
原料比例与接种量对猪粪秸秆厌氧干发酵产气率及微生物群落的影响
2019-11-16 06:37:54刘丽丽张克强杜连柱齐利格娃丁文涛高文萱
中国沼气 2019年6期
李 奥,刘丽丽,张克强,杜连柱,齐利格娃,丁文涛,高文萱
(1.农业部环境保护科研监测所,天津 300191;2. 天津师范大学生命科学学院,天津 300387;3.北京化工大学,北京 100029)
能源与环境是当今人类面临的两大问题。将农业废弃物用于沼气发酵对于改善农村环境、缓解能源消耗具有重要意义。目前的沼气工程主要以畜禽粪便为原料,而畜禽粪便的大量消耗以及养殖数量的波动使得畜禽粪便供应不足的问题日益突出[1]。将农作物秸秆与畜禽粪便混合进行厌氧发酵制沼气,不仅能改善农村环境实现秸秆资源化利用,更能够减少畜禽粪便的消耗,提高沼气工程运行的持续性和稳定性[2]。此外混合秸秆发酵还可以改善单一原料进行厌氧发酵消化效率不高的问题,提高厌氧发酵的消化效果[3-5]。由此可见将农作物秸秆与畜禽粪便混合厌氧发酵制沼气,是解决当前沼气生产原料问题的首要途径[6]。而干发酵的含水率低,处理后的残留物可做堆肥使用,不会造成水体污染;发酵过程中容积产气量高,因此采用厌氧干发酵的方式制沼气符合环保、节约型社会发展的要求。
为了获得较佳的产气性能,刘战广[7]等研究表明,调节粪草比并不能提高原料的产气潜能,但在营养调节和结构改良方面有一定促进作用。同时有研究表明接种物的来源、富集培养方式及添加比例对厌氧发酵影响很大,一般认为接种物添加量为发酵料液的 10 %~15%,既可实现正常启动其运行也比较稳定[8]。对于干发酵来讲,由于含固量(Total solid,TS)比较高(一般在 17%以上)[9],若接种量不足,产甲烷菌数量相对较少,容易造成VFA积累,出现“酸中毒”,需要重新调节,给生产带来不便。孙国朝[10]等研究发现,加大接种量,是防止前期酸化、缩短干发酵启动时间的关键措施[6]。马传杰[11]等人研究发现若接种量较大,可以实现快速启动,但会占据较多池容积,造成消化池容积利用率低。房明[12]等研究了接种比对餐厨垃圾中温厌氧消化的影响,结果表明,随着接种比逐渐提高(1∶1~4∶1),餐厨垃圾的发酵延滞期逐渐缩短。李文哲[13]等在不同接种量对稻秆厌氧发酵特性研究中发现,适量的接种物可以提高消化系统的缓冲能力,有利于产气高峰提前。由此可见,明确不同接种量对厌氧发酵的影响,进而实现厌氧发酵条件的科学控制,对于促进厌氧发酵工程实践具有重要意义。本试验选取常见的农业废弃物猪粪和水稻秸秆为底物,研究猪粪与秸秆在不同VS质量比条件下混合产沼气的特性,探索不同接种量对猪粪秸秆厌氧干发酵的影响,为厌氧干发酵的科学运行与管理奠定理论基础。
1 材料和方法
1.1 底物与接种物
猪粪和秸秆均取自天津市西青区益利来养殖有限公司,猪粪为养殖场日产鲜猪粪,取回后储存于4℃±1℃的冰箱,秸秆风干后粉碎至0.5~1.0 cm,并存放于干燥阴凉处。接种物取自实验室正常运行的中温混合厌氧反应器(continuous stirred tank reactor,CSTR)。活性污泥取出后4000 r·min-1离心 20 min,去上清液后留沉淀物(接种物)备用。并将接种物在室温下活化微生物3天后使用。上清液用于调节发酵体系的总固体含量(TS)到20%。底物与接种物的理化指标见表1。

表1 底物和接种物的化学组分
1.2 试验装置
采用气袋法进行厌氧批式试验,具体方法如下:采用有效容积为500 mL的厌氧瓶作为厌氧发酵装置(见图1)。瓶口顶部为丁基橡胶塞,并在橡胶塞中部打孔以连接气袋搜集气体,发酵过程中产生的沼气收集于3 L铝制集气袋中。

图1 发酵装置结构图
1.3 试验设计
试验方案设计如表2所示。猪粪与秸秆的比例(VS质量比)选择1∶1和2∶1;接种量为接种物占底物(猪粪+秸秆)的VS质量比(30%,40%,50%)。物料TS浓度为20%。每种发酵方式有3组平行,每个发酵瓶中有机负荷为110 gVS·L-1,装有物料总质量为250 g。发酵原料(见表2)添加完毕后,向厌氧发酵瓶中冲入氮气,持续2 min,以排尽厌氧瓶顶部空间中的空气,保持发酵体系的厌氧环境。然后在厌氧瓶顶部塞上丁基橡胶塞,加盖拧紧并在塞口用橡皮管连接气袋。将各厌氧瓶于37℃±1℃的恒温培养箱中避光培养,每天下午2点取下气袋,用气筒抽取测量产气体积,并定期用2 mL注射器采集气体样品,用于CH4和CO2含量测定。

表2 各处理组原料、接种物加入量
1.4 分析方法
沼气产量使用气袋进行搜集,并用气筒抽取进行测定;总固体含量(TS)、挥发性固体含量(VS)采用标准方法测定[14];将所取发酵样品用蒸馏水稀释10倍测量pH值;甲烷百分含量通过气相色谱仪测定(Trace1300,Thermo,美国),色谱柱采用2 m×φ3 mm的Porapak Q 柱,检测器为热导检测器(TCD),采用高纯氦气作为载气,测定条件为:载气流速8 mL·min-1,柱温40℃,检测器温度200℃,进样口温度120℃;样品稀释后用稀硫酸调节pH值<3.0,离心(10000 rpm,25℃条件下离心10 min),过滤(0.45 μm有机滤膜)滤液经丙酮稀释后采用气相色谱仪(Thermotrace1300)测定挥发性脂肪酸(VFAs),M 12毛细管柱(30 m×0.53 mm×1 μm);进样口温度为200℃,检测器温度为220℃,载气流速8 mL·min-1。根据胡荣笃[15]的计算方法,将发酵液中各种脂肪酸的浓度换算成乙酸的浓度来计算分析。
DNA采用Fast DNAs Spin Kit (Mpbio,美国)试剂盒提取,发酵前后各处理的3个重复分别提取DNA,通过超微量分光光度计(Nano Drop 2000,Thermo Scientific,Wilmington,美国)测定浓度,然后分别将各处理3个重复提取的DNA混匀,样品送至生工生物工程(上海)股份有限公司进行微生物分类测序,测序类群为细菌和古菌,测序平台为MiseqTM,测序长度为2×300bp,扩增区域均为 V3-V4,采用巢式PCR;细菌测序引物为341F,(序列F:CCTACGGGNGGCWGCAG);805R,(序列R:GACTACHVGGGTATCTAATCC);古菌测序引物为340F,(序列F:CCCTAYGGGGYGCASCAG);1000R,(序列R:GGCCATGCACYWCYTCTC)。测序后的 DNA 序列进行拼接,通过 barcode 标签序列区分样品序列,采用 Prinseq(0.20.4)对样本序列做质量控制,在 QIIME 中调用Uclust(1.1.579)软件,设置 97%相似性,对有效 DNA序列数据进行操作分类单元(OTU)分类,采用 RDP软件比对 Silva 数据库进行物种分类,在门、纲、目、科和属分类水平上统计样本的物种丰度。采用 Mothur软件计算种群丰富度指数(Chao 指数、ACE 指数)和群落多样性指数(Shannon 指数和 Simpson 指数)。
2 结果与分析
2.1 不同原料比例与接种量下干发酵产气特性
各处理组日产气量和甲烷体积分数如图2和图3所示,各试验组在发酵前期和后期各出现一个产气高峰。猪粪∶秸秆为1∶1,接种量为50%的试验组(PH-1)在第7天最先达到产气高峰,产量为233.33 mL,甲烷体积分数为32.66%,随后迅速下降;第2个产气高峰在第31天出现,产量为228.33 mL,甲烷体积分数为62.46%。PH-1发酵启动速度明显要快于其它试验组,表明在此比例下厌氧发酵过程中加大接种量有利于加快产气高峰出现。接种量为30%的试验组(PL-1)在第17天达到第1个产气高峰,40%接种量的试验组(PM-1)在第16天达到第1个产气高峰,产气量分别为231.67 mL和176.67 mL,甲烷体积分数分别为51.88%和39%。猪粪∶秸秆为2∶1,接种量为50%的试验组(PH-2)在第14天达到第一个产气高峰,产气量213.33 mL,甲烷体积分数为55%;接种量为30%(PL-2)与40%(PM-2)的试验组分别在第18天和第12天达到产气高峰,产气量分别191.66 mL和211.67 mL,甲烷体积分数分别为57%和41%;在此比例下产气高峰时间与产气量相差不大,说明接种量对其产气性能影响不大。

图2 各处理组沼气日产量变化曲线
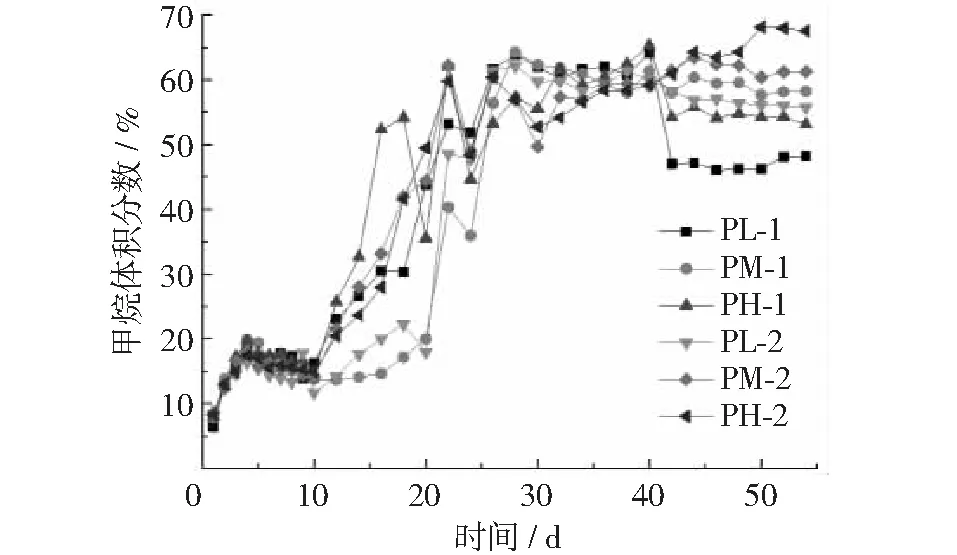
图3 各处理组甲烷体积分数变化曲线
累积VS甲烷产率如图4所示。猪粪∶秸秆为1∶1和2∶1时累积产气量趋势大致相同,产气速率均为前期缓慢,后期加快,最终处于平缓状态,累积甲烷产气率均为接种量50%的试验组最高,累积沼气产量分别为6801.67 mL和6038.33 mL,累积VS甲烷产率分别达到127.07 mL·g-1VS,116.91 mL·g-1VS。同种接种量、不同原料比例下累积VS甲烷产率均为1∶1高于2∶1。

图4 各处理组累积VS甲烷产率的变化曲线
2.2 不同原料比例及接种量对TS、VS降解率的影响
发酵原料的TS,VS降解率是衡量厌氧消化性能的重要指标[9]。不同试验组TS,VS降解率(见图5)采用发酵前后总TS,VS量计算。不同试验组VS降解率为29% ~46%,TS降解率为12%~29%。猪粪∶秸秆为2∶1接种量为40%(PM-2)降解率较高,TS降解率为46%,VS降解率为29%,显著高于其他处理组,其余处理组并无显著性差异(p>0.05)。

图5 不同配比及接种量TS,VS下发酵降解率
2.3 微生物群落多样性分析
2.3.1 Alpha多样性分析
提取发酵前后样品总DNA,采用宏基因组测序的方法分析细菌、古菌群落结构。在相似度97%的条件下利用mothur做rarefaction 分析得知发酵前后各处理细菌和古菌Shannon指数稀释曲线随着序列数增加而快速趋于平坦,说明测序数据合理,可以反应样品的Alpha多样性[16]。
对样本进行细菌Alpha多样性分析,结果如表3所示。Coverage指数值反映测试结果的真实情况,各样本Coverage值均大于0.99,说明能很好的反映测试结果。比较各组处理后多样性指数,发现Shannon,Simpson,ACE和Chao1指数均相差不大,说明各处理发酵后细菌群落多样性基本一致,原料配比和接种量对发酵后细菌多样性无显著影响。

表3 发酵过程中细菌的丰富度和多样性变化
发酵前后各处理组古菌丰富度及多样性变化见表4。各处理古菌OTU数量显著低于细菌,且发酵前后OTU数量变化趋势随接种量的不同而差异。30%和40%接种量处理(PL-1,PM-1,PL-2和PM-2)使得发酵后OTU数量和Chao1指数增加,而50%接种量处理(PH-1和PH-2)使得发酵后OTU数量和Chao1指数降低,说明30%和40%接种量处理增加了古菌丰富度,而50%接种量降低了古菌丰富度。究其原因,各处理发酵后OTU数量相差不大,而50%接种量发酵前OTU数量高于30%和40%接种量,这些增加的OTU随着干发酵的进行而被竞争消失,所以呈现不同的变化规律。发酵后接种量为30%的试验组(PL-1和PL-2)Shannon指数明显低于接种量为40%(PM-1和PM-2)和50%(PH-1和PH-2)的试验组,且接种量为40%的试验组(PM-1和PM-2)Shannon指数最高,说明在一定范围内随着接种量的增加古菌群落多样性也随之增加;而相同接种量不同原料比下Shannon指数相差不大,说明不同原料比对古菌群落多样性的影响不大。接种量为30%的试验组(PL-1和PL-2)Simpson指数最大,而同种接种量不同原料比下各处理Simpson指数相近,表明在一定范围内随接种量的增加古菌优势菌群的生物量占总生物量的比例在增加;而不同原料比例对优势古菌基本没有影响。

表4 发酵过程中古菌的丰富度和多样性变化
2.3.2 菌群结构分析

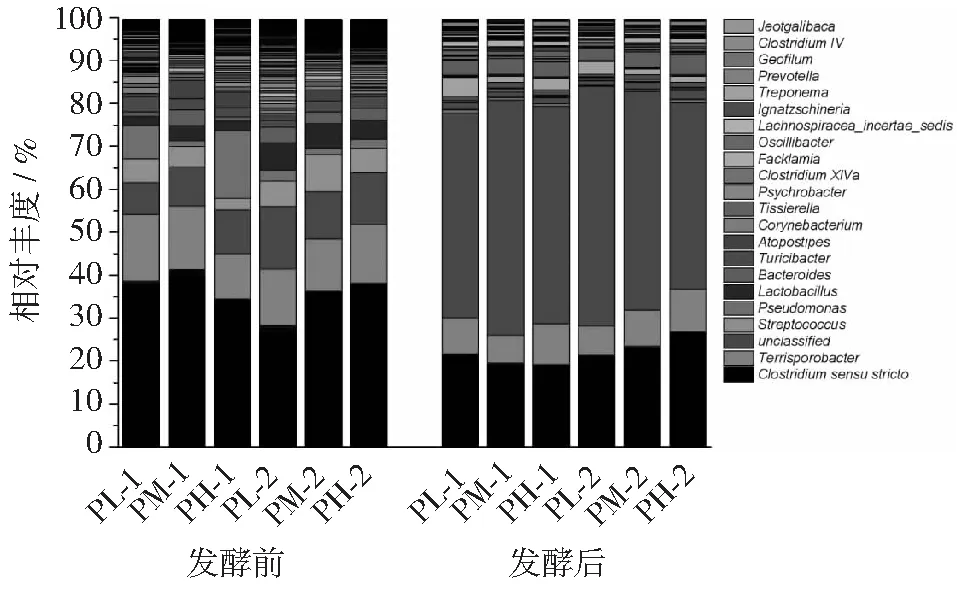
图6 细菌群落结构变化(属水平)
本研究发酵前后古菌群落结构变化见图7。Methanobrevibacter、Methanoculleus和Methanomassiliicoccus为优势古菌,丰度分别为15.1%~45.84%,39.35%~61.97和13.99%~42.21%,且Methanoculleus为发酵后新增优势菌群。另一种优势菌群Methanosarcina是已知的唯一能够利用所有产甲烷途径的菌属,可利用甲基类化合物生长,有的也利用乙酸,H2/CO2,CO甚至丙酮酸盐[20]。其抗逆性较强,可高效利用有机酸转化为甲烷[21],其发酵后的丰度明显升高,发酵前菌群丰度为0.07%~0.65%,而发酵后菌群丰度为2.91%~14.30。Methanoculleus和Methanosphaerula均是氢营养型产甲烷菌[22],Methanosphaerula发酵前丰度在6.94%~24.06%,发酵后丰度降为3.44%~9.3%。接种量与古菌群落结构存在相关性,在两种原料配比下Methanomassiliicoccus均在接种量为40%的处理组中丰度最高,而Methanosphaerula均在接种量为50%的处理组中丰度最高。原料配比的改变未观察到对古菌群落结构产生明显影响,说明古菌群落结构对接种量较敏感而对原料配比不敏感。

图7 古菌群落结构变化(属水平)
发酵前后细菌(属水平)主成分分析如图8和图9所示。发酵前环境因子TS、pH值与优势菌属Pseudomonas,Clostridiumsensustricto呈正相关;菌群Terrisporobacter,Streptococcus以及Lactobacillus的生长与VFA有密切的相关性,在发酵后受VFA降低的影响,菌群Terrisporobacter,Streptococcus以及Lactobacillus的丰度也随之降低。且随着发酵的进行发现嗜蛋白质菌属Proteiniphilum与密螺旋体Treponema菌群的丰度大量增加。在主分类轴PCA1(解释75.8%差异)上,两种比例各试验组分布均较远,且在猪粪∶秸秆为2∶1比例下随着接种量的提高,各样品在PCA1上从左至右依次分布,在PCA2(解释8.9%差异)上从下至上依次分布,说明在此比例下接种量成为影响细菌群落结构的主要因素。
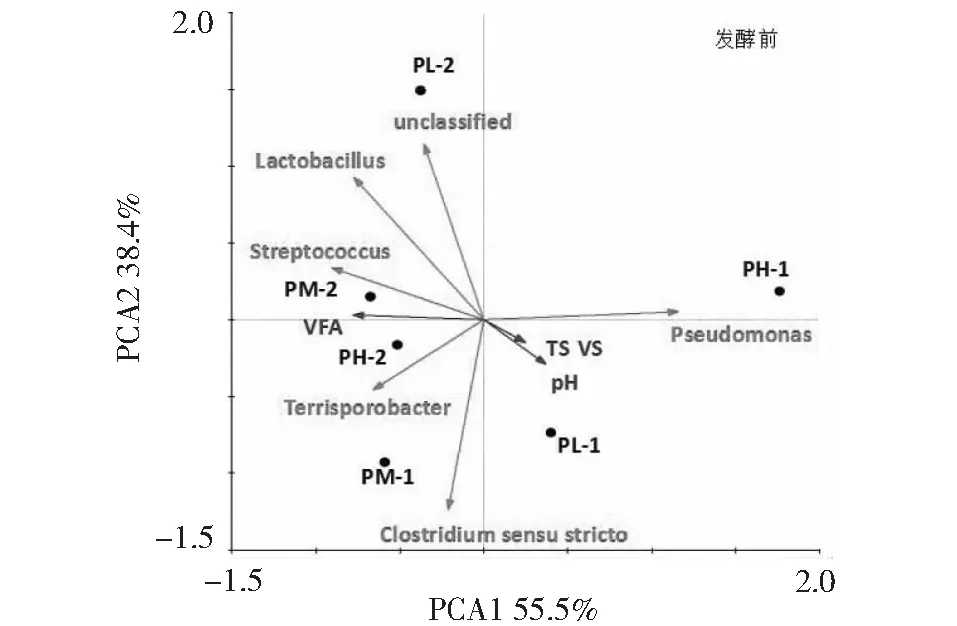
图8 发酵前细菌群落结构主成分分析

图9 发酵后细菌群落结构主成分分析
发酵前后古菌(属水平)群落主成分分析如图10和图11所示。发酵前环境因子TS、VFAs与优势菌属Methanomassiliicoccus呈正相关,与Methanobrevibacter、Methanosphaerula呈负相关。环境因子pH对优势菌群Methanobrevibacter和Methanosphaerula有密切影响,TS对菌群Methanomassiliicoccus影响较大。发酵前猪粪∶秸秆为1∶1的试验组随接种量的升高,各样品在主分类轴PCA1(解释92.6%上的差异)上从右至左依次分布,说明接种量成为影响古菌群落结构的主要因素。而发酵前PL-2和PM-2在PCA1轴上分布较近,发酵后在PCA1轴上分布较远,说明随着发酵的进行接种量对古菌群落结构影响逐渐增加。
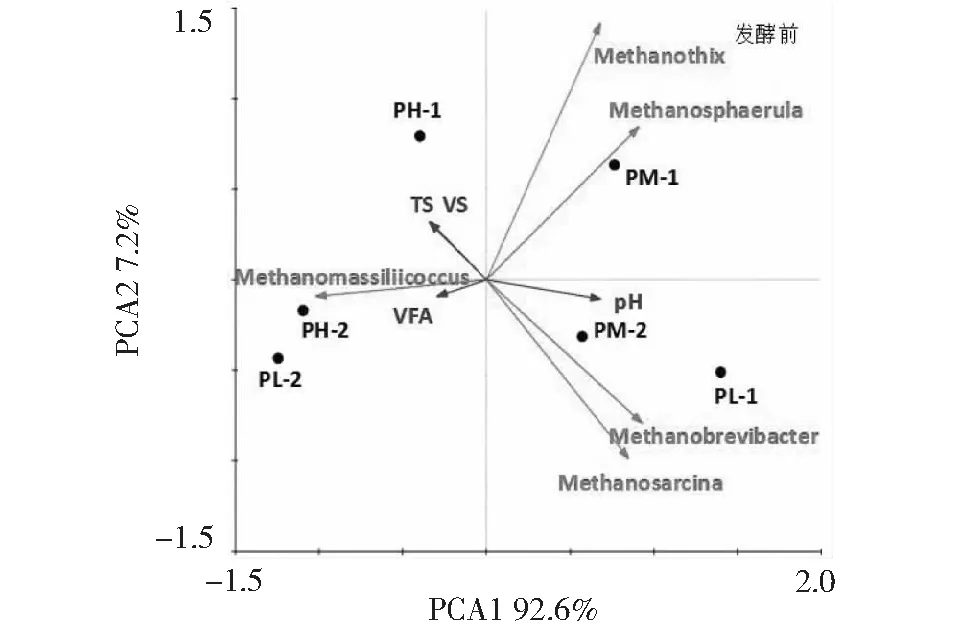
图10 发酵前古菌群落结构主成分分析

图11 发酵后古菌群落结构主成分分析
3 讨论
接种物的质量对于厌氧消化中产甲烷阶段的运行效果和稳定性很重要,如果接种量偏少,产甲烷数量相对较少,容易造成“酸中毒”;若接种量较大虽可实现快速启动,在保证高处理效率的条件下,反应器的容积必然增大。而刘战广[7]等的研究表明,调节粪草比例虽不能提高原料的产气潜能,但在营养调节和结构改良方面有一定促进作用。因此,选择合适的原料比例和接种量对厌氧干发酵的运行有重要的意义。
本文研究结果显示,猪粪∶秸秆为1∶1和2∶1累积甲烷产气量均为接种量50%的试验组最高,说明接种物的增加有利于提高产气量;而相同接种量下甲烷累积产量均为1∶1高于2∶1。可能因为2∶1试验组中猪粪含量较高,而猪粪主要由蛋白质、糖类和脂肪等易降解的组分组成[23],在发酵过程中容易发生挥发性脂肪酸的积累,抑制产甲烷菌的活性。猪粪∶秸秆为1∶1试验组随接种量增加,累积甲烷产量也随之增加,这一结果与李文哲[13]等对不同接种量对稻秆厌氧发酵特性的影响的试验结果一致;而猪粪∶秸秆为2∶1试验组累积甲烷产气量随接种量的增加呈现先降低后增加的趋势,这与李铁[24]等研究结果一致,说明菌种对于沼气产量的增加作用在一定比例下不明显,当菌种量增加到一定程度后才对沼气产量有明显的促进作用。总体来看,相同接种量下猪粪∶秸秆为1∶1产气量均高于2∶1试验组,且在1∶1的比例下,随接种量的增加其累积VS甲烷产气量也随之增加,因此,猪粪∶秸秆为1∶1,接种量为50%时达到最佳发酵效果。
属水平上的细菌群落种类丰富,梭菌属(Clostridium,梭菌目,厚壁菌门)为优势菌群,包括Clostridiumsensustricto和Terrisporobacter,这一结果与孔德望[25]等在猪粪厌氧发酵消化液回流体系微生物群落结构特征与产气关系研究中的研究结果一致。各处理组发酵后Simpson指数值变化规律一致均有所减少,表明经过发酵细菌优势菌群的生物量占总生物量的比例在增加。本研究中古菌在属水平上有7个主分类,其中Methanobrevibacter,Methanoculleus和Methanomassiliicoccus为优势古菌。在相同接种量不同原料比例下,细菌、古菌群落结构均较相近,说明在实验条件下猪粪∶秸秆为1∶1或2∶1对干发酵微生物的群落结构影响不大,这与Dennehy[26]等研究结果一致。相同原料配比下不同接种量对细菌的多样性和优势菌群丰度无明显影响,其原因可能是随着发酵的进行,反应底物基质浓度不断降低阻碍了细菌的生长。但对于古菌,在一定范围内接种量的增加有利于古菌多样性和优势古菌丰度的提高。
4 结论
采用批式发酵试验,研究猪粪与秸秆在不同比例下(1∶1;2∶1,VS质量比)接种量分别为发酵底物30%,40%,50%(VS质量比)6个处理对厌氧干发酵产气特性以及微生物群落结构的影响,得到结论如下:
(1)相同接种量下猪粪:秸秆为1∶1产气量均高于2∶1试验组;在相同原料比例下,接种量的提高有利于产气高峰的提前以及VS产气量的增加;猪粪∶秸秆为1∶1,接种量为50%时达到最佳发酵效果。
(2)本研究厌氧发酵微生物中,优势细菌为梭菌属(Clostridium),包括Clostridiumsensustricto和Terrisporobacter;优势古菌为Methanobrevibacter,Methanoculleus和Methanomassiliicoccus,其中Methanoculleus为发酵后新增优势古菌。
(3)不同原料比例对细菌和古菌的群落结构和多样性影响较小,但古菌群落结构对接种量较为敏感,古菌多样性在一定范围内也随接种量的增加而提高。
猜你喜欢
土壤学报(2022年3期)2022-08-26 12:15:26
大自然探索(2022年5期)2022-07-11 03:10:33
知识就是力量(2022年6期)2022-06-16 20:19:36
煤气与热力(2021年12期)2022-01-19 05:19:26
现代畜牧科技(2021年3期)2021-07-21 08:41:48
今日农业(2020年19期)2020-12-14 14:16:52
中国资源综合利用(2018年12期)2019-01-08 02:55:40
猪业科学(2018年4期)2018-05-19 02:04:38
浙江农业科学(2016年11期)2016-05-04 04:16:44
当代化工研究(2016年5期)2016-03-20 16:21:30
