
反刍动物纤维酶制剂作用机理及其应用效果
2019-10-22 02:28闫佰鹏李发弟
草业科学 2019年9期
闫佰鹏,李发弟,2,李 飞
(1.兰州大学草地农业生态系统国家重点实验室 / 兰州大学农业农村部草牧业创新重点实验室 /兰州大学草地农业科技学院,甘肃 兰州 730020;2.甘肃省肉羊繁育生物技术工程实验室,甘肃 民勤 733300)
酶主要是枯草芽孢杆菌属(Bacillus subtilis)、嗜酸乳杆菌(Lactobacillus acidophilus)、植物乳杆菌(Lactobacillus plantarum)等细菌微生物及米曲霉(Aspergillus oryzae)、里氏木霉 (Trichoderma reesei)、酿酒酵母(Saccharomyces cerevisiae)等真菌微生物分泌产生,是一种具有催化活性的生物催化剂[1-3],具有催化效率高、高选择性、绿色无毒害等特性,已被广泛使用于食品生产、医药加工、饲料配制等行业[4],为全球可持续发展作出巨大贡献。如何生产高质量肉、蛋及奶等畜产品是现阶段畜牧领域亟待解决的重要问题,而饲粮是生产高质量、高标准畜产品的关键因素。粗饲料是反刍动物的能量来源之一,在饲粮中的比重与饲养成本、瘤胃健康、生长性能等息息相关,尽管反刍动物拥有强大的瘤胃系统,但消化纤维能力有限,因而限制了粗饲料的充分利用[1]。基于饲养成本降低、安全可行及效益增加等优点使得酶制剂成为反刍动物营养研究的热点科学问题之一[5]。大量学者研究发现饲粮添加酶制剂可显著提高反刍动物生产性能、养分消化率(P<0.05)以及改变瘤胃微生物区系,进而降低饲养成本[6-7],但不同研究间存在一定差异[8],主要是由于酶制剂的利用受饲粮类型、饲养管理、环境因素及自身活力等诸多因素的影响导致动物生产效果存在较大差异,因此有必要系统、全面对酶制剂的作用机制、生化特性及影响因素深入了解与研究,旨在为相关研究提供科学依据。
1 饲用酶制剂的主要作用机理
饲粮碳水化合物主要是由非纤维性碳水化合物和纤维性碳水化合物二大类组成。非纤维性碳水化合物主要包括有机酸、可溶性糖、淀粉、果胶等组成,据报道,除淀粉外其余部分在瘤胃中几乎被完全降解,而淀粉的降解率可高达60%~100%[9],限制反刍动物消化利用饲粮的主要因素为机体对纤维性碳水化合物的消化率。因此,如何提高机体对纤维性碳水化合物的消化利用是反刍动物高效生产的科学瓶颈。
粗饲料的主要成分纤维素与半纤维素是构成反刍动物饲粮不可或缺的纤维性碳水化合物,其在饲粮中比例与动物生产性能、瘤胃健康、饲养成本紧密相关[10]。研究表明植物细胞的“天然屏障”细胞壁使得反刍动物采食植物源性饲料在体内的消化利用率较低[11]。细胞壁由多层复合结构网状连接形成,主要是由纤维素、半纤维素、木质素及结构蛋白组成[12],其纤维素是多个β-D-葡萄糖残基连接形成的高分子化合物[13];半纤维素是由多种糖基、乙酰基等构成的具有分支的高聚糖,在结晶纤维素外表面交织形成复杂的细胞网络干扰水解过程,水解产物主要为五碳糖和六碳糖[14-15];木质素是由苯丙烷单元(C6H3)连接形成的复杂的高分子聚合物,水解效率极其低下,很难被反刍动物消化利用。随着生物技术的飞速发展,酶制剂以无毒害、无副作用被广泛用作于畜禽饲粮添加剂[16-17]。酶制剂在畜禽体内主要通过降解细胞壁,使纤维素、半纤维素等结构多糖水解为可溶性多糖,被机体消化吸收[18]。
纤维素的降解主要由包括内切葡聚糖酶、外切葡聚糖酶、β-葡糖苷酶及一些辅助酶在内的纤维素酶协同进行[19]。通常内切葡聚糖酶随机水解β-1,4-糖苷键,产生大量纤维素短链分子;外切葡聚糖酶主要作用于纤维素分子链的还原端或非还原端水解β-1,4-糖苷键,产生纤维二糖;β-葡糖苷酶主要作用于纤维素分子链的非还原端以纤维寡糖和纤维二糖为水解底物,产生葡萄糖[20]。半纤维素具有复杂的化学结构,因此降解过程需要多种特定酶协同完成。根据其降解部位的不同,半纤维素酶主要有主链降解酶和侧链降解酶二大类:主链降解酶主要有内切-β-1,4-木聚糖酶、内切-β-1,4-甘露聚糖酶与β-1,4-甘露糖苷酶等;侧链降解酶主要有L-阿拉伯呋喃糖苷酶、β-葡糖醛酸糖苷酶等[19]。同样,木质素的降解也需要许多酶的参与,主要有木质素过氧化物酶、多功能过氧化物酶、漆酶及一些辅助酶共同参与降解过程[21]。
瘤胃是反刍动物特有的消化系统,其中栖居数量庞大的细菌、纤毛虫、真菌等微生物,而纤维的消化利用正是通过微生物所分泌的酶进行降解,为机体供能[22]。由于纤维性碳水化合物及木质素形成的复杂网络结构与机体内源酶分泌有限致使水解效率低下,摄入纤维在机体未能充分消化利用[9],随代谢产物排出机体。自20世纪80年代饲料添加剂工艺的兴起,以纤维素酶、半纤维素酶、木聚糖酶和β-葡聚糖酶为主的外源酶被单一或混合应用于反刍动物饲粮中[23],以此增加机体对纤维性碳水化合物的消化利用,此外果胶酶也可以降解纤维素。饲用酶制剂主要是由真菌和细菌等微生物分泌所产生[24],主要通过弥补内源酶分泌不足,将难于消化的纤维素等结构多糖降解为可消化吸收的可溶性糖,同时降低非淀粉多糖在肠道的黏性及消除植酸、α-半乳糖苷等营养物质的抗营养作用,进而提高动物的生长性能、养分消化率,改善瘤胃内环境,有利于动物机体健康(表1)[25]。
2 酶制剂在反刍动物营养的研究
2.1 酶制剂对生长性能的影响
大量学者结合体内、体外和原位降解等方法研究表明,酶制剂的添加显著提高饲料利用率和动物生长性能(P<0.05)[34-35],但受添加方式与剂量、酶活力、饲粮结构与营养水平等大量因素的干扰,酶制剂在动物机体生长性能的作用效果不尽相同。Romero等[26]研究发现荷斯坦奶牛饲喂添加木聚糖酶与纤维素酶处理的全混合日粮(total mixed ration, TMR),木聚糖酶处理组显著增加奶牛干物质采食量(dry matter intake, DMI)、有机物摄入量(organic matter intake, OMI)、粗蛋白摄入量(crude protein intake,CPI)(P<0.05)及提高泌乳早期的产奶量,且对于乳脂及脂肪校正乳(fat corrected milk, FCM)产量有增加的趋势 (P<0.10);冯文晓[36]以杜泊×小尾寒羊杂交F1代公羔为研究对象,分别在水稻秸秆与小麦秸秆饲粮中添加复合酶制剂,研究表明酶处理组均可提高肉羊平均日增重(average daily gain,ADG)、饲料转化效率(feed conversion ration,FCR),显著提高水稻秸秆处理组DMI(P<0.05)。以上研究表明,饲粮中植物细胞壁可被酶制剂水解,释放出大量的水溶性碳水化合物及酚类化合物,这些小分子可溶性糖可以被机体消化吸收利用。与之类似,Togtokhbayar等[27]研究发现给12月龄蒙古公羔(21.6 ± 0.48 kg)饲喂添加纤维素酶与木聚糖酶的饲粮后,纤维素酶对肉羊ADG、FCR及净收入都显著增加(P<0.05)。但也有研究与上述结果不同,Peters等[8]以经产荷斯坦奶牛为研究对象,结果表明酶制剂处理组(6.2 mL·kg-1TMR)对奶牛DMI、OMI和泌乳性能都无显著影响(P>0.05)。Rodriguez等[37]以20月龄公羔为研究对象,在饲粮中添加不同水平纤维酶制剂(0和2 g·kg-1TMR),结果表明纤维酶制剂处理组同样对肉羊采食量、瘤胃发酵参数及生长性能无显著影响(P>0.05)。
综上所述,酶制剂的添加可增加机体消化酶活性,同时可直接作用于日粮中难于消化的纤维素等结构多糖,使其降解为可消化吸收的可溶性糖被机体消化吸收,进而提高动物生长性能。但不同研究间酶制剂添加方式与剂量、饲粮结构、饲养管理等因素的不同,在生产实践中应用效果存在较大变异,因此合理、科学地利用酶制剂、调整饲粮配方对酶制剂在反刍动物生产实践具有重要的科学指导意义。
2.2 酶制剂对养分消化率的影响
植物纤维以侵蚀的方式在反刍动物瘤胃中被水解,而瘤胃微生物定植与穿透能力有限,导致瘤胃微生物分泌的水解酶只能黏附在食糜表面,进而限制纤维降解[38]。Avellaneda等[39]以35和90 d刈割鲜草为日粮来源,每只每天添加0和3 g的酶制剂饲喂萨福克羊,研究发现与0 g组相比,3 g处理组前胃消化系统消化率显著提高(P<0.05),但日粮中OM、DM、NDF与ADF的全消化道消化率无显著影响(P>0.05);赵连生等[28]同样研究发现,在奶牛基础日粮中添加0.10%、0.15%和0.20%复合酶制剂,持续饲喂56 d,研究表明日粮中添加复合酶制剂可显著提高奶牛对日粮DM与NDF的消化率(P<0.05)。与之相似,Rajamma等[40]利用原位降解方法,在不同精粗比的饲粮中添加外源纤维酶制剂,研究表明与精粗比(70∶30)饲粮相比,在精粗比(60∶40)饲粮中添加外源纤维酶制剂能够显著提高饲粮DM、CP、NDF及ADF的降解率(P<0.05)。以上研究结果表明,酶制剂的添加可提高日粮营养成分的消化率,其可能是由于改变了瘤胃微生物区系,选择性提高相应微生物定植与黏附作用,与内源酶协同作用于纤维素、半纤维素,进而增加饲粮中DM、CP、NDF及ADF的降解率。但也有研究表明,在精粗比为(50∶50)饲粮中每只每天分别添加0、8、16和24 g酶制剂于经产荷斯坦泌乳奶牛饲粮中,与0 g组相比,酶制剂处理组DM、CP、NDF及EE的消化率均无影响(P>0.05)[33],其主要原因在适宜条件下(最适pH、温度等)酶制剂能够作用于纤维素、半纤维素的分子键将纤维性碳水化合物水解为可溶性单糖,但机体内环境与酶理想环境存在差异,因此合理选择酶种类,以及保证酶在机体内的活力才能有利于动物生产,增加饲料利用率。此外,酶制剂在机体内的作用效果还受到酶的添加方式与剂量[16]、动物生理阶段、饲粮精粗比及饲粮类型(TMR、颗粒料、青贮料等)等因素的影响。目前针对不同动物、不同生理阶段所需酶制剂种类、添加剂量与方式的研究较少,寻找最佳的作用效果仍是酶制剂在反刍动物营养研究的热点之一。
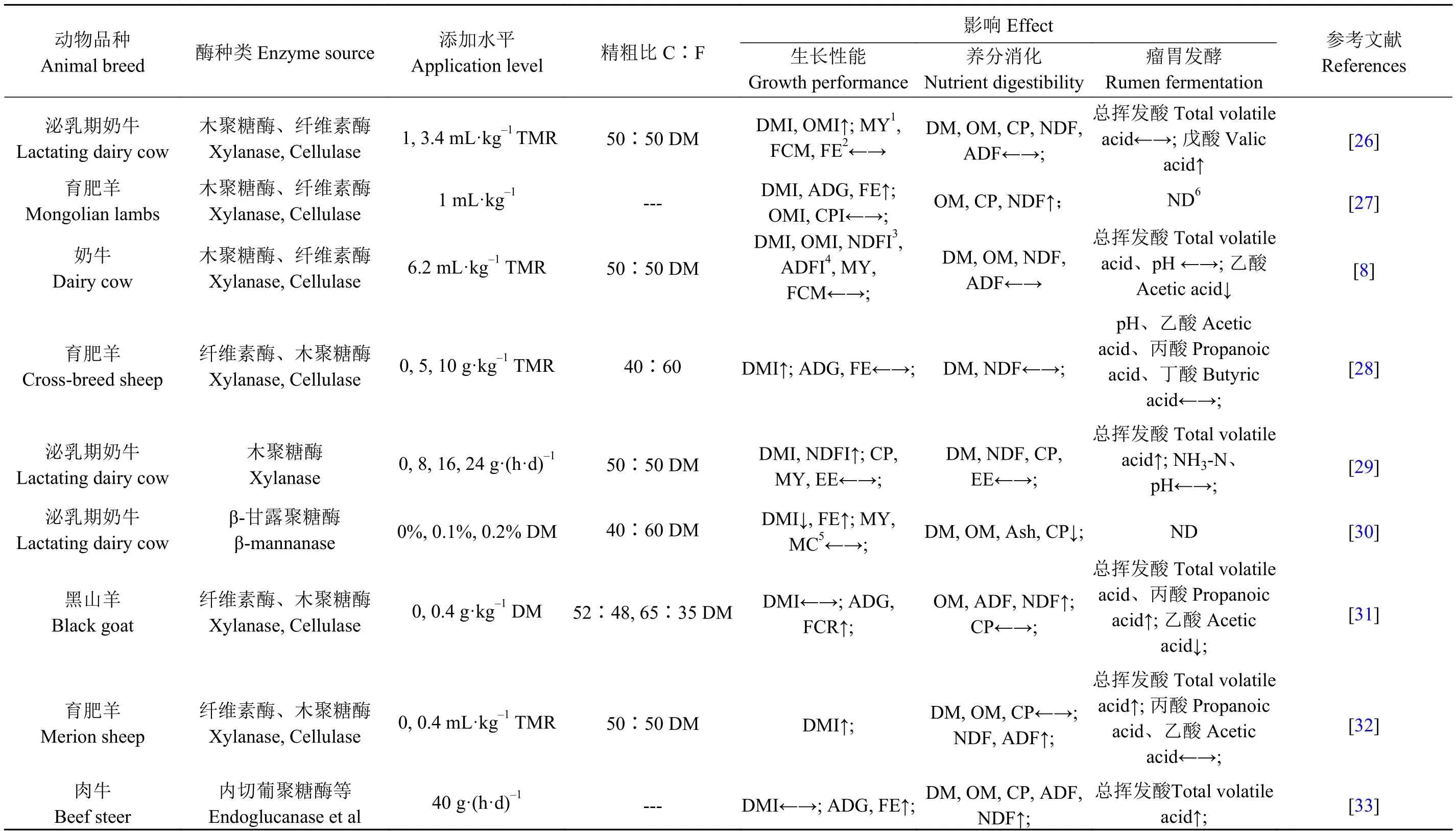
表1 饲粮添加酶制剂对反刍动物的影响Table 1 Effect of feeding enzyme preparation on ruminants
2.3 酶制剂对瘤胃发酵参数及微生物区系的影响
酶制剂通过酶-底物作用轴在机体内发挥作用,进而影响瘤胃微生物群落结构、消化酶活性及改变瘤胃发酵模式[41]。Arriola等[42]以泌乳期荷斯坦奶牛为研究对象,在饲粮中添加酶制剂(3.4 g·kg-1TMR)研究发现,酶制剂组TVFA浓度显著高于对照组 (125.7 vs.114.5 mmol·L-1,P<0.05);Estrada 等[43]同样发现肉羊饲粮中添加酶制剂可显著增加TVFA的浓度 (54.4 vs.52.8 vs.40.2 mmol·L-1,P<0.05)。综上表明,酶制剂的添加可以显著增加TVFA的含量,主要原因是酶制剂有利于食入碳源转化为VFA及参与微生物蛋白合成的过程,进而导致TVFA浓度增加,同时提高了瘤胃微生物数量。国春艳[38]利用PCR-DGGE技术对后备牛饲喂无酶处理TMR与酶处理TMR,结果表明酶处理组瘤胃微生物DGGE图谱条带数量增加,进一步通过序列分析表明酶制剂增加了瘤胃中普雷沃氏菌(Prevotella brevis)和黄色瘤胃球菌(Ruminococcus flavefaciens)的数量(P<0.05);Yusuf等[44]以16只母羊为研究对象,在饲粮中分别添加0、0.2、0.4和0.6 g·kg-1外源纤维酶制剂,持续饲喂56 d,结果发现酶制剂组瘤胃中总细菌数目和真菌数目显著高于对照组(P<0.05);与之类似,李朝云[45]在不同精粗比饲粮(20:80、50∶50、80∶20)条件下添加纤维素酶与半纤维素酶,结果表明在精粗比为(20∶80)的饲粮添加下,纤维素酶的添加显著增加产琥珀丝状杆菌(Fibrobacter succinogenes)和甲烷菌(Methanogens)的数量(P<0.05);半纤维素酶的添加显著降低了原虫数量(P<0.05),对总菌数量、产琥珀丝状杆菌等无影响(P>0.05);瘤胃微生物数量会受到饲粮结构、生理阶段、动物品种及环境因素等诸多自身与外界的影响,但饲粮是影响微生物的主要因素。酶制剂的添加有可能提高了纤维分解菌与淀粉分解菌数量,同时也有研究指出对产琥珀丝状杆菌和溶纤维丁酸弧菌(Butyrivibrio fibrisolvens)属有影响,由于上述菌群的改变导致瘤胃微生物数量发生改变,而这一改变与瘤胃发酵息息相关[46]。综上,饲粮中添加酶制剂可以改变瘤胃发酵模式及瘤胃微生物区系,但也有研究报道发现不会影响TVFA及单个VFA的浓度及瘤胃微生物区系,因此未来有必要从微生物基因组角度深入研究与探索。
3 酶制剂对反刍动物作用效果的主要因素
3.1 酶活力与添加剂量
饲粮结构不同对消化所需酶活力与剂量也不同,酶活力是保证具有催化作用的前提条件之一,其催化活性主要受pH、温度、作用底物等因素的影响[47],而瘤胃pH范围为6.0~7.0,温度为39 ℃左右,无论是pH或温度都具有差异,因此如何保证酶活力在反刍动物生产中至关重要[48]。同时,酶制剂的添加剂量与动物机体作用效果并非成线性关系,Azam等[22]在水牛饲粮中每只每天添加0、10、15和20 g外源纤维酶制剂及Silva等[29]在精粗比为(50:50)饲粮中每只每天分别添加0、8、16和24 g酶制剂饲喂荷斯坦泌乳期奶牛,结果均表明生长性能、养分消化率及瘤胃发酵参数等指标并非与酶制剂剂量有显著线型相关性(P>0.05)。综上所述,确保酶活力及适宜添加水平才能使机体发挥最大的生产潜力,盲目利用只会增加饲养成本且对饲料利用率无影响,因此,科学、合理地利用才会对动物生产具有指导意义。
3.2 添加方式
目前,外源酶主要以添加到TMR[6]、粗饲料[49]、精料补充料、青贮饲料[50]等最为普遍,不同的添加方式导致动物生产中作用效果也有很大差异[38,48]。Sutton等[51]利用瘤胃灌注、喷洒TMR、喷洒精料补充料等方式,研究发现不同的添加方式对奶牛养分消化及瘤胃发酵都存在差异(P>0.05)。与之相比较,Dean等[52]分别将纤维酶制剂添加到青贮料、精料补充料、TMR及粗饲料中饲喂泌乳期奶牛,结果表明精料补充料处理组产奶量与瘤胃pH有降低的趋势,粗饲料处理组TVFA降低,青贮料处理组增加了瘤胃降解速率但TVFA浓度却降低,TMR处理组乙丙比、β-羟丁酸(B-hydroxybutyric acid,BHBA)及血液尿素氮(Blood urea nitrogen,BUN)都显著降低(P<0.05),却增加乳脂与乳蛋白含量;结合以上研究表明,不同的添加方式必然会导致作用效果的差异,合理的添加方式对机体瘤胃健康、生产性能及经济效益息息相关。目前,普遍一致的观点认为酶制剂添加到高水分饲粮(青贮、鲜草)作用效果优于精料、预混料补充料等饲粮成分,主要原因在于水是酶促反应重要的介质,在水解植物细胞壁过程中具有促进作用。
3.3 饲粮结构类型
生长性能、养分消化率及瘤胃发酵参数主要受到饲粮结构的影响。研究表明当饲粮能量浓度不足时,添加酶制剂有利于动物发挥最大的消化潜力[53]。Arriola等[42]通过不同精粗比饲粮(精料补充料:48%和33%)添加酶制剂饲喂泌乳期奶牛,研究发现低精料添加酶可以弥补与高精料的能量差,因此导致生产性能与高精料饲粮一致。杨永明[54]以内蒙古半细毛羯羊为研究对象,在玉米(Zea mays)秸秆型饲粮与精粗比为30∶70饲粮中分别添加外源纤维酶,结果发现玉米秸秆型饲粮酶处理组VFA含量显著提高(P<0.05),其作用效果优于精粗比为30∶70饲粮。综上所述,酶制剂添加于粗饲料型饲粮作用效果优于高精料饲粮,探索饲粮结构-瘤胃功能-生产性能三者之间的关系对酶制剂的利用具有重要意义。
4 小结
酶制剂通过酶-底物作用轴有效水解饲粮中的纤维成分,对畜禽高效生产、机体健康具有重要作用,但受机体自身及外部环境等诸多因素的影响,造成应用效果间存在一定差异。因此,深入探究酶制剂、动物机体与饲粮三者之间的影响因素对酶制剂合理、科学应用具有重要生产意义。
猜你喜欢
吉林畜牧兽医(2022年6期)2022-11-16
养殖与饲料(2021年6期)2021-11-30
中国动物保健(2020年6期)2020-11-27
三农资讯半月报(2020年16期)2020-09-14
科学(2020年2期)2020-08-24
湖南饲料(2019年5期)2019-10-15
猪业科学(2018年5期)2018-07-17
特产研究(2016年3期)2016-04-12
中国粮油学报(2015年5期)2015-02-06
养殖与饲料(2012年10期)2012-12-10
