
超量表达CsGH3.6通过抑制生长素信号转导增强柑橘溃疡病抗性
2019-11-19 11:19:34邹修平龙俊宏彭爱红陈敏龙琴陈善春
中国农业科学 2019年21期
邹修平,龙俊宏,彭爱红,陈敏,龙琴,陈善春
超量表达通过抑制生长素信号转导增强柑橘溃疡病抗性
邹修平,龙俊宏,彭爱红,陈敏,龙琴,陈善春
(西南大学/中国农业科学院柑桔研究所国家柑桔品种改良中心,重庆 400712)
由柑橘黄单胞杆菌柑橘亚种(subsp,)引起的柑橘溃疡病是柑橘生产上最具毁灭性的一种病害。植物生长素在调控柑橘溃疡病菌引起的寄主侵染部位脓疱形成中起重要作用。生长素早期响应基因通过酰基化吲哚-3-乙酸(indole-3-acetic acid,IAA)调控植物激素动态平衡。前期研究发现柑橘在调控生长素响应溃疡病侵染中起着重要作用。【】通过对超量表达转基因晚锦橙的抗病性、植株表型、细胞和激素变化进行分析,利用RNA-Seq解析调控的信号通路,探明调控激素动态平衡影响柑橘溃疡病抗性的内在机制。利用针刺法对离体转基因叶片接种溃疡病菌,统计接种第10 天时病斑面积和病情指数,以野生型为对照,评价转基因植株的抗性水平;提取感病前后叶片内源激素,利用高效液相色谱技术(high performance liquid chromatography,HPLC )检测转基因植株中激素含量变化;温室中观察转基因植株表型变化;通过测量叶片纵径、横径和厚度分析转基因植株叶型变化特征;制备叶片表皮切片,显微观察表皮细胞和气孔,并统计转基因植株表皮细胞长度和气孔密度;采用RNA-Seq测序技术研究转基因植株转录组变化情况,并利用Nr、Nt、Pfam、COG、SwissProt和gene ontology (GO)数据库注释基因功能,进一步利用KEEG数据库和MapMan软件解析超量表达影响的重要基因、功能和途径,阐明调控柑橘溃疡病抗性的分子机制。超量表达显著增强转基因植株的溃疡病抗性;转基因植株分枝增多且下垂,叶片向上卷曲,变小,颜色浅;转基因植株气孔密度增加,表皮细胞变短;激素含量分析显示,转基因植株自由生长素(IAA)和茉莉酸(jasmonic acid,JA)含量显著降低,而水杨酸(salicylic acid,SA)含量显著增加;转录组测序分析表明,超量表达显著抑制生长素信号转导相关基因表达,特别是所有注释的Aux/IAA家族基因均下调表达,相反,与生物胁迫相关基因的表达为上调,其中绝大部分基因为病程相关蛋白基因。超量表达通过酰基化自由IAA抑制生长素信号转导,调控JA和SA的动态平衡,改变细胞和植株的形态建成,从而增强柑橘对溃疡病的抗性。研究结果暗示调控激素动态平衡在柑橘抗病育种中具有潜在价值。
柑橘黄单胞杆菌柑橘亚种;柑橘溃疡病;生长素;;抗病性
0 引言
【研究意义】柑橘溃疡病(citrus bacterial canker,CBC)是一种危害极严重的世界性病害,其病原菌为柑橘黄单胞杆菌柑橘亚种(subsp.,)。感染柑橘溃疡病的植株在叶、枝和果实上出现火山状病斑,严重时落叶、枯枝、落果,产量降低、果实品质变劣,一旦疫情在柑橘产区传播蔓延,对柑橘产业具有毁灭性、灾难性打击[1-2]。柑橘产业中大部分栽培品种属于溃疡病易感品种。因此,大力开展柑橘溃疡病抗(感)机理和重要基因资源挖掘的研究,对柑橘优良抗性新品种的选育和柑橘产业的健康稳定发展具有重要意义,也将进一步深化对病原菌致病机理、植物与病原互作机制的理解。【前人研究进展】植物生长素在调控柑橘溃疡病菌侵染引起的脓疱形成中起重要作用。Costacurta等[3]研究发现,柑橘溃疡病菌能分泌吲哚-3-乙酸(indole-3-acetic acid,IAA),而且甜橙叶片提取液能促进病原菌繁殖和IAA合成;Cernadas等[4]研究表明,柑橘溃疡病菌侵染促进甜橙生长素合成、运输和信号转导相关基因的转录。进一步研究发现,萘乙酸(NAA)处理促进甜橙感病部位水浸状脓疱的形成,而生长素抑制剂1-N-萘基邻氨甲酰苯甲酸(NPA)抑制溃疡病症状的发展[5-6],显示生长素具有促进柑橘溃疡病症状发展的作用。目前,关于植物生长素介导寄主感病的机制研究主要集中在拟南芥、水稻等植物中[7]。病原菌通过向寄主侵染部位细胞分泌IAA或增强寄主IAA的合成,致使寄主细胞内IAA的水平急剧上升,高浓度的IAA使植物细胞壁pH下降,胞壁结构蛋白酸化,细胞壁发生重排,从而引起细胞壁松弛,细胞扩张和膨大,以利于病原菌的入侵和扩散[8-9]。而且,侵染部位高浓度的IAA抑制水杨酸(salicylic acid,SA)介导的抗病反应[7,10-11]。面对病原菌对生长素的操控,植物必然启动相应的对抗机制来维持感病部位细胞激素的动态平衡,努力消除病原菌的危害。其中,生长素响应因子GH3起着重要作用[7-8]。编码蛋白具有植物激素酰胺合成酶活性,催化自由IAA与氨基酸结合,从而使其失活,当植物需要IAA时,通过IAA酰胺水解酶水解作用释放出IAA,以此调控细胞内IAA动态平衡[12-13]。另外,一些GH3也具有酰胺化SA和茉莉酸(jasmonic acid,JA)等激素的活性[14]。的功能研究主要集中在拟南芥和水稻中,在调控植物抗病性反应中起重要作用[12,15]。目前认为参与植物抗病反应的机制为:病原菌侵染导致寄主感病部位细胞IAA水平急剧上升,寄主通过生长素受体TIR1感知上升的IAA水平并触发生长素信号途径,迅速激活表达,GH3酶将过量的IAA酰胺化,使活性IAA水平下降,进而限制植物细胞壁增生和松弛,从而正向增强寄主的抗性[9,16-17]。比如超量表达和能增强水稻对白叶枯病和稻瘟病抗性[16-17],而能赋予植物广谱抗性[9]。随着基因组、转录组、蛋白组等大规模组学技术的应用,GH3家族基因在豆类、苹果、玉米、苜蓿、番茄、棉花等作物中的功能研究已在开展,其在生物和非生物抗性、激素动态平衡、信号途径中的作用备受关注[18-23]。但在柑橘中,鲜有关于GH3家族基因参与调控植物生长发育和抗性的研究报道。【本研究切入点】前期研究发现[24],柑橘生长素早期响应基因在调控生长素响应溃疡病菌侵染中起着重要作用,超量表达显著降低柑橘对溃疡病的感病性,但其机制尚待解析。【拟解决的关键问题】通过分析超量表达对转基因植株溃疡病抗性、激素水平、生长发育和转录组的影响,阐明调控柑橘溃疡病抗性的机制,为进一步解析柑橘溃疡病菌侵染中生长素途径调控寄主抗性的机理打下基础,为柑橘抗病育种提供新思路和新材料。
1 材料与方法
试验于2015—2018年在中国农业科学院柑桔研究所国家柑桔品种改良中心完成。
1.1 材料
供试植物材料为超量表达转基因晚锦橙()和野生型晚锦橙[24]。转基因植株以及野生型晚锦橙种植于中国农业科学院柑桔研究所国家柑桔品种改良中心育种圃。育种圃的温度控制在25—28℃,相对湿度控制在60%—70%,光周期L﹕D= 16 h﹕8 h。
1.2 转基因植株表型观察与叶型指数分析
转基因植株种植在田间2年后,观察其表型。选取完全成熟的春稍,从顶部开始选取第3—5节间的叶片,使用直尺测量叶片的纵径和横径,利用游标卡尺测量叶片的厚度。
1.3 表皮细胞观察
上午采摘完全伸展的成熟叶片,将离主脉0.5 cm处的叶片剪成0.5 cm×1 cm长方形,迅速放入3 mol·L-1的NaOH溶液中70℃水浴15 min,去除溶液,无菌水冲洗两次,再用蒸馏水冲洗4次。将叶片下表皮粘在透明胶带上,用手术刀片去尽残余叶肉组织。将粘有表皮细胞的透明胶带剪下,置于载玻片上制成临时切片,于光学显微镜下观察,并在40×物镜和10×目镜(放大倍数为400倍)条件下成像。试验重复3次,每次每个株系检测3片叶。使用ImageJ1.47软件统计气孔的数目和表皮细胞长度。气孔密度=视野范围内的气孔数/视野范围面积(个/mm2)。
1.4 激素含量测定
柑橘生长素IAA、水杨酸、茉莉酸含量测定参照Marques等[25]的方法。采摘1 g鲜重叶片,液氮速冻,研磨成粉;加入5 ml 80%甲醇浸提过夜,然后13 000×离心10 min。倒掉上清液,加入1ml 1%乙酸重悬沉淀。重悬液按照Oasis cartridges(Waters,美国马萨诸塞州)的方法进行纯化,纯化后的激素溶解于100 µL 10%的甲醇。提取的激素送钟鼎生物公司(南京)进行HPLC检测。
1.5 转基因柑橘的抗病性评价
以转基因晚锦橙和野生型晚锦橙的成熟叶片为材料,利用针刺接种法[26]接种溃疡病菌。具体方法:采集转基因和野生型晚锦橙的成熟叶片,清水洗净,70%乙醇杀菌3—5 s,再用无菌水清洗干净。将叶片背面向上平铺在无菌培养皿中,培养皿底部铺上湿润滤纸保湿,叶柄处用湿棉花覆盖保湿,每一片叶背面针刺12个小孔,用移液枪吸取1 µL(5×104CFU/mL)溃疡病菌悬浮液滴加到小孔处。培养皿用Parafilm封口膜密封,置于28℃光照培养箱培养。接种后第10天拍照,用软件Image J 1.47 统计病斑面积(mm2)。病斑面积的大小分为8个级别,分级方法如下:以字母R表示病斑面积,0级(R≤0.25 mm2),1级(0.25 mm2<R≤0.5 mm2),2级(0.5 mm2<R≤0.75 mm2),3级(0.75 mm2<R≤1 mm2),4级(1 mm2<R≤1.25 mm2),5级(1.25 mm2<R≤1.5 mm2),6级(1.5 mm2<R≤1.75 mm2),7级(R>1.75 mm2)。病斑面积分级后,根据以下公式统计病情指数(disease index,DI):DI=100×Σ(各级病斑数×相应级数值)/(病斑总数×最大级数)。根据病情指数,分析转基因植株抗溃疡病情况。
1.6 转录组测序分析
取转基因和野生型植株的叶片,液氮速冻,送北京百迈克科技股份有限公司进行转录组测序和信息学分析。试验设置3个生物学重复。以甜橙基因组序列(http://citrus.hzau.edu.cn/orange/index.php)为参考,利用TopHat2软件对测序数据进行比对分析。使用FPKM计算基因的表达水平。以野生型为对照,采用DESeq2软件进行表达差异显著性分析,差异表达基因(differential expressed gene,DEG)的筛选标准为错误发现率(false discovery rate,FDR)<0.01 且差异倍数 |log2fold change|>1。
利用Nr(non-redundant protein database,非冗余蛋白数据库)、Nt(NCBI non-redundant nucleotidedatabase,非冗余核苷酸数据库)、SwissProt(SwissProt protein database,蛋白质序列数据库)、COG(Cluster of Orthologous Groups,蛋白质直系同源数据库)、Pfam(Protein families database,蛋白质家族域数据库)、GO(Gene Ontology,基因本体论数据库)、KEGG(Kyoto Encyclopedia of Genes and Genomes,东京基因与基金组百科全书)数据库对基因进行功能注释。
为了详细分析调控的代谢途径和基因,对获得的转录组数据进一步进行MapMan功能注释(http://mapman.gabipd.org/web/guest/mercator)。以|log2fold change|>1且-value校正值padj<0.05为显著性标准可视化MapMan途径和功能,并使用Benjamin-Hochberg方法(FDR≤0.05)对MapMan途径和功能进行Wilcoxon双尾检测,筛选显著富集的途径、功能和基因[27]。
1.7 数据分析
试验结果均为3次重复的平均值,用Excel 2016进行数据整理、标准偏差计算及图表的绘制,差异显著性用SPSS 20.0统计软件进行分析。
2 结果
2.1 转基因植株溃疡病抗性评价
利用叶片离体接种技术对两年生转基因晚锦橙植株进行溃疡病抗性评价。接种溃疡病菌10 d时,统计病斑面积和病情指数(图1)。结果显示,野生型植株病斑面积为2.67 mm2,4株(b7、b13、b19和b20)转基因株系病斑面积分别为1.52、2.48、1.21、2.22 mm2,其中,b7和b19的病斑面积显著小于野生型(图1-A)。b7、b13、b19和b20株系的病情指数分别为45.55、89.12、44.48、77.02,而野生型植株的病情指数为83.48,转基因株系b7和b19的病情指数显著低于野生型对照(图1-B)。结果表明超量表达能显著提高柑橘抗溃疡病的能力。根据抗性评价结果,以下研究主要以b7和b19株系为材料开展。

WT:野生型Wild type;b7、b13、b19和b20:转基因植株Transgenic plant。下同 The same as below
柱上不同小写字母表示差异显著(Tukey’s检验,<0.05)。图2同 Different lowercases on the bars represent significant differences (Tukey’s test,<0.05). The same as Fig. 2
图1转基因植株溃疡病抗性评价
Fig. 1 Resistance evaluation oftransgenic plants to citrus bacterial canker
2.2 转基因植株自由IAA含量变化
为了分析超量表达对自由IAA含量的影响,利用HPLC检测在正常情况和溃疡病诱导条件下转基因株系 b7和b19的IAA含量。结果显示,病原菌诱导前转基因植株中自由IAA浓度显著低于野生型植株(图2)。接种溃疡病菌3 d时,野生型与转基因植株中自由IAA含量均显著下降,转基因植株自由IAA含量依然显著低于野生型植株。结果表明,超量表达不但抑制自由IAA的积累,而且促进病原菌侵染引起的自由IAA的下降。
2.3 转基因植株SA和JA含量变化
为了分析对植株体内SA和JA含量变化影响,检测了野生型与转基因植株中SA和JA的含量。结果表明,与野生型相比,转基因株系b7和b19中SA的含量显著升高,而JA的含量显著降低(图3)。
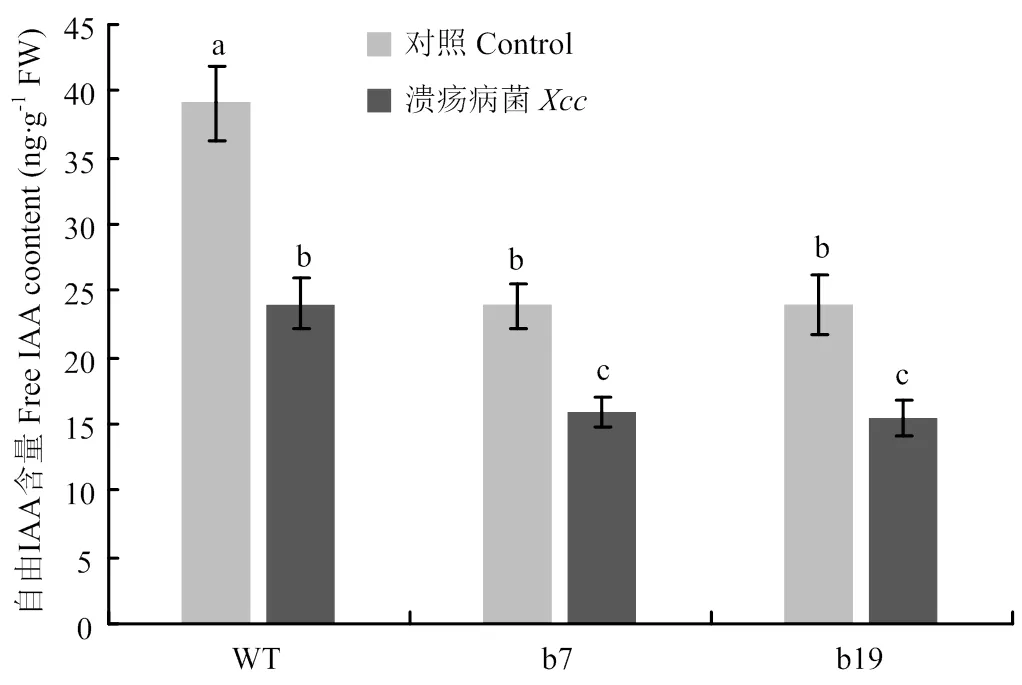
Control:溃疡病菌侵染前自由IAA含量检测Free IAA content detection before Xcc inoculation;Xcc:溃疡病菌侵染3 d时自由IAA含量检测Free IAA content detection at 3 d after Xcc inoculation

*表示与野生型(WT)相比差异显著(Tukey’s检验,p<0.05)。图4、图6同
2.4 超量表达CsGH3.6转基因晚锦橙的表型变化
两年温室表型观察发现,与野生型相比,b7转基因株系出现轻微卷叶;b19转基因株系叶片明显向上卷曲,叶片变小,颜色浅,整片叶子较野生型叶片明显下垂,茎较软易弯曲,植株呈枯萎状态(图4-A)。叶型指数统计分析表明,转基因植株叶片的纵径(图4-B)和横径(图4-C)以及叶片厚度(图4-D)均显著小于野生型。
2.5 转基因植株叶片表皮细胞变化
进一步分析了转基因株系b7和b19表皮细胞和气孔的变化(图5)。结果显示,转基因植株的表皮细胞长度显著降低(图6-A),气孔密度显著增加(图6-B)。
2.6 转录组分析
为了进一步探讨与柑橘溃疡病抗性的关系,对抗性水平最高的转基因株系b19进行了转录组测序分析。聚类热图分析表明,b19株系中基因表达谱与野生型相比有明显的差异(图7-A)。GO等功能注释共获得31 035个Unigene基因。与野生型相比,b19株系中有1 456个差异表达基因(DEG),940个DEG上调表达,516个DEG下调表达(图7-B)。KEGG富集分析显示(图7-C),148个DEG明显富集于20个途径和功能组,其中29个基因显著富集于植物激素信号转导途径、17个基因富集于植物-病原菌互作途径、15个基因富集于苯丙酸生物合成途径、13个基因富集于氨基糖和核苷酸的糖代谢途径。差异表达基因MapMan可视化分析进一步表明,超量表达显著影响细胞壁、压力和信号传递功能组(图7-D)。特别地,超量表达显著上调生物胁迫功能组,而显著下调生长素信号转导Aux/IAA 家族功能组(图7-D)。

图4 CsGH3.6转基因植株的表型分析

箭头指示气孔arrows indicate stomata

图6 CsGH3.6转基因植株表皮细胞长度(A)和气孔密度(B)统计分析
2.6.1 生长素途径相关基因 进一步利用MapMan详细调查了转录组数据库中与生长素合成和降解、生长素运输和信号转导相关的差异表达基因(表1)。在生长素合成-降解中有4个基因差异表达,其中3个上调表达,1个下调表达;生长素运输的基因有3个,均下调表达;而在生长素信号通路中差异表达的基因有19个,其中有17个基因下调表达,注释的13个AUX/IAA家族基因均下调表达。这些结果表明超量表达明显抑制生长素的运输和信号转导相关基因表达。
2.6.2 生物胁迫相关基因 图8显示MapMan注释的转基因株系b19中与生物胁迫相关的差异基因情况。有60个基因展现差异表达水平,这些基因涉及到压力感知、活性氧爆发、信号转导、基因转录和防御基因表达(图8)。除4个防御基因下调表达外,所有与生物胁迫相关的差异基因均上调表达,特别是42个防御基因中有38个上调表达。基因功能注释表明,所有防御基因均为病程相关蛋白PR基因。这些结果暗示,超量表达激活了植株的防御反应。
2.6.3 植物生长发育相关基因 MapMan分析显示,与植物生长发育相关的差异基因富集于细胞分裂与细胞周期、细胞组织、发育功能组和细胞壁。上调表达基因数目(60个)明显多于下调表达基因数目(50个)。细胞壁在植物生长发育中起着重要作用,转录组数据显示15个细胞壁相关基因上调表达,26个基因下调表达(图9)。
表2展示了差异倍数≥2的细胞壁相关基因功能注释。参与细胞壁合成的两个重要基因纤维素合成酶基因和细胞壁蛋白基因差异表达水平在5倍以上。3个参与细胞壁修饰的木葡聚糖内糖基转移酶基因也展现出较高的表达水平。角质合成相关基因和蜡质合成相关基因表达水平上调,而与细胞壁疏松相关的延展蛋白基因下调表达。
3 讨论
生长素响应因子在调控植物的生长发育、激素动态平衡、生物和非生物胁迫相关抗性中起着重要作用。本研究中,超量表达转基因植株中自由IAA水平显著降低,植株表型出现过表达导致自由IAA缺失引起的典型表型变化[16],证明CsGH3.6具有酰基化IAA的生物学功能。另外,转基因植株JA含量也显著下降,暗示CsGH3.6也可能具有酰基化JA的功能[28]。溃疡病抗性评价表明,超量表达显著增强植株对溃疡病的抗性[24]。而且,本研究发现,超量表达不仅抑制活性IAA的积累,且促进病原菌侵染后活性IAA水平的显著下降。生长素IAA被认为是一种致病因子,具有促进柑橘溃疡病菌致病的作用[5,24]。这些结果暗示,通过酰基化侵染部位活性IAA来正向调控柑橘对溃疡病的抗性。

A:基因表达量聚类热图 Clustering heat map of gene expression quantity;B:差异基因表达分析火山图Volcano map of differentially expressed genes;C:KEGG富集图 KEGG enrichment;D:MapMan可视化分析,红色表示上调,蓝色表示下调 MapMan visualization, red and blue indicate up-regulated and down-regulated, respectively
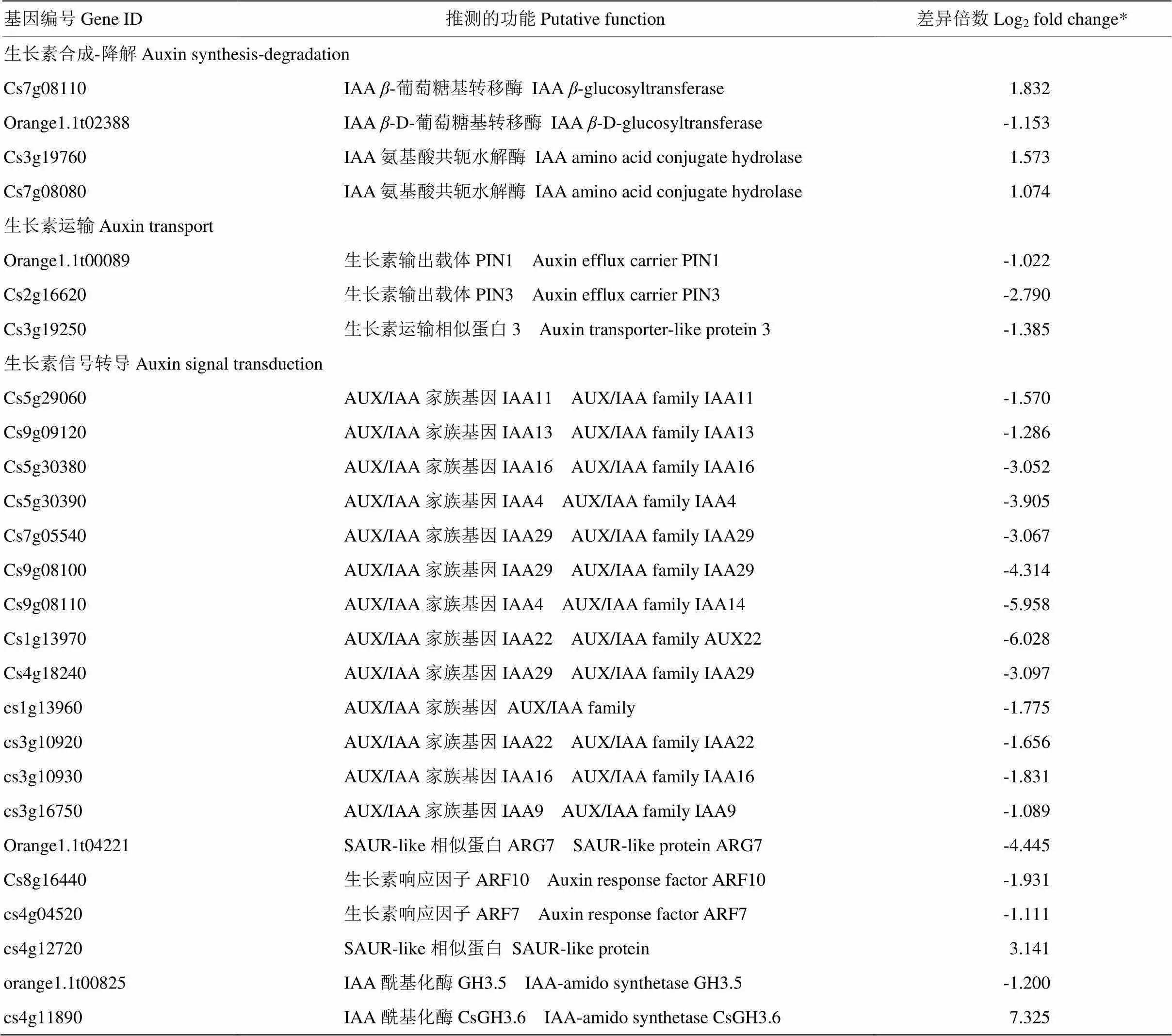
表1 b19转基因植株中生长素相关差异表达基因统计分析
*仅展示差异倍数|log2fold change|≥1的差异基因Differentially expressed genes with |log2fold change|≥1 are presented

图8 生物胁迫相关差异基因统计分析

图9 生长发育相关差异基因统计分析

表2 b19转基因植株中部分细胞壁相关的差异表达基因
病原菌侵染中脓疱形成是柑橘溃疡病症状发展的关键[29]。病原菌通过分泌IAA或促进寄主IAA的合成,调控寄主细胞的膨大和增生,进而促进脓疱的形成[5,24],以利于病原菌突出寄主表面向周围非侵染组织扩散。因此,植物表皮细胞在柑橘调控溃疡病抗性中起着重要作用[30]。本研究对转基因植株表皮细胞分析发现,表皮细胞变短。表皮细胞变短暗示活性IAA的下调抑制了细胞的正常生长,这种细胞生长的钝化可能不利于病原菌侵染中脓疱的形成和扩散。细胞壁是植物防御病原菌侵染的第一道物理屏障。转录组数据显示,细胞壁合成相关基因表达水平明显升高,特别是与增强细胞壁强度相关的角质和蜡质合成基因和表达水平上调;相反,与细胞壁疏松相关的延展蛋白基因表达水平下调。结果暗示转基因植株细胞壁强度增强。细胞壁的这种变化同样有利于抑制脓疱的形成,增强植株的抗性[29]。综上所述,超量表达抑制侵染部位活性IAA的积累,IAA含量的下降改变了细胞壁重构相关基因的表达或蛋白质的活性,进而抑制寄主细胞壁松弛和细胞膨大,阻碍病原菌的入侵和扩散,这可能是调控侵染部位脓疱形成的内在机制。另外,本研究发现转基因植株气孔密度显著增大。IAA负调控气孔发育[31],抑制IAA的积累和极性运输会诱发气孔簇生,比如拟南芥和突变体展现气孔簇生状[32-33]。转录组测序表明,转基因植株中和表达水平显著下降,与转基因植株气孔密度增大相关。气孔是溃疡病菌入侵柑橘的自然入口,有研究表明气孔密度与柑橘品种溃疡病抗性呈一定的负相关性,即气孔密度越大,品种的抗病性越低[34-35]。因此,气孔密度的增大可能会削弱超量表达对脓疱形成的抑制作用,影响转基因植株抗性水平的进一步提升。是如何从细胞发育水平影响病原菌侵染中脓疱的形成有待进一步深入研究。
本研究显示,超量表达增强了柑橘的抗病反应。首先,激素分析表明,转基因植株SA含量显著增加,暗示SA介导的抗性反应增强。在温室和田间中,外施SA及其类似物能有效增强柑橘对溃疡病的抗性,已广泛用于柑橘溃疡病防治中[36]。同样,超量表达SA信号下游的中心调节子基因显著增强转基因柑橘对溃疡病的抗性,这种抗性增强与PR基因的表达增强紧密相关[37]。因此,转基因植株中大量PR基因的上调表达暗示超量表达增强了SA介导的抗病反应。其次,JA通常拮抗SA介导的抗病反应[38],因此,超量表达转基因植株JA含量降低有利于增强植物抗病性。有研究表明,侵染部位高浓度的IAA能抑制SA介导的抗病反应[7,10-11],说明转基因植株中IAA积累的下降能促进SA介导的抗病反应。综上,超量表达可能通过削弱或解除IAA和JA对SA抗病途径的抑制,进而增强植株的防御反应。
超量表达对植株的形态发育造成了严重影响。转基因植株呈现叶片向上卷曲,分枝增多,植株萎蔫,丧失顶端优势的表型,与拟南芥超量表达及其获得性突变体植株表型相似[39]。生长素是顶端优势形成的重要调控物质。当生长素运输受阻或其信号通路受抑制,生长素对侧芽活动的抑制解除,进而导致植株顶端优势丧失,分枝增多,叶小而卷曲[40]。转录组测序显示,转基因植株中减少的自由IAA显著抑制生长素的运输和信号转导途径相关基因表达,特别是所有注释的AUX/IAA家族基因均显著下调表达。AUX/IAA是生长素早期响应因子,通过泛素化途径调控转录因子的表达,进而影响细胞大小和植株生长发育[41]。当细胞中IAA降低,AUX/IAA抑制ARF激活下游基因、、的表达,最终抑制IAA的信号转导,影响植株的正常生长[40]。这种抑制作用会进一步抑制转录[41]。而和的表达下调说明生长素在植物体内的分配受阻。另外,超量表达导致的自由IAA减少进一步上调IAA糖基化和酰基化基因表达,这会进一步抑制自由IAA的积累。这些结果清楚说明,超量表达显著抑制了IAA的积累和信号转导,进而影响许多与细胞、组织和个体发育相关基因的表达,改变植株的形态特征。
由于超量表达转基因植株生长发育受到影响,尽管已经获得了溃疡病抗性提高的转基因株系,但在柑橘溃疡病抗性育种方面依然受到限制。一种解决策略是利用病原菌诱导型启动子驱动的表达,当植物感染病原病菌时才会启动表达,从而有利于获得表型正常且具有溃疡病抗性的转基因柑橘[42]。
4 结论
超量表达通过酰基化自由IAA来抑制生长素信号转导,改变植物细胞的形态建成以利于寄主抗性的提升,同时,解除IAA和JA对SA的拮抗,促进SA介导的抗病反应,增强柑橘对溃疡病的抗性,研究结果为利用生长素信号转导改良柑橘溃疡病抗性提供了依据。
[1] 何秀玲, 袁红旭. 柑橘溃疡病发生与抗性研究进展中国农学通报, 2007, 23(8): 409-412.
HE X L, YUAN H X. Research advances on the occurrence and resistance of citrus bacterial canker disease., 2007, 23(8): 409-412. (in Chinese)
[2] Gong X Q, Liu J H. Genetic transformation and genes for resistance to abiotic and biotic stresses inand its related genera, 2013, 113(2): 137-147.
[3] Costacurta A, Mazzafera P, Rosato Y B. Indole-3-acetic acid biosynthesis bypv.is increased in the presence of plant leaf extracts, 1998, 159(2): 215-220.
[4] Cernadas R A, Camillo L R, Benedetti C E. Transcriptional analysis of the sweet orange interaction with the citrus canker pathogenspv.andpv., 2008, 9(5): 609-631.
[5] Cernadas R A, Benedetti C E. Role of auxin and gibberellin in citrus canker development and in the transcriptional control of cell-wall remodeling genes modulated bypv., 2009, 177(3): 190-195.
[6] Chen M, HE Y r, XU L z, Peng A H, Lei T G, Yao L X, LI Q, Zhou P F, Bai X J, Duan M J, JIANG X Y, JIA R R, ZOU X P, CHEN S C. Cloning and expression analysis ofgenesandresponding topv.infection, 2016, 2(4): 193-202.
[7] Duca D, Lorv J, Patten C L, Rose D, Glick B R. Indole-3- acetic acid in plant-microbe interactions, 2014, 106(1): 85-125.
[8] 傅晶. 抑制病原菌诱导的生长素的积累赋予水稻广谱抗性[D]. 武汉: 华中农业大学, 2010.
FU J. Manipulating broad-spectrum disease resistance by suppressing pathogen-induced auxin accumulation in rice[D]. Wuhan: Huazhong Agricultural University, 2010. (in Chinese)
[9] Fu J, Liu H B, Li Y, Yu H H, Li X H, Xiao J H, Wang S P. Manipulating broad-spectrum disease resistance by suppressing pathogen-induced auxin accumulation in rice, 2011, 155(1): 589-602.
[10] Mutka A M, Fawley S, Tsao T, Kunkel B N. Auxin promotes susceptibility tovia a mechanism independent of suppression of salicylic acid-mediated defenses, 2013, 74(5): 746-754.
[11] Robert-seilaniantz A, Grant M, Jones J D G. Hormone crosstalk in plant disease and defense: More than just jasmonate- salicylate antagonism, 2011, 49: 317-343.
[12] Chen Y, Shen H, Wang M, Li Q, He Z. Salicyloyl-aspartate synthesized by the acetyl-amido synthetase GH3.5 is a potential activator of plant immunity in, 2013, 45(10): 827-836.
[13] Westfall C S, Herrmann J, Chen Q, Wang S, Jez J M. Modulating plant hormones by enzyme action: the GH3 family of acyl acid amido synthetases, 2010, 5(12): 1607-1612.
[14] Hagen G, Guilfoyle T J. Rapid induction of selective transcription by auxins, 1985, 5(6): 1197-1203.
[15] Du H, Wu N, Fu J, Wang S P, Li X H, Xiao J H, Xiong L Z. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice, 2012, 63(18): 6467-6480.
[16] Domingo C, Andres F, Tharreau D, Iglesias D J, Talon M. Constitutive expression ofreduces auxin content and enhances defense response and resistance to a fungal pathogen in rice, 2009, 22(2): 201-210.
[17] Ding X, Cao Y, Huang L, Zhao J, Xu C, Li X, Wang S. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice, 2008, 20(1): 228-240.
[18] Singh V K, Jain M, Garg R. Genome-wide analysis and expression profiling suggest diverse roles ofgenes during development and abiotic stress responses in legumes, 2015, 5: Article 789.
[19] Yuan H, Zhao K, Lei H, Shen X, Liu Y, Liao X, Li T. Genome-wide analysis of the GH3 family in apple (×), 2013, 14: 297.
[20] Feng S, Yue R, Tao S, Yang Y, Zhang L, Xu M, Wang H, Shen C. Genome-wide identification, expression analysis of auxin- responsive GH3 family genes in maize (L.) under abiotic stresses, 2015, 57(9): 783-795.
[21] Kumar R, Agarwal P, Tyagi A K, Sharma A K. Genome- wide investigation and expression analysis suggest diverse roles of auxin-responsivegenes during development and response to different stimuli in tomato (), 2012, 287(3): 221-235.
[22] Yu D, Qanmber G, Lu L, Wang L, Li J, Yang Z, Liu Z, Li Y, Chen Q, Mendu V, Li F, Yang Z. Genome-wide analysis of cotton GH3 subfamily II reveals functional divergence in fiber development, hormone response and plant architecture, 2018, 18: 350.
[23] Yang Y, Yue R, Sun T, Zhang L, Chen W, Zeng H, Wang H, Shen C. Genome-wide identification, expression analysis of GH3 family genes inunder stress-related hormones andinfection, 2015, 99(2): 841-854.
[24] 陈敏. 超量表达生长素早期响应基因增强柑橘溃疡病抗性[D]. 重庆: 西南大学, 2017.
Chen M. Overexpressing early auxin-responsive geneenhances canker resistance in citrus[D]. Chongqing: Southwest University, 2017. (in Chinese)
[25] Marques J P R, Amorim L, Silva-Junior G J, Sposito M B, Appezzato-da Glória B. Structural and biochemical characteristics of citrus flowers associated with defence against a fungal pathogen, 2014, 7: plu090.
[26] Peng A H, Chen S C, Lei T G, Xu L Z, He Y R, Wu L, Yao L X, Zou X P. Engineering canker-resistant plants through CRISPR/Cas9- targeted editing of the susceptibility genepromoter in citrus, 2017, 15: 1509-1519.
[27] Thimm O, Blasing O, Gibon Y, Nagel A, Meyer S, Kruger P, Selbig J, Muller L A, Rhee S Y, Stitt M. MAPMAN: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes, 2004, 37(6): 914-939.
[28] Gutierrez L, Mongelard G, Floková K, Păcurar D I, Novák O, Staswick P, Kowalczyk M, Păcurar M, Demailly H, Geiss G, Bellini C. Auxin controlsadventitious root initiation by regulating jasmonic acid homeostasis, 2012, 24(6): 2515-2527.
[29] Hu Y, Zhang J L, Jia H G, Sosso D, Li T, Frommer W B, Yang B, White F F, Wang N, Jones J B.is a disease susceptibility gene for citrus bacterial canker disease, 2014, 111(4): E521-E529.
[30] Brunings A M, Gabriel D W.: breaking the surface, 2003, 4(3): 141-157.
[31] 陈亮, 侯岁稳. 植物气孔发育的分子遗传调控中国科学: 生命科学, 2017, 47(8): 798-807.
Chen L, HOU S W. Molecular genetic control of plant stomatal development., 2017, 47(8): 798-807. (in Chinese)
[32] 周丽娟, 陈尔娟, 韩笑, 何用娟, 陈善娜, 陈小兰. 激素与气孔发育研究进展西北植物学报, 2015, 35(4): 845-851.
ZHOU L J, CHEN E J, HAN X, HE Y J, CHEN S N, CHEN X L. Review on hormone regulation of stomatal development., 2015, 35(4): 845-851. (in Chinese)
[33] Le J, Liu X G, Yang K Z, Chen X L, Zou J J, Wang H Z, Wang M, Vanneste S, Morita M, Tasaka M, DING Z J, Friml J, Beeckman T, Sack F. Auxin transport and activity regulate stomatal patterning and development, 2014, 5: 3090.
[34] 李敏, 段硕, 李中安, 周彦, 周常勇, 谭锦, 彭耀武. 叶片微形态结构特征与柑桔溃疡病抗性的关系中国南方果树, 2013, 42(2): 1-5.
LI M, DUAN S, LI Z A, ZHOU Y, ZHOU C Y, TAN J, PENG Y W. Analysis of relationship between citrus canker resistance and leaf micro-morphological characteristics., 2013, 42(2): 1-5. (in Chinese)
[35] 温寿星, 黄镜浩, 陈瑾, 蔡子坚, 包榕, 张凌媛. 叶片结构与柑橘溃疡病抗性的初步研究中国农学通报, 2009, 25(13): 66-69.
WEN S X, HUANG J H, CHEN J, CAI Z J, BAO R, ZHANG L Y. Preliminary studies on leaves structure in resistant and susceptible cultivars of citrus., 2009, 25(13): 66-69. (in Chinese)
[36] Francis M I, Redondo A, Burns J K, Graham J H. Soil application of imidacloprid and related SAR-inducing compounds produces effective and persistent control of citrus canker, 2009, 124(2): 283-292.
[37] Zhang X, Francis M I, Dawson W O, Graham J H, Orbović V, Triplett E W, Mou Z. Over-expression of thegene in citrus increases resistance to citrus canker, 2010, 128(1): 91-100.
[38] Li J, Brader G, Palva E T. The WRKY70 transcription factor: a node of convergence for jasmonate-mediated and salicylate-mediated signals in plant defense, 2004, 16(2): 319-331.
[39] Zhang Z, Li Q, Li Z, Staswick P E, Wang M, Zhu Y, He Z. Dual regulation role ofduring Arabidopsis-interaction, 2007, 145(2): 450-464.
[40] Zhao Y. Auxin biosynthesis and its role in plant development, 2010, 61: 49-64.
[41] Tiwari S B, Wang X J, Hagen G, Guilfoyle T J. AUX/IAA proteins are active repressors, and their stability and activity are modulated by auxin, 2001, 13(12): 2809-2822.
[42] 宋二玲. 三个病原物诱导启动子在转基因柑橘中受溃疡病菌和创伤诱导的表达分析[D]. 重庆: 西南大学, 2013.
SONG E L. Canker bacterium- and wound-response characteristics of three pathogen-induced promoters in transgenic citrus[D]. Chongqing: Southwest University, 2013. (in Chinese)
Overexpression ofenhanced resistance to citrus canker disease by inhibiting auxin signaling transduction
Zou XiuPing, Long JunHong, Peng AiHong, Chen Min, Long Qin, Chen ShanChun
(National Center for Citrus Variety Improvement, Citrus Research Institute, Southwest University/Chinese Academy of Agricultural Sciences, Chongqing 400712)
【】Citrus canker, induced bysubsp(), is one of the most destructive disease in citrus production. Auxin plays an important role in regulating-induced pustule formation in citrus., an early auxin response gene, regulates plant hormone homeostasis through acylating indole-3-acetic acid (IAA). The previous study found thathad a vital role in response toinfection. 【】Here, to explore the internal mechanism of) overexpressingwere performed in this study.【】To evaluate resistance to citrus canker in transgenic plants overexpressing,fully expanded intact leaves gathered from transgenic plants were infected withby the pin-prick inoculation, and the diseased areas and disease severity index were investigated 10 d after inoculation, the wild type (WT) plant was used as the control. To detect hormone levels in transgenic plants, different hormones were isolated from leaves before and afterinfection and their contents were determined through high performance liquid chromatography (HPLC). Compared with WT plant, changes of phenotypes (plant types and leaf longitudinal diameter, transverse diameter and thickness) in transgenic plants were investigated in the greenhouse, and the length of epidermal cell and stomata density were further analyzed using optical microscopy. RNA sequencing was performed to investigate transcript changes in transgenic plant, and gene functions were annotated based on Nr, Nt, Pfam, COG, SwissProt and gene ontology (GO) databases. To elucidate the molecular mechanism ofwere investigated using the KEGG database and MapMan software.【】Overexpression ofsignificantly enhanced citrus canker resistance in transgenic plants. The branches of transgenic plants increased and drooped, the leaves curled upward, became smaller and lighter in color. The stomata density of transgenic plants increased and the length of epidermal cells became shorter. Hormone analyses showed that the contents of free auxin IAA and jasmonic acid (JA) in transgenic plants decreased significantly, while the content of salicylic acid (SA) increased significantly. Transcriptome sequencing showed that overexpression ofsignificantly inhibited the expression of auxin signal transduction related genes, especially the expression level of all the predicted Aux/IAA genes was down-regulated in transgenic plant. Conversely, the expression level of genes related to biological stress was up-regulated, most of which were pathogenesis-related genes. 【】Overexpression ofcan inhibit auxin signal transduction through acylating free IAA, regulate the homeostasis of JA and SA, change the morphogenesis of cells and plants, and finally enhance the plant resistance to citrus canker. The results suggest that the regulation of hormone homeostasis has potential value in citrus disease resistance breeding.
subsp(); citrus canker; auxin;; resistance
10.3864/j.issn.0578-1752.2019.21.009
2019-05-23;
2019-07-15
国家重点研发计划(2018YFD1000300)、国家现代农业产业技术体系建设专项资金(CARS-27)、重庆市自然科学基金(cstc2017jcyjBX0020)、广东省科技创新战略专项资金(2018B020202009)
邹修平(通信作者),E-mail:zouxiuping@cric.cn
(责任编辑 岳梅)
猜你喜欢
中国医院用药评价与分析(2022年3期)2022-04-07 09:10:12
基层中医药(2021年2期)2021-07-23 01:41:48
生物学通报(2021年4期)2021-03-16 05:41:34
科学大众(2020年12期)2020-08-13 03:22:38
乡村科技(2019年9期)2019-06-26 03:52:04
今日农业(2019年14期)2019-01-04 08:57:40
数码世界(2018年7期)2018-12-24 15:06:05
现代园艺(2018年3期)2018-02-10 05:18:39
光明中医(2016年8期)2016-08-15 03:17:43
体育世界(学术版)(2016年7期)2016-08-15 00:42:52
