯合度(Ho)较高,杂合度(He)则相对较低,表明这3 个SNP 位点在群体中的变异较小;3 个位点的有效等位基因数(Ne)分别为1.27、1.56、1.99;而A426G 位点最接近于实际检测到的2 个等位基因数,说明该位点的等位基因在群体中分布最均匀;3 个位点的多态信息含量(PIC)分别为0.19、0.30、0.37,表明C77T 位点表现为低度多态(PIC<0.25),C225T、A426G 位点变现为中度多态(0.25<PIC<0.50),经χ2适合性检验结果表明,C77T、A426G、C225T 位点均未偏离Hardy-weinberg 平衡(P>0.05)。

表2 从江香猪SIM1 基因3 个SNP 位点基因型频率、等位基因频率及遗传特异性分析
2.4 连锁不平衡分析与单倍型构建
利用SHEsis 在线软件对SIM1基因3 个多态位点进行连锁不平衡参数估计及单倍型构建,见表3和表4,结果表明,r2和D'大于0,说明3 个多态位点间分别处于连锁不平衡状态。可构建7 种单倍型,即:M-C-G、M-C-A、M-T-A、N-C-A、M-T-G、N-T-A、N-C-G;分别命名为H1、H2、H3、H4、H5、H6、H7,其频率分别为39.3%、28.2%、13.0%、8.1%、7.2%、3.4%、0.7%。
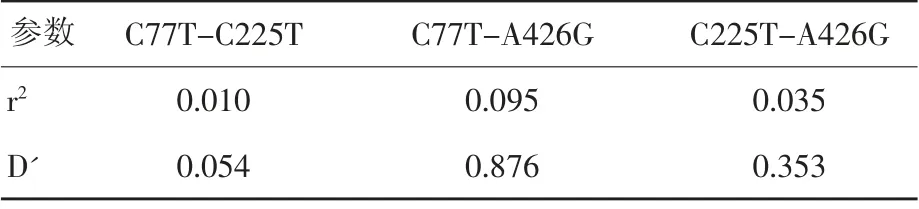
表3 3 个SNP 位点之间的连锁不平衡参数估计

表4 3 个SNP 位点的单倍型构建及频率
2.5 SIM1 基因不同基因型与胴体性状的关联性分析
从江香猪SIM1基因C77T 位点、C225T 位点、A426G 位点不同基因型与肌内脂肪、眼肌面积、背膘厚、皮厚的最小二乘分析结果见表5。由表5 可知,在C77T 位点上,MM 基因型的肌内脂肪和眼肌面积高于MN 和NN 基因型,但差异不显著(P>0.05),背膘厚为NN 基因型<MN 基因型<MM 基因型,且NN 基因型与MM 基因型差异显著(P<0.05),皮厚为NN 基因型最低。对于C225T 位点,肌内脂肪和眼肌面积均为CC 基因型高于CT 基因型和TT 基因型,背膘厚和皮厚均为TT 基因型低于CC 基因型和CT基因型,且差异均不显著(P>0.05)。对A426G 位点,肌内脂肪和眼肌面积为GG 基因型>AG 基因型>AA基因型,但差异不显著(P>0.05),背膘厚和皮厚均为AA 基因型最低,且AA 基因型的背膘厚显著低于GG 基因型(P<0.05)。

表5 不同基因型与胴体性状的关联性分析
通过SIM1基因的3 个SNP 位点不同基因型的聚合,产生15 种聚合基因型,采用最小二乘法分析与聚合基因型与胴体性状的关联性(表6),结果显示,肌内脂肪含量最高的为MMCCAA 组合基因型,最低的为MMTTAA 组合基因型;眼肌面积最高的为MNCCGG 组合基因型,最低的为MNTTAA 组合基因型;背膘厚和皮厚最低的均为MNTTAA 组合基因型,背膘厚和皮厚最高的均为MNCCGG 组合基因型;但不同聚合基因型间的肌内脂肪、眼肌面积、背膘厚、皮厚差异均不显著(P>0.05)。
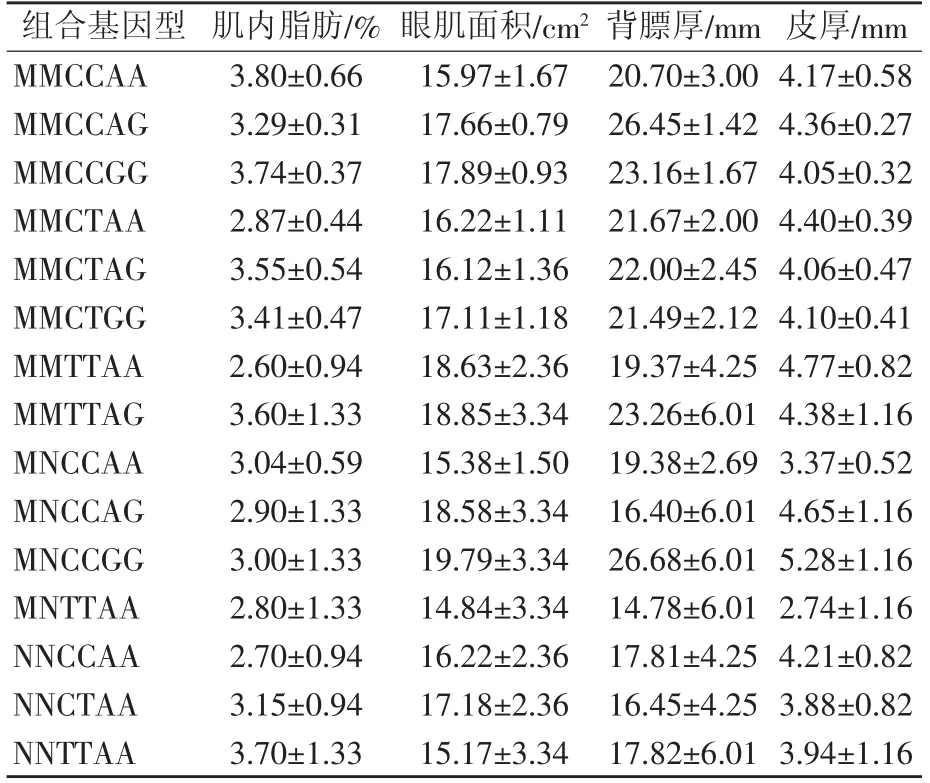
表6 3 个SNP 位点聚合基因型与胴体性状的关联性分析
3 讨论
SIM1基因是人类和鼠肥胖症的候选基因,缺失SIM1基因可导致视前核神经元细胞不发育,影响哺乳动物能量平衡和采食行为。Holder 等[8]报道,一名患者1 号和6 号染色体之间发生易位,破坏6 号染色体上的SIM1基因,导致出现肥胖症。而小鼠试验研究[4]显示,SIM1基因纯合子小鼠在出生后不久就死亡,而杂合子小鼠能够存活下来且出现食欲过盛和肥胖的症状,可见SIM1基因在维持动物能量、食欲和体脂方面起着重要的作用。但目前关于SIM1基因的研究主要在人和小鼠上,在家畜家禽方面的研究较少。郭晓令等[9]对169 头国内外猪种的SIM1基因SNP 分析发现,国外猪种均存在CC、CT、TT 基因型,且纯合基因型个体的背膘厚显著大于杂合基因型个体;但在国内猪种中只发现TT 基因型,这与陈哲等[10]在金皮F2 代资源家系猪SIM1基因发现CC、CT 和TT 3 种基因型不一致,推测其原因可能是品种差异或者SNP 位点的不同造成,且TT 基因型个体的背膘厚高于CC 基因型个体的背膘厚。Nezer等[11]、Beeckmann 等[12]和De 等[13]均在猪SIM1基因所在区域发现一个影响背膘厚、采食量和生长率的数量性状基因座(Quantitative Trait Locus, QTL)。由此可见,SIM1基因对猪的脂肪沉积具有显著效应。
本研究通过对74 头从江香猪SIM1基因扩增的PCR 产物全部测序发现,与李俊等[7]利用DNA 池筛选出的SIM1基因C77T、C225T、A426G 位点结果相一致,证实DNA 混池筛查出的SNP 位点也具有一定的可靠性。而多态性分析发现3 个SNP 位点分别产生3 种不同的基因型,其中MM 基因型、CC 基因型、AA 基因型表现为优势基因型,且M、C 和A 表现为优势等位基因。遗传结构分析显示3 个SNP 位点在该群体中分布均匀,且能比较合理地提供遗传信息。因此,通过连锁不平衡分析发现,上述3 个SNP 位点两两之间均处于连锁不平衡状态。一个群体中单倍型的频率不能小于0.03%,因此单倍型频率未达到0.03%的未参与统计,对应的单倍型组合也应忽略,SHEsis 在线软件在单倍型分析过程中,系统会自动删除不符合条件的单倍型组合[14-15],从而构建7 种单倍型,其中H1、H2 和H3 为主要单倍型。由于该研究的试验样本较少,发现的SNP 位点少,有些单倍型所占比例太小,对试验结果造成一定的误差,未进行单倍型与胴体性状的关联性分析,因此,还需进一步扩大试验样本加以验证。
关联性分析发现,C77T、C225T、A426G 位点不同基因型以及聚合基因型与肌内脂肪、眼肌面积、皮厚之间差异均不显著(P>0.05),C77T、A426G 位点不同基因型从江香猪背膘厚差异显著(P<0.05)。MM基因型、CC 基因型、GG 基因型的肌内脂肪和眼肌面积高于其他基因型;NN 基因型、TT 基因型、AA 基因型的背膘厚和皮厚低于其他基因型;肌内脂肪最高的为MMCCAA 聚合基因型,眼肌面积最高的为MNCCGG 聚合基因型;背膘厚和皮厚最低的均为MNTTAA 聚合基因型。但由于本试验样本数少、品种单一,具有一定局限性。因此,在今后的研究中,将扩大样本数,并在肌内脂肪、眼肌面积、背膘厚、皮厚差异较大品种中验证这3 个SNP 位点是否对从江香猪胴体性能产生影响,为今后推动从江香猪高产肉性能品系的选育提供参考。
4 结论
本试验发现SIM1基因C77T、C225T、A426G 位点可能是影响从江香猪胴体性能的3 个分子标记,可以将MM 基因型、CC 基因型、GG 基因型、MMCCAA聚合基因、MNCCGG 聚合基因型作为影响从江香猪肌内脂肪和眼肌面积的有利基因型;将NN 基因型、TT 基因型、AA 基因型和MNTTAA 聚合基因型作为影响从江香猪背膘厚和皮厚有利基因型。
猜你喜欢
畜牧兽医学报(2022年12期)2023-01-03
川北医学院学报(2022年6期)2022-06-24
农技服务(2021年1期)2021-12-06
智慧健康(2021年17期)2021-07-30
养猪(2020年2期)2020-04-14
养猪(2019年3期)2019-06-19
晚晴(2016年6期)2016-05-14
中国诗歌(2015年6期)2015-11-15
青少年科技博览(中学版)(2015年10期)2015-01-11
文史天地(2009年1期)2009-02-18
