
凝结芽孢杆菌抗葡萄糖分解代谢物阻遏突变株合成培养基的筛选和发酵特性研究
2019-10-25 06:10李媛媛王永红
食品工业科技 2019年19期
李媛媛,王永红
(华东理工大学生物反应器工程国家重点实验室,上海 200237)
乳酸是一种多用途的有机酸,在食品、化妆品、农业和制药等行业都有广泛应用[1],特别是近年来乳酸作为可降解生物基塑料制品聚乳酸材料的单体,其生产研究持续受到广泛关注[2]。凝结芽孢杆菌(Bacilluscoagulans)是一种兼性厌氧的革兰氏阳性菌[3],其可在高温、低pH、不灭菌条件下发酵生产高光学纯度的L-乳酸[4-5],这些特性极大降低了乳酸的生产成本,也让其成为了乳酸工业生产的优势菌种。
工业上生产乳酸通常采用淀粉质水解物作为碳源,但在乳酸生产停止时发酵液中会留有较高浓度的残糖,这不仅会降低糖酸转化率,也致使后续乳酸分离纯化的难度增加[6]。因此,筛选抗分解代谢物阻遏的菌株对降低残糖浓度和后续降解残糖都十分必要。在前期的研究中,本实验室以B.coagulansHL5-C为出发菌株,通过常压室温等离子体(ARTP)诱变技术并结合抗葡萄糖分解代谢物阻遏筛选模型获得了一株抗碳源分解代谢物阻遏能力的菌株B.coagulans5E-1[7]。此突变株5E-1除表现出耐高温、耐酸、高产、产高光学纯度的L-乳酸等良好生物学特性外,还可以在利用葡萄糖的同时快速利用其他糖类,大大缩短了乳酸发酵过程残糖的降解时间和整个发酵周期,提高发酵效率。
为了更好地阐释突变株和原始菌株在生理代谢上的差异,本研究通过出发菌株和突变株发酵特性的对比,从其合成培养基的优化、单一碳源及混合碳源的利用情况等方面考察了两菌株之间的差异,为研究凝结芽孢杆菌碳源分解代谢物阻遏效应的机制及其应用奠定基础。
1 材料与方法
1.1 材料与仪器
B.coagulansHL5-C(由B.coagulansHL5通过实验室进化方法选育得到)、B.coagulans5E-1(由B.coagulansHL5-C通过常压室温等离子诱变方法并结合抗葡萄糖代谢物阻遏筛选模型获得) 均由本实验室保藏;无水葡萄糖、碳酸钙 上海凌峰化学试剂有限公司;L-氨基酸(20种) 上海阿拉丁生化科技有限公司;蛋白胨、酵母膏、牛肉膏 英国OXOID;麦芽糖、木糖、果糖、海藻糖、蔗糖、甘露糖、乳糖、半乳糖 上海生工生物工程股份有限公司;低聚异麦芽糖 南京都莱生物技术公司。
离子色谱HPAEC-PAD 美国Thermo公司;SBA-40E生物传感分析仪 山东省科学院生物研究所;DHP-9082电热恒温培养箱 上海华连医疗器械有限公司;ZWY-211C恒温摇床振荡培养柜 上海智城分析仪器制造有限公司;752型紫外可见分光光度计 上海菁华科技仪器有限公司。
1.2 实验方法
1.2.1 培养基的配制 茄子瓶斜面培养基(g/L):蛋白胨10,酵母膏10,牛肉膏10,无水乙酸钠4,柠檬酸氢二铵2,磷酸氢二钾2,100倍母液(MgSO40.2,MnSO40.2,NaCl 0.03,FeSO40.01)10,吐温80(mL)1,琼脂18,无水葡萄糖40,CaCO310,NaOH调节pH至6.0。
MRS种子培养基(g/L):蛋白胨10,酵母膏10,牛肉膏10,无水乙酸钠4,柠檬酸氢二铵2,磷酸氢二钾2,100倍母液(MgSO40.2,MnSO40.2,NaCl 0.03,FeSO40.01)10,吐温80(mL)1,无水葡萄糖40,CaCO310,NaOH调节pH至6.0。
全物质合成培养基[7](g/L):无水葡萄糖40;20种氨基酸0.5;腺嘌呤0.075,鸟嘌呤0.075,胸腺嘧啶0.075,胞嘧啶0.075,尿嘧啶0.075;生物素0.005,泛酸钙0.005,氯化胆碱0.005,肌醇0.005,叶酸0.005,烟酸0.005,对氨甲苯甲酸0.005,维生素B10.005,维生素B20.005,维生素B60.005,维生素B120.005;乙酸钠2,柠檬酸氢二铵1,K2HPO43,KH2PO43,(NH4)2SO43,CaCl20.1,MgSO40.5,MnSO40.05,FeSO40.02,CoSO40.01,CuSO40.01,NiSO40.01,ZnSO40.01,(NH4)6Mo7O240.01,NaOH调节pH至6.0。
合成培养基MCDM3++[8](g/L):无水葡萄糖40;丙氨酸0.5,精氨酸0.5,半胱氨酸0.5,谷氨酸0.5,组氨酸0.5,亮氨酸0.5,异亮氨酸0.5,甲硫氨酸0.5,苏氨酸0.5,酪氨酸0.5,缬氨酸0.5;叶酸0.005,维生素B10.005;鸟嘌呤0.075;乙酸钠2,柠檬酸氢二铵1,K2HPO43,KH2PO43,MgSO40.5,MnSO40.05,FeSO40.02,NaOH调节pH至6.0。
1.2.2 凝结芽孢杆菌的种子制备和发酵培养 利用本实验室冷冻保藏的凝结芽孢杆菌甘油管,取其内菌液250 μL均匀涂布于茄子瓶斜面培养基上,在55 ℃培养箱内进行第一次活化,24 h后转接至新斜面,培养14 h,完成二次活化,用50 mL灭菌水将活化菌株从茄子瓶斜面培养中洗脱下来后接入(按15%接种量(V/V))种子摇瓶培养基中,55 ℃ 100 r/min振荡培养12 h,种子制备过程完成。
发酵培养:种子液4000 r/min离心7 min,去上清并用无菌水重悬,相同条件下二次离心、去上清,并用与原种子液等体积的无菌水重悬、振荡均匀,上述菌液按10%(V/V)的接种量接入发酵摇瓶培养基,摇床(55 ℃,100 r/min)振荡培养。
1.2.3 检测方法 菌体光密度:紫外-可见分光光度计在波长620 nm处测定;葡萄糖、乳酸含量:使用SBA-40E生物传感分析仪测定(定量前需将样品稀释至量程范围内,葡萄糖测定范围为0~1 g/L,乳酸测定范围为0~0.5 g/L);海藻糖、半乳糖、甘露糖、果糖、异麦芽糖和麦芽糖的测定[9]:利用戴安IC-3000离子色谱系统检测,搭配Dionex Carbopac PA 200分离柱(3 mm×250 mm)和Dionex Carbopac PA 200保护柱(3 mm×50 mm),淋洗液体系由18 MΩ超纯水、200 mmol/L NaOH溶液和1 mol/L CH3COONa溶液三部分组成,检测系统需要实时接入氮气,防止空气组分(主要是CO2)与淋洗液接触、反应。
1.2.4 合成培养基的筛选方法
1.2.4.1 出发菌HL5-C合成培养基关键组分的确定 大类物质补全添加实验:基础培养基MCDM3++主要包括氨基酸、维生素、碱基三大类,其中含有11种氨基酸、2种维生素、1种碱基(详见1.2.1)。在MCDM3++基础上,参照全物质合成培养基组分进行上述三类物质的补全添加实验,初步确立HL5-C的生长和代谢营养需求。例如,全物质合成培养基含有20种氨基酸,若培养基添加组分为氨基酸,则指的是在MCDM3++的11种氨基酸基础上外源添加余下的9种氨基酸。大类物质补全添加实验在摇瓶中进行,55 ℃ 100 r/min振荡培养12 h,测定菌体HL5-C的光密度(OD620)。
单一碱基添加实验:在合成培养基MCDM3++的基础上,分别外源添加胞嘧啶C、尿嘧啶U、胸腺嘧啶T、腺嘌呤A中的一种作为实验组培养基,对照组培养基中4种碱基(C、U、T、A)均添加,55 ℃ 100 r/min振荡培养12 h,测定菌体HL5-C的光密度(OD620),并将筛选出的合成培养基命名为MCDM4。

表1 培养基组分设计及HL5-C生长结果Table 1 Medium component design and HL5-C growth results
1.2.4.2 突变株5E-1合成培养基的确定 大类物质补全添加实验:同1.2.4.1,在MCDM4基础上,参照全物质合成培养基组分分别进行氨基酸、维生素和碱基的补全添加实验,确定影响5E-1生长和代谢的关键物质是氨基酸、维生素、碱基中的一类或几类。
单氨基酸添加实验:参照全物质合成培养基中的20种氨基酸,在MCDM4的基础上进行余下9种氨基酸(谷氨酰胺、甘氨酸、赖氨酸、苯丙氨酸、天冬氨酸、天冬酰胺、脯氨酸、丝氨酸和色氨酸)的单一添加实验。培养基中每次添加一种氨基酸,若菌体光密度(OD620)提高,则将该种氨基酸加入基础培养基中,再一一进行余下氨基酸的添加;若OD620无明显变化,则将该氨基酸剔除,最终获得对菌株生长影响明显的氨基酸种类,并将筛选出的合成培养基命名为MCDM5。
1.2.4.3 合成培养基的验证 突变株5E-1种子液分别接入合成培养基MCDM5和全物质合成培养基中,5 L发酵罐(55 ℃ 100 r/min,通气量7.2 L/h)培养19 h,测定5E-1在两种培养基中的OD620和乳酸产量。
1.2.5 出发菌株与突变株利用单一碳源的乳酸发酵 在1.2.4.2优化的合成培养基MCDM5上,分别考察原始菌株HL5-C和突变株5E-1利用不同碳源作为底物的情况。其中对照组以40 g/L葡萄糖作为单一碳源,实验组则分别以40 g/L麦芽糖、低聚异麦芽糖、果糖、甘露糖、半乳糖、乳糖、海藻糖、阿拉伯糖、木糖和蔗糖作为单一碳源。55 ℃ 100 r/min,培养12 h,如果实验组的细胞密度(OD620)高于对照组的1/3,则认为此菌株能够有效利用该碳源。
1.2.6 出发菌株与突变株利用混合碳源的乳酸发酵 出发菌株、突变株利用的混合碳源分别由可被HL5-C、5E-1代谢的非葡萄糖碳源(即1.2.5实验结果)和葡萄糖组成,非葡萄糖碳源浓度为20 g/L,葡萄糖浓度为40 g/L。将HL5-C和5E-1在各种混合碳源中进行培养,在培养基中葡萄糖尚未耗竭的前提下,测定两菌株对非葡萄糖碳源的利用及乳酸生产情况。
1.2.7 抗葡萄糖分解代谢物阻遏效应的验证 混合碳源(30 g/L葡萄糖和12 g/L异麦芽糖)作为底物分别加入到HL5-C和5E-1的合成培养基中,5 L发酵罐(55 ℃ 100 r/min,通气量7.2 L/h)培养21 h,分别测定OD620、葡萄糖消耗量、异麦芽糖消耗量和乳酸产量。
2 结果与分析
2.1 合成培养基的筛选
合成培养基(Chemically defined medium,CDM)的成分明确且可以定量检测[10-11],根据合成培养基上的发酵过程,能够明确发酵底物的代谢流向,从而解析不同的代谢表型差异。本课题在前期研究中利用单因素缺失及模型验证的实验方法对B.coagulans36D1的合成培养基(MCDM3++)进行了优化[8],但是此合成培养基却无法满足出发菌株HL5-C和突变株5E-1的生长。因此,本研究在MCDM3++基础上进一步进行优化,从而保证HL5-C和5E-1的正常生长和代谢。
2.1.1 出发菌株HL5-C合成培养基关键组分的确定 参照全物质合成培养基组分进行氨基酸、维生素和碱基三类物质的补全添加实验,培养基设计和结果如表1所示。实验结果表明,仅添加碱基类物质即可大幅提高最终的细胞密度(OD620),因此后续将进一步研究单一碱基对细胞生长的影响。
单一碱基添加实验结果(表2)表明,C、U、T、A的添加均可以让HL5-C正常生长,其中尿嘧啶U的促进效果最为明显,此时获得最大的细胞密度(OD620)和乳酸积累量。综上所述,HL5-C的生长和乳酸生产合成培养基(MCDM4)为:MCDM3++外加0.075 g/L尿嘧啶(U)。
2.1.2 突变株5E-1合成培养基的确定 虽然MCDM4适合于出发菌株HL5-C生长和乳酸生产,但是突变菌5E-1却无法在此培养基上正常生长和代谢。突变株5E-1合成培养基的优化方法如1.2.4.2所述。实验结果表明,氨基酸是影响5E-1生长和代谢的最重要因素,继而在MCDM3++已有的11种氨基酸的基础上进行单氨基酸添加实验,确定苯丙氨酸、赖氨酸、脯氨酸为5E-1生长和代谢所必需的氨基酸,且三种氨基酸必须同时加入才能使5E-1正常生长和代谢(图1)。综上,5E-1的生长和乳酸生产合成培养基(MCDM5)为:MCDM4,外加0.5 g/L脯氨酸、0.5 g/L赖氨酸和0.5 g/L苯丙氨酸。

表2 培养基组分设计及HL5-C发酵结果Table 2 Medium component design and HL5-C fermentation results
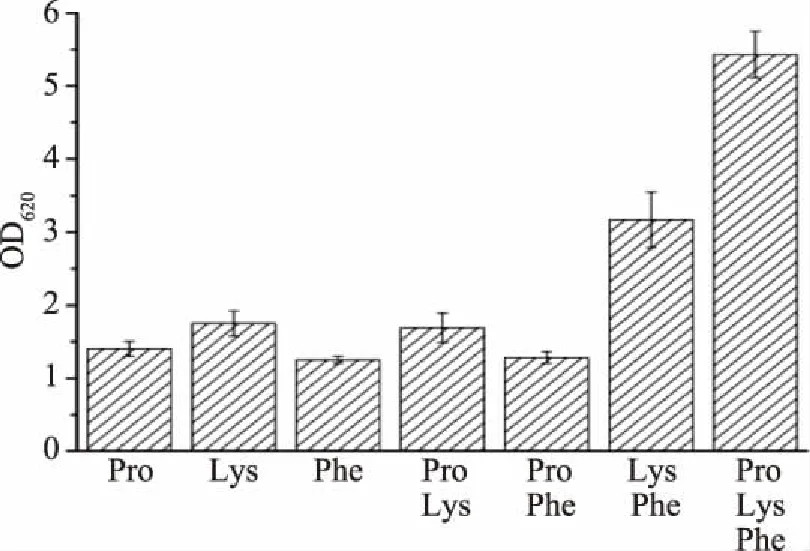
图1 3种氨基酸对5E-1生长的影响Fig.1 Effects of 3 amino acids on 5E-1 growth
凝结芽孢杆菌是产乳酸细菌的一种,产乳酸细菌属于化能异养型微生物,不可以只利用无机氮源生长,在乳酸发酵过程中必须加入菌株生长所需的氨基酸和维生素才能保证菌株生长。突变株5E-1在含有11种氨基酸的培养基MCDM4中无法正常生长和代谢,外源添加的脯氨酸、赖氨酸和苯丙氨酸明显促进了菌株生长,说明此3种氨基酸是突变株5E-1合成培养基中必不可少的物质。
2.1.3 合成培养基MCDM5的验证 为了进一步验证突变株5E-1在MCDM5培养基中的生长和乳酸生产效果,分别利用全物质合成培养基和MCDM5培养基在5 L发酵罐中对5E-1进行发酵实验。从图2中可以发现,利用MCDM5作为基质的5E-1细胞生长和乳酸生产水平与全物质合成培养基相差不大,两种培养基质下5E-1的最大细胞密度(OD620)均达到8左右,最大乳酸积累量也均达到30 g/L以上。因此,经过优化后的合成培养基MCDM5完全可以替代全物质合成培养基作为5E-1生长和代谢的基质。
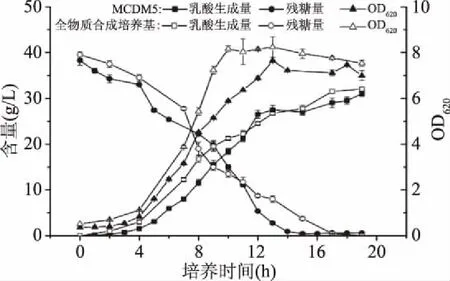
图2 5E-1分别在MCDM5、全物质合成培养基中的批发酵结果Fig.2 Fermentation results of 5E-1 on MCDM5 or complete chemically defined medium
2.2 出发菌株与突变株利用单一碳源产乳酸情况的对比考察
突变株5E-1由原始菌株HL5-C通过ARTP诱变结合抗葡萄糖阻遏效应模型筛选获得,而且在复合培养基中具有良好的表现,但是在合成培养基中突变株5E-1利用不同碳源的情况仍不甚明晰。本研究在上述合成培养基优化的基础上,分别对原始菌株HL5-C和突变株5E-1利用不同碳源作为底物的情况进行了考察。实验结果如图3所示,10种非葡萄糖碳源中有4种糖可被HL5-C单独利用,分别是麦芽糖、果糖、甘露糖和半乳糖。相比之下,5E-1则能单独利用麦芽糖、低聚异麦芽糖、果糖、甘露糖、半乳糖和海藻糖这6种非葡萄糖碳源。可见,5E-1能够比HL5-C利用更多种类的糖,即突变株5E-1的糖代谢谱更广。
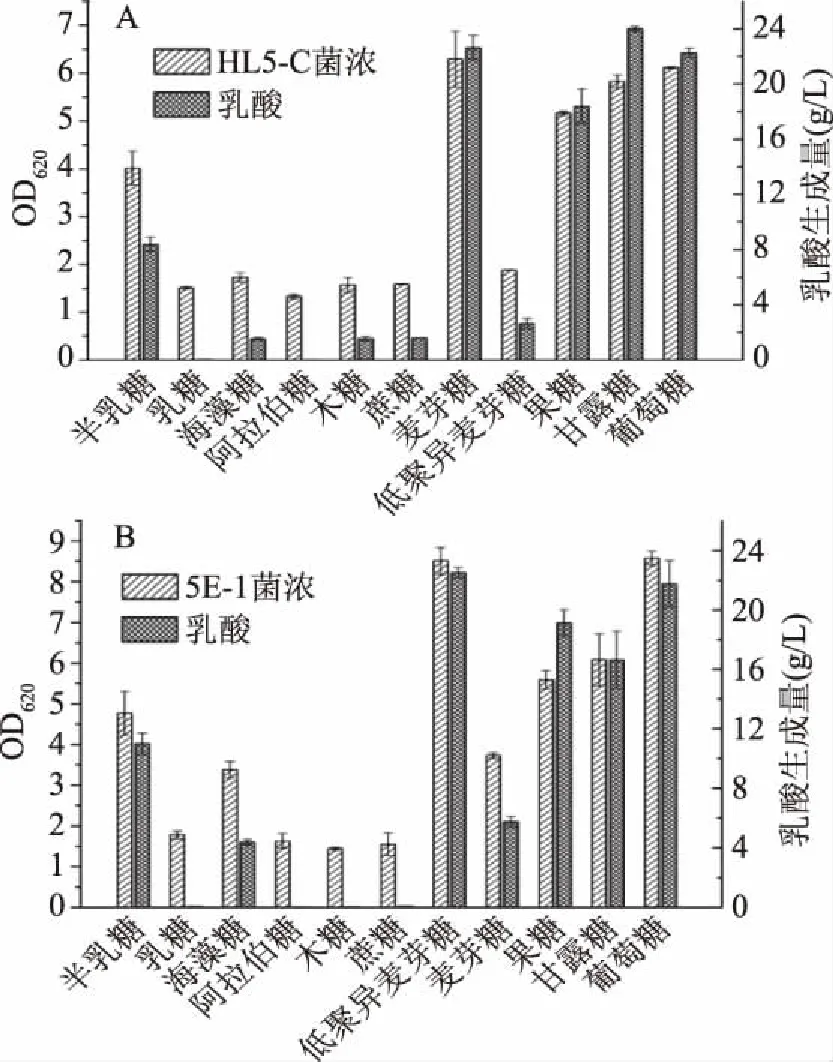
图3 HL5-C(A)和5E-1(B)利用单一碳源的生长情况Fig.3 Growth of HL5-C(A)and 5E-1(B) using a single carbon source
2.3 出发菌株与突变株利用混合碳源生产乳酸研究
出发菌株HL5-C和突变株5E-1在非葡萄糖碳源和葡萄糖组成的混合碳源中的生长和代谢情况如表3所示。实验结果表明,以混合碳源为发酵底物时,原始菌株在葡萄糖存在的前提下对其他非葡萄糖碳源几乎不利用,这是因为葡萄糖代谢引起的分解代谢物阻遏(CCR)效应延滞了菌体对其他糖类的利用[12]。突变株能够同时代谢葡萄糖和其他非速效糖类,说明突变株5E-1能够有效缓解葡萄糖分解代谢物阻遏效应。在实际乳酸发酵过程中,发酵液中主要残糖包括异麦芽糖、麦芽糖和海藻糖[7],而突变株能够有效地在细胞利用葡萄糖的同时消耗这些糖,因此对发酵终点糖含量指标的控制具有重要作用。
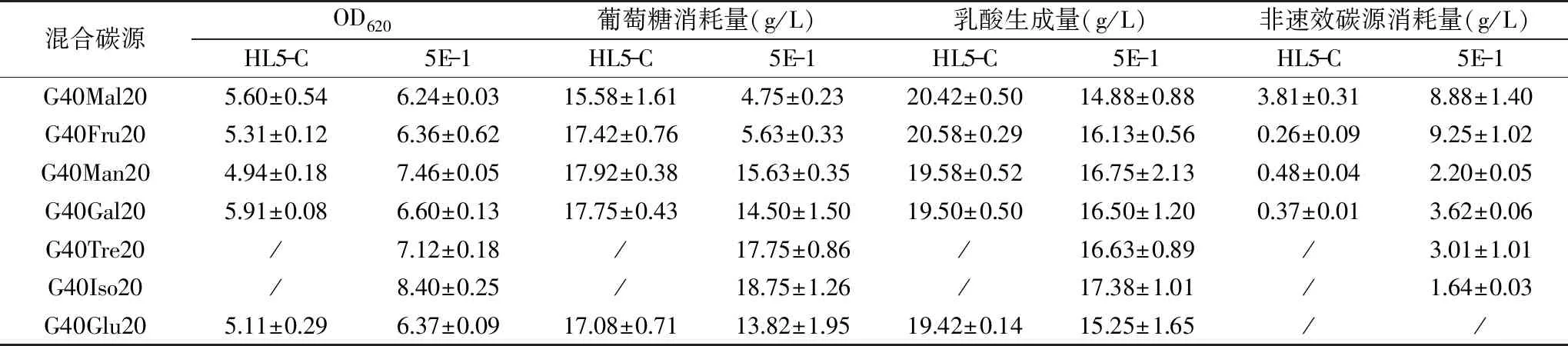
表3 HL5-C和5E-1利用混合碳源的发酵结果Table 3 Fermentation results of HL5-C and 5E-1 with mixed carbon source
注:40 g/L葡萄糖+20 g/L麦芽糖、果糖、甘露糖、半乳糖、海藻糖、异麦芽糖和葡萄糖分别记为G40Mal20、G40Fru20、G40Man20、G40Gal20、G40Tre20、G40Iso20、G40Glu20;/代表未做此组实验,单一碳源利用情况表明HL5-C无代谢海藻糖和异麦芽糖的能力,因此这两种糖未纳入HL5-C混合碳源实验。
2.4 抗葡萄糖分解代谢物阻遏效应的验证
乳酸发酵液中的残糖主要来自于玉米粉水解过程,残糖中含量最高的成分为异麦芽糖,而异麦芽糖不易被菌体自身代谢,因此在5 L罐中通过具体考察原始菌株和突变菌株在合成培养基下对混合碳源(葡萄糖和异麦芽糖)的利用情况,为后续进一步研究突变株抗分解代谢物阻遏的机制奠定基础。
HL5-C和5E-1的生长、异麦芽糖消耗及乳酸生成情况如图4所示。HL5-C和5E-1的葡萄糖消耗速率分别为2.95和3.34 g/L/h,相比之下,5E-1的最大乳酸积累量略高于HL5-C。与摇瓶发酵结果一致,当葡萄糖存在时,基质中的非速效碳源异麦芽糖明显被5E-1消耗,利用量为4.07 g/L;而HL5-C中则不存在同时消耗混合碳源的情况。综上,突变株5E-1是一株去除或缓解了葡萄糖分解代谢物阻遏的凝结芽孢杆菌菌株,能够在利用异麦芽糖的同时,更快地利用葡萄糖,因此可以作为一株潜在的工业乳酸生产菌。

图4 出发菌株和突变株5 L罐规模利用混合碳源发酵结果Fig.4 The fermentation results of mutant and parent strains in 5 L bioreactor with mixed carbon source
3 结论
本文通过大类物质补全添加实验和单因素添加实验,确定影响HL5-C生长的关键物质是尿嘧啶,影响5E-1生长的关键物质是赖氨酸、脯氨酸和苯丙氨酸,以此筛选得到了分别适合出发菌HL5-C和突变菌5E-1生长、代谢的合成培养基MCDM4、MCDM5。HL5-C、5E-1利用单一碳源发酵产乳酸的研究结果表明,突变株5E-1的糖代谢谱更广;两菌株利用混合碳源发酵产乳酸的研究发现5E-1具有抗CCR效应,即在与葡萄糖等速效碳源共存时,5E-1同时还可以利用异麦芽糖、果糖等非速效碳源发酵产酸。突变株5E-1的优良发酵特性—抗CCR效应不仅大大缩短了乳酸发酵过程中残糖的降解时间、提高了发酵效率,也为工业乳酸发酵过程中残糖降解工艺的优化提供了一种新策略。
猜你喜欢
保健与生活(2022年13期)2022-07-06
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
中华养生保健(2020年8期)2021-01-14
天津科技(2020年4期)2020-05-09
作文与考试·小学高年级版(2017年24期)2017-12-14
腹腔镜外科杂志(2016年12期)2016-06-01
老友(2016年3期)2016-05-26
恋爱婚姻家庭·养生版(2016年2期)2016-02-17
