
瘦素受体OB-RGRP水平对脂肪细胞的影响及其作用机制研究
2019-11-23 08:54石卫红李小林巫国辉
中国美容医学 2019年11期
石卫红 李小林 巫国辉

[摘要]目的:为了探讨不同水平的瘦素受体(OB-RGRP)调节大鼠脂肪细胞的研究及其作用机制,明确其对脂肪细胞的影响,为瘦素受体治疗肥胖症提供理论依据。方法:原代分离大鼠脂肪细胞,利用油红O染色进行鉴定,构建过表达OB-RGRP和干扰OB-RGRP载体,利用Western blot验证细胞转染效果,并检测JAK2、STAT3及其磷酸化的表达情况。结果:细胞分离正确,构建的OB-RGRP-siRNA转染有效,与对照组相比,过表达OB-RGRP组的p-JAK2和p-STAT3表达明显升高,干扰OB-RGRP组的p-JAK2和p-STAT3表达明显下降,差异具有统计学意义(P<0.05)。而JAK2、STAT3总蛋白各组之间无差异。结论:OB-RGRP水平通过JAK2/STAT3信号通路在调节瘦素抵抗中起着重要作用,调节脂肪代谢,有助于改善瘦素抵抗及肥胖防治。
[关键词]OB-RGRP;瘦素受體;Western blot;JAK2/STAT3信号通路;脂肪细胞;肥胖症
[中图分类号]R329.2+8 [文献标志码]A [文章编号]1008-6455(2019)11-0090-03
Abstract: Objective To investigate the mechanism of leptin receptors(OB-RGRP) in regulating obesity In order to investigate the regulation of different levels of leptin receptor on adipocytes in rats and its mechanism of action, and to clarify its effect on adipocytes, so as to provide theoretical basis for the treatment of obesity by leptin receptor. Methods Primary rat adipocytes were isolated and identified by oil red O staining. OB-RGRP overexpressed and OB-RGRP interfering vectors were constructed. The cell transfection effect was verified by Western blot, and the expression of JAK2, STAT3 and their phosphorylation were detected. Results The cells were isolated correctly and the constructed OB-RGRP-siRNA was effective in transfection. Compared with the control group, the overexpression of p-JAK2 and p-STAT3 in the OB-RGRP group was significantly increased, while the expression of p-JAK2 and p-STAT3 in the OB-RGRP group was significantly decreased, and the difference was statistically significant (P <0.05).There was no difference between JAK2 and STAT3 groups. Conclusion OB-RGRP level plays an important role in regulating leptin resistance through the JAK2/STAT3 signaling pathway, regulating fat metabolism and contributing to the improvement of leptin resistance and the prevention and treatment of obesity.
Key words: OB-RGRP; leptin receptor; Western blot; JAK2/STAT3 signal path; adipocyte; adiposis
随着经济的发展和人们生活水平的提高,肥胖问题越来越受到关注,发达国家肥胖率显著高于发展中国家,而我国近年来的肥胖率也显著升高[1-2]。据报道,我国成人肥胖率由1989年的1.32%迅速增长到2009年的9.62%[3]。肥胖是由多种因素决定的,过度饮食,超范围的热量摄取,运动量不足以及一些疾病或遗传等因素均可能导致肥胖。除去外在的饮食与运动因素,肥胖还与基因的调控有关,瘦素(1eptin)是由肥胖基因(OB gene)编码,脂肪组织分泌的激素具有抑制食欲、调节能量代谢,通过结合瘦素受体(1eptin receptor,OB-RGRP)发挥其调节脂肪蛋白质等能量代谢的作用[4-5]。因此机体的瘦素水平或是瘦素受体的水平对肥胖起到关键作用。
本研究通过构建瘦素受体(OB-RGRP)过表达载体和干扰载体,转染至大鼠脂肪细胞,通过Western blot检测相关蛋白的表达,探索不同瘦素受体水平对大鼠脂肪细胞的影响及其可能的作用机制和调节方式。
1 材料和仪器
1.1 实验动物:SD大鼠购于湖南斯莱克景达实验动物有限公司,许可证号:SCXK(湘)(2016-0002)。SD大鼠饲养:采用无毒塑料鼠盒,不锈钢丝笼盖,金属笼架。笼架可移动,并可经受多种消毒方法灭菌。
1.2 实验试剂与仪器:DMEM(1×)+GlutaMAXTM-1 Dulbeccos Modified Eagle Medium(gibco 1859228);Lipofectamine? 3000(invitrogen 18882752);OB-RGRP-si-RNA(通用生物BIO33001);I型胶原酶(Solarbio C8150);油红染色试剂盒(KGA329,KeyGen);EcoRI(ER0271,Thermo Scientific);XhoI(ER0691,Thermo Scientific);Mouse Monoclonal Anti-GAPDH(TA-08,中杉金桥,1/2 000);辣根酶标记山羊抗鼠IgG(H+L)(ZB-2305,中杉金桥,1/2 000);Rabbit Polyclonal Anti-OB-RGRP(别名:LEPR)(DF7139,Affinity,1/500);Rabbit Monoclonal Anti-JAK2 (ab108596,abcam,1/5 000);Rabbit Monoclonal Anti-JAK2 (phospho Y1007+Y1008)(ab32101,abcam,1/1 000);Rabbit Monoclonal Anti-STAT3 (phospho S727) (ab32143,abcam,1/1 000);Rabbit Polyclonal Anti-STAT3 (A1192,ABclonal,1/500);倒置荧光显微镜(广州市明美光电有限公司MF53);显微镜(CX41 OLYMPUS);蛋白垂直电泳仪(DYY-6C,北京市六一仪器厂);超高灵敏度化学发光成像系统(Chemi DocTM XRS+,伯乐生命医学产品(上海)有限公司)。
1.3 原代分离大鼠脂肪细胞:采用断颈处死SD大鼠,并迅速浸入75%的酒精中,消毒3min。将SD大鼠置于无菌冰板上,使用75%酒精棉球擦拭腹部,采用低位横向切口打开大鼠腹腔。用镊子夹住腹股沟处的脂肪,手术剪将其剪下。将取出的脂肪放于无菌的培养皿中,用含10%的双抗D-PBS液反复清洗数次,再使用无菌手术剪仔细剔除附着的组织。把清理好的脂肪组织转移至新的培养皿中,并用无菌的手术剪将脂肪组织剪成糜状。把剪成糜状的脂肪组织用吸管吸至10ml的离心管中,加入I型胶原酶使其工作浓度为0.1%,并吹打均匀。将离心管置于37℃水浴中消化75min,期间每隔15min吹打1次,具体可视情况而定。加入含20% FBS的DMEM/F-12培养基终止消化。200目筛网过滤,收集滤液,1 800rpm离心5min,留上层油脂。用含20%胎牛血清、1×双抗的DMEM培养基重悬上层油脂。将重悬液铺板,放在37℃、5% CO2饱和湿度的培养箱内培养。
1.4 油红O染色:去除培养基,加入4%的组织细胞固定液固定15min;蒸馏水充分清洗,60%异丙醇充分浸润,油红染色液染色8min,去除染色液,60%异丙醇快速分化,蒸馏水清洗,苏木素染色2min,去除染色液,水洗,然后风干,甘油明胶封片,显微镜观察。
1.5 OB-RGRP过表达载体和干扰载体的构建:NCBI查找OB-RGRP基因序列,引入酶切位点(EcoRI/XhoI)生物合成基因片段[克隆至pcDNA3.1(+)载体上]。利用软件分析设计出最佳的靶序列,同时参考文献上所用的靶序列,将潜在的靶序列和基因组数据库进行比对,排除和其他基因编码序列同源的序列,设计出siRNA,再交由通用生物系统(安徽)有限公司合成。基因序列见表1。
1.6 细胞转染:根据实验需要对细胞铺12孔板,与细胞传代方法相同,将细胞根据实验需要进行稀释,每孔约8×104个细胞,均匀铺到细胞培养板中,做好标记,放置培养箱中培养,待细胞完全贴壁后,且当细胞密度达70%时,准备转染。将细胞的培养基更换为不含血清的培养基,体积为0.5ml;取灭菌的EP管2个,每管加125μl Opti-MEM,其中一管加入5μl lipofectamine 3000,另一个EP管加入12.5μl siRNA(siRNA干粉使用DEPC水溶解;125μl/1OD),混匀后室温孵育5min;将上述两个EP管混匀,室温孵育15min,将混合液分成两等份滴到12孔板中对应的孔内,将细胞放回孵箱培养;转染4h后在12孔板中加入血清含量为20%的完全培養基1ml;48h后进行PCR验证。
1.7 实验分组:①大鼠脂肪细胞(正常培养细胞,Control);②大鼠脂肪细胞+过表达空载(转染过表达空载48h Vector);③大鼠脂肪细胞+OB-RGRP过表达载体(转染过表达OB-RGRP 48h,OB-RGRP);④大鼠脂肪细胞+干扰空载(转染过表达si-OB-RGRP NC 48h,si-OB-RGRP NC);⑤大鼠脂肪细胞+OB-RGRP RNAi慢病毒载体(转染过表达si-OB-RGRP 48h,si-OB-RGRP)。
1.8 Western blot:取各组细胞加入相应的裂解液中,4℃裂解30min,在10 000rpm/min离心10min,小心吸取上清液,即得总蛋白。利用BCA试剂盒进行蛋白浓度测定。蛋白变性、上样、电泳1~2h,湿法转膜30~50min。4℃孵育一抗溶液过夜;室温孵育二抗1~2h。在膜上滴加ECL曝光液,曝光。用“Quantity one”软件分析各抗体条带灰度值。
1.9 統计学分析:所有数据均采用SPSS 19.0进行统计分析,经t检验判定显著性差异,以P<0.05为差异有统计学意义。
2 结果
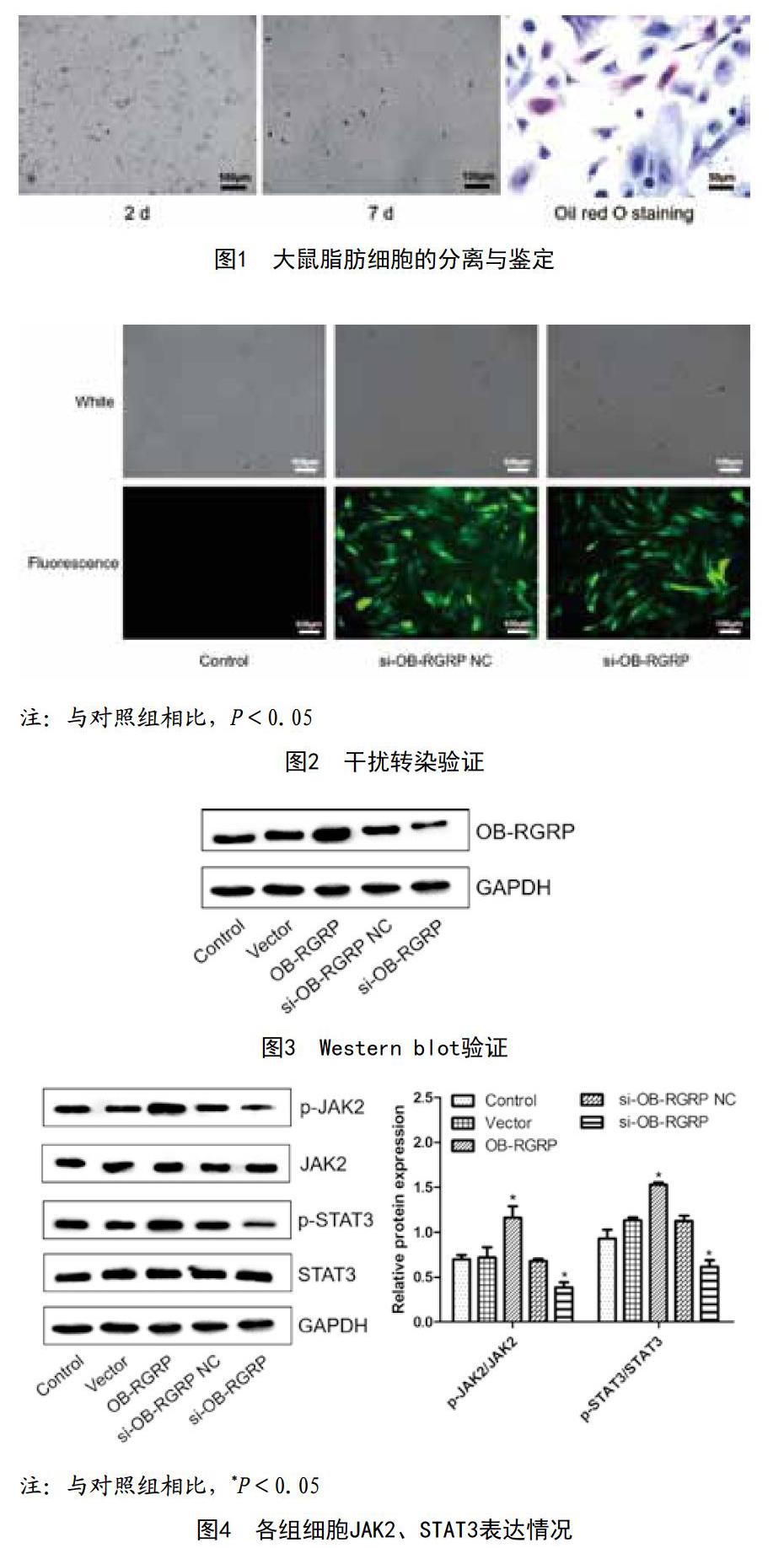
2.1 大鼠脂肪细胞的分离与鉴定:脂肪细胞刚接种时呈类圆形,悬浮于培养基中,2d后细胞开始伸展,逐渐成多角梭形;7d左右进入指数增长期,呈漩涡状。经油红O染色,胞浆内的反光颗粒着橘红色,因此鉴定细胞为脂肪细胞。见图1。
2.2 干扰验证:细胞转染后与对照组相比,si-B-RGRP NC组、si-B-RGRP组绿色荧光表达显著增多,表明这两个组分成功转染si-B-RGRP NC与si-B-RGRP,慢病毒干扰载体构建成功。见图2。
2.3 干扰与过表达Western blot验证:与对照组相比,过表达OB-RGRP组的OB-RGRP表达明显升高,干扰OB-RGRP组的OB-RGRP表达明显下降。见图3。
2.4 各组细胞JAK2、STAT3表达情况:JAK2、STAT3总蛋白各组之间无差异,但与对照组相比,过表达OB-RGRP组的p-JAK2/JAK2与p-STAT3/STAT3表达明显升高,干扰OB-RGRP组的p-JAK2/JAK2与p-STAT3/STAT3表达明显下降,差异有统计学意义(P<0.05)。见图4。
3 讨论
肥胖患者机体瘦素水平较正常人较低或是瘦素受体表达量较低,导致瘦素与瘦素受体不能够有效的结合,从而不能够有效地调节体内的能量代谢,导致肥胖相关的症状[6]。本研究成功分离大鼠原代脂肪细胞,并通过油红O染色鉴定为脂肪细胞,构建OB-RGRP过表达载体与干扰siRNA并成功验证其有效。通过Western blot检测JAK2、STAT3及其磷酸化水平确定OB-RGRP的功能及其不同水平对机体的影响。
JAK2/STAT3信号转导通路具有广泛的生理作用,参与机体的生长,发育和免疫活动,近年来许多研究发现其与肥胖疾病关系密切,有研究表明机体瘦素通过介导JAK2/STAT3信号通路参与脂肪和能量代谢,促进机体分解和利用各种能量物质,维持自身代谢平衡[7-8]。根据本研究结果瘦素受体(OB-RGRP)水平也能通过介导JAK2/STAT3信号通路实现对肥胖的调控,过表达的OB-RGRP促进JAK2/STAT3的磷酸化,致使抑制瘦素调控脂肪代谢,诱发肥胖[9-10],而低表达的OB-RGRP能够使JAK2/STAT3的磷酸化降低,从而促进瘦素调控脂肪代谢,抑制肥胖的发生[11-12]。
瘦素受体有六种异形体,广泛存在于下丘脑、骨骼肌及脂肪组织等。有研究表明,瘦素可以透过血脑屏障直接作用于下丘脑内的瘦素受体,将外周能量存贮相关信号反馈至下丘脑[13],下丘脑调节能量平衡的通路为刺鼠相关蛋白(agouti-related peptide,AgRP)神经元/阿黑皮素原(proopiomelanocortin,POMC)神经元通路,维持能量平衡[14-15]。因此瘦素或瘦素受体的不足,也可能会抑制AgRP/POMC通路,导致脂肪能量不能正常消耗而产生肥胖。
肥胖的诱发因素多种多样,因人而异,因此关于肥胖慢性疾病的产生机理众说纷纭。目前笔者只对JAK2/STAT3信号通路做了简要的研究,更加错综复杂的调节机制还有待于进一步研究和阐明。
[参考文献]
[1]贺媛,赵小兰,曾强,等.城市成人超重、肥胖、中心性肥胖的流行特征和相关危险因素分析[J].实用预防医学,2015,22(4):390-394.
[2]Mcpherson NO,Lane M.Male obesity and subfertility,is it really about increased adiposity?[J].Asian J Androl,2015,17(3):450-458.
[3]倪国华,张璟,郑风田.中国肥胖流行的现状与趋势[J].中国食物与营养,2013,19(10):70-74.
[4]孙志娟,黄之瑜.肥胖的研究进展[J].生理科学进展,2001,32(2):39-44.
[5]马敬弟,卢姗,马向华,等.脂肪细胞因子与肥胖的关系[J].南京医科大学学报(自然科学版),2012,12(12):1743-1745.
[6]马晓洁,陈杨,郭建玲,等.奥氮平诱导性肥胖大鼠糖脂代谢紊乱和脂肪源性细胞因子网络调控研究[J].精神医学杂志,2014,27(2)93-96.
[7]Ernst M,Najdovska M,Grail D,et al.STAT3 and STAT1 mediate IL-11-dependent and inflammation-associated gastric tumorigenesis in gp 130 receptor mutant mice[J].J Clin Invest,2008,118(5):1727-1738.
[8]Galien R.Janus kinases in inflammatory bowel disease:four kinases for multiple purposes[J].Pharmacol Rep,2016,68(4):789-796.
[9]Gurzov EN,Stanley WJ,Pappas EG,et al.The JAK/STAT pathway in obesity and diabetes[J].FEBS J,2016,283(16):3002-3015.
[10]Roxburgh CS,McMillan DC.Therapeutics targeting innate immune/inflammatory responses through the interleukin-6/JAK/STAT signal transduction pathway in patients with cancer[J].Transl Res,2016,167(1):61-66.
[11]柳成荫,刘倩男,赵剑,等.膳食诱导肥胖小鼠睾丸JAK2/STAT3蛋白表达变化研究[J].实用预防医学,2018,25(12):1413-1416.
[12]Zhao J,Zhai L,Liu Z,et al.Leptin level and oxidative stress contribute to obesity-induced low testosterone in murine testicular tissue[J].Oxid Med Cell Longev,2014(12):190-194.
[13]Van Swieten MM,Pandit R,Adan RA,et al.The neuroanatomical function of leptin in the hypothalamus[J].J Chem Neuroanat,2014(61):207-220.
[14]Wauman J,Tavernier J.Leptin receptor signaling:pathways to leptin resistance[J].Front Biosci( Landmark Ed),2011,16:2771-2793.
[15] Lee B,Kim SG,Kim J,et al.Brain-specific homeobox factor as a target selector for glucocorticoid receptor in energy balance[J].Mol Cell Biol,2013,33(14):2650-2658.
[收稿日期]2019-03-12
本文引用格式:李小林,巫國辉,万冠群,等.瘦素受体(OB-RGRP)水平对脂肪细胞的影响及其作用机制研究[J].中国美容医学,2019,28(11):90-93.
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
纺织服装周刊(2022年13期)2022-04-15
中国信息化周报(2019年8期)2019-03-27
商情(2018年9期)2018-03-29
考试周刊(2018年8期)2018-01-19
江苏农业科学(2017年17期)2017-11-15
中国市场(2017年5期)2017-03-15
商情(2009年17期)2009-09-23
销售与管理(2006年9期)2006-09-17
