
10株猪流行性腹泻病毒序列分析
2019-11-28 01:31杨小蓉赵晨璐周建民
中国兽医杂志 2019年7期
杨小蓉,马 良,许 磊,赵晨璐,周建民
(1.中牧实业股份有限公司,北京 丰台100070;2.农业部兽用生物制品与化学药品重点实验室,北京 海淀 100095;3.北京市兽用多肽疫苗设计与制备工程技术研究中心,北京 海淀 100095)
猪流行性腹泻(PED)是由猪流行性腹泻病毒(PEDV)引起猪的一种急性、高度接触性、致死性肠道传染病,临床特征为腹泻、呕吐、脱水,各种年龄段和品系的猪群均易感[1]。该病于1971 年首次被报道后,相继在亚洲及南美暴发,给养猪业造成了严重的经济损失,我国以CV777 疫苗株研制的商品化弱毒疫苗可有效防控PED 的流行。但2010 年秋冬PED 在中国多个省份暴发并迅速波及到全国以来,呈高发病率、高死亡率的特点[2]。近几年PED流行范围不断扩大,成为严重危害中国养猪业的传染病之一[3]。
PEDV 基因组为线性单股正链RNA 病毒,属于尼多病毒目(Nidovirales)冠状病毒科(Coronaviridae)甲型冠状病毒属(Alphacoronavirus)成员[1]。PEDV 的S 蛋白,在识别靶细胞以及病毒和细胞膜融合过程中发挥重要作用,能刺激机体产生中和抗体,在抗病毒免疫机制中发挥着重要作用[4]。S 蛋白目前认为是可用于监测PEDV 的变异及流行趋势的基因,也与疫苗的有效性密切相关[5]。本研究对2015-2018 年北方地区流行的PED 病原的S 基因进行了测序,与国内外参考毒株进行遗传变异分析,构建了进化树,了解PEDV 的变异趋势,以期为疫病防控和疫苗研制提供参考。
1 材料与方法
1.1 病料来源 2015-2018 年来自黑龙江、吉林、北京、山东、河北、山西、河南等北方省份发病猪场采取的病料。
1.2 试剂材料 病毒RNA 提取试剂盒、病毒RNA反转录试剂盒、高保真DNA 聚合酶和dDNP 等试剂,均购自北京全式金生物技术有限公司。
1.3 病毒RNA 的提取及S 基因RT-PCR 扩增测序 采用Viral RNA kit 试剂盒进行病毒核酸的提取纯化,将提取的RNA 采用First-Stand cDNA kit试剂盒进行反转录,获得cDNA。
根据GenBank 发表的PEDV 序列设计了5 对PEDV S 基因扩增引物。PCR 反应体系:10×Buffer 5 μL,dNTPs(2.5 mmol/L)4 μL,上下游引物各1μL,cDNA 2 μL,高保真Taq酶1 μL。PCR 反应条件:94 ℃4 min,94 ℃30 s,55 ℃30 s,72 ℃30 s,共34 循环;72 ℃8 min。
用1%琼脂凝胶电泳对PCR 扩增产物进行观察,然后对PCR 阳性产物进行回收,送北京铂尚公司进行测序。
1.4 序列拼接与分析 应用DNAStar、ClustalX、GeneDoc 等生物学软件进行S 基因的拼接,与Gen-Bank 收录的国内外参考毒株进行比对与分析。用于本试验分析的毒株见表1。
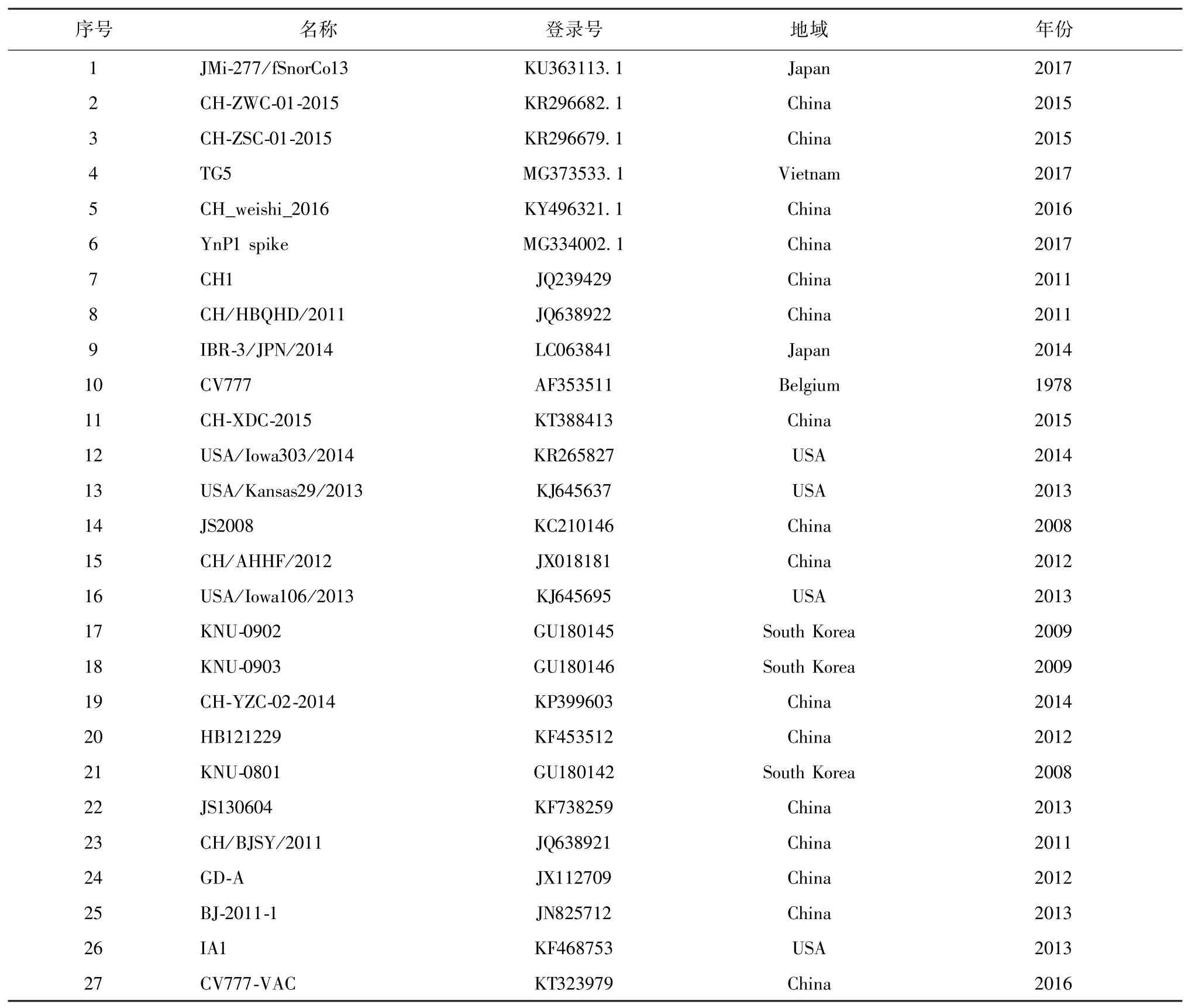
表1 试验中所用的PEDV 毒株及来源
2 结果与分析
2.1 S 基因核苷酸序列及其推导编码的氨基酸序列同源性分析 通过应用软件分析发现,本试验获得的10 个PEDV S 基因序列相互之间核苷酸相似性为96.8%~99.4%,其推导的氨基酸相似性为96.5%~99.3%。其中,CH-HBhs1803、CH-SDbz1804、CH-SDdy1804、CH-SXyc1803、CH-HBcz-1702 和 CH-HNny-1611 共6 株序列基因长度均为4 158 nt,编码1 385 个氨基酸;CH-SDbz1803、CH-BJCP1509、CH-HLJqqhe1803和CH-JLSP-1504 共4 株序列基因长度均为4 161 nt,编码1 386 个氨基酸。

表2 S 基因核苷酸及推导编码的氨基酸序列与国内外参考毒株同源性比较 /%
2.2 依据S 基因推导编码的氨基酸序列绘制遗传进化树 利用MEGA 6 软件对所获得的10 个S 基因和国内外具有代表性的27 个毒株序列进行多序列比较,构建系统进化树(见图1)。
从进化树图谱中显示,PEDV 流行毒株依据S基因推导的氨基酸构建进化树,可以分为G1 群和G2 群。G1 可以进一步分为以CV777 为代表毒株的经典G1a 亚群,以及以CH-ZWC-01-2015 和USA/Iowa106/2013 为代表的G1b 亚群。G2 亚群可分为G2a 和G2b 两个亚群,其中G2b 亚群可进一步分为以G2ba 亚群和G2bb 亚群。与国内外参考毒株相比,本研究扩增得到的8 个S 基因序列位于以YnP1 spike、CH1 和GD-A 为代表的G2bbb 亚群,而CHHLJqqhe1803 位于以CH-YZC-02-2014 为代表的G2bba 亚群,CH-JLSP-1504 位于以CH_weishi_2016为代表的G2ba 亚群。
10 个S 基因序列与国内流行毒株CH-XDC-2015亲缘性更近,推导编码的氨基酸与CH-XDC-2015 相似性为97.7%~99.4%;与疫苗株CV777-VAC 亲缘性较远,推导编码的氨基酸与CV777-VAC 相似性为92.0%~93.1%;与经典株CV777 亲缘性较远,推导编码的氨基酸与CV777 相似性为92.6%~93.7%。由此说明病毒在流行过程中,发生了变异,与疫苗株相似性较低。

图1 依据S 基因推导的氨基酸绘制进化树
2.3 S 基因推导编码氨基酸序列的变异分析 本试验共得到10 个S 基因序列,与亲缘性更近的国内流行毒株CH-XDC-2015 进行序列比对,其推导编码的氨基酸序列存在缺失和点突变。尤其存在特征性的氨基酸缺失,CH-HBcz-1702 毒株存在131 位I缺失;CH-HBhs1803、CH-SDbz1804、CH-SDdy1804、CH-SXyc1803 和CH-HNny-1611 共5 个毒株存在1 196 位N 缺失。与CH-XDC-2015、CV777 等流行毒株相比,10 个毒株在核心表位COE 结构域(499~638 位氨基酸区域,有中和表位)存在氨基酸变异。

表3 S 蛋白氨基酸核心表位COE 结构域的变异
3 讨论
S 蛋白是PEDV 的一种结构蛋白,通过S 蛋白能把病毒粒子吸附在宿主细胞受体上,然后通过膜融合渗进细胞中,以刺激宿主诱导产生中和抗体,因此S 蛋白具有PEDV 抗原的作用[5-6]。而PEDV的核心表位(COE)存在于S 蛋白的编码基因中,被认为是PEDV 的亚单位疫苗的重要靶标抗原[4]。
基于S 基因核苷酸序列分析,2015-2018 年国内发生的PEDV 可分为3 个亚群,其中8 个S 基因序列位于以YnP1 spike、CH1 和GD-A 为代表的G2bbb 亚群,而CH-HLJqqhe1803 位于以CH-YZC-02-2014 为代表的G2bba 亚群,CH-JLSP-1504 位于以CH_weishi_2016 为代表的G2ba 亚群。结合核苷酸及其推导的氨基酸变异分析发现,S 基因变异株已经成为优势流行毒株。其中6 株S 基因推导的氨基酸序列已分别在131 位、1196 位出现1 个氨基酸缺失。10 个毒株在核心表位COE 结构域(499-638位氨基酸区域,有中和表位)均存在不同程度的氨基酸变异。10 个S 基因序列在进化树中已形成多个小分支,相对来说与国内流行毒株CH-XDC-2015亲缘性更近,其推导编码的氨基酸与以CH-XDC-2015 为代表的变异株相似性为97.7%~99.4%;与疫苗株CV777-VAC 亲缘性较远,推导编码的氨基酸与CV777-VAC 相似性为92.0%~93.1%;与经典株CV777 亲缘性较远,推导编码的氨基酸与CV777相似性为92.6%~93.7%。由此说明目前养殖场暴发的PEDV 在流行过程中,已经发生了变异,与疫苗株相似性程度较低。目前,经典毒株CV777 灭活疫苗对于我国PEDV 疫病的防控起着重要作用。而S 基因序列变异株的出现,则可能导致现有疫苗失去保护作用[7]。因此,监测PEDV 的S 基因变异趋势,对于制定防控策略和疫苗开发均有重要指导意义。
猜你喜欢
动物医学进展(2022年9期)2022-11-26
中国医药科学(2022年5期)2022-05-05
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
天津市教科院学报(2021年5期)2021-11-10
山东医药(2020年19期)2020-12-29
科学导报(2020年25期)2020-04-28
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
腹腔镜外科杂志(2016年9期)2016-06-01
