
新疆果园不同寄主腐烂病菌交互侵染研究
2019-10-22 01:10朱宗财张王斌李亚鹏严海璘仪子博
新疆农业科学 2019年6期
朱宗财,张王斌,2,李亚鹏,严海璘,仪子博
(1.塔里木大学植物科学学院,新疆阿拉尔 843300;2.塔里木大学南疆农业有害生物综合治理重点实验室,新疆阿拉尔 843300)
0 引 言
【研究意义】新疆果园主要是以苹果和库尔勒香梨为主。在我国中西部和西南省区,均将苹果产业列为经济产业[1]。新疆苹果产量在全国苹果产量所占比只有2%左右,但新疆苹果具有明显的产业发展优势,特别是阿克苏地区的冰糖心苹果[2]。库尔勒香梨作为新疆的特色果业,在我国的梨产业中占有重要地位,库尔勒香梨的收入已占到农民人均收入的1/3[3]。腐烂病是普遍存在的一类重要病害,造成许多木质宿主死亡[4]。腐烂病在新疆的发生逐年加重严重影响了该地区林果产业的发展,造成了巨大的经济损失[5-6]。【前人研究进展】苹果树腐烂病(ValsamaliMiyabeetYamada)在新疆阿克苏局部地区发生严重,主要原因是当年冬季干旱少雪,发生冻害,导致了腐烂病严重发生[7]。D.E.Brown-Rytlewski 和 P.S.McManus在美国威斯康星州首次发现苹果腐烂病,并确定为Leucostoma(Cytospora)引起的[8]。Alan R.Biggs等[9]的研究结果表明,苹果腐烂病病原菌的无性型为苹果干腐烂壳囊孢菌(CytosporamandshuricaMiura),属无性孢子类壳囊泡属,有性型为苹果黑腐皮壳菌(ValsamaliMiyabeetYamada),属子囊菌门黑腐皮壳属。王卫雄[10]利用多基因联合分析甘肃省苹果树腐烂病菌的类群。库尔勒香梨树腐烂病菌为梨幹腐烂壳蕉孢菌 (C.carphospermaFr.)属于半知菌亚门壳囊孢属[11-12]。新疆杨树腐烂病病原有性型为子囊菌亚门的污黑腐皮壳属(ValsasordidaNit),其无性型为金黄壳囊孢菌(Cytosporachrysosperm)。病原菌交互侵染主要发生于人与动物之间,而在植物间也发现病原菌的交互侵染。吕蕊花等[13]发现油菜(BrassicanapusL.)和桑葚(FructusMori)的菌核病菌存在交互侵染。朱婕[14]发现新疆库尔勒香梨和杨树腐烂病的症状十分相似,进行室内枝条接种试验发现两种腐烂病存在交互侵染。交互侵染还存在于禾本科作物之间,研究发现稻瘟病在杂交稻和糯稻(OryzasativaL.var.GlutinosaMatsum)之间可以进行交互侵染[15]。黄萎病作为一种棉花(Gossypiumspp.)的主要病害,研究发现黄萎病可以在棉花、向日葵(HelianthusannuusL.)、苘麻(AbutilontheophrastiMedicus)、绿豆(Vignaradiata(Linn.)Wilczek)、榆树(UlmuspumilaL.)、白菜(Brassicarapavar.glabra)和茄子(SolanummelongenaL.)之间交互侵染,所以在棉区要避免这几种作物的混种[16]。在瓜类上也存在交互侵染的情况,例如瓜类白粉菌(Erysiphecucurbi-tacearumZhengetchen)可以在甜瓜(Cucumismelo)、黄瓜(CucumissativusL.)、西葫芦(CucurbitapepoL.)和南瓜(Cucurbitamoschata)上交互侵染[17]。B.K.M.Lakshmi[18]研究了热带水果的重要病害炭疽病(ColletotrichumgloeosporioidesPenz),他将不同寄主来源的炭疽病(Bacillusanthracis)分别在芒果(MangiferaindicaL.)、酸橙(CitrusaurantiumL.)、番荔枝(AnnonasquamosaLinn)、石榴(PunicagranatumL.)、木瓜(Chaenomelessinensis(Thouin)Koehne)、腰果(AnacardiumoccidentalieLinn)和番石榴(PsidiumguajavaLinn)发现这七种水果之间存在交互侵染。【本研究切入点】以新疆果园为微生态环境,研究不同寄主间腐烂病菌是否存在交互侵染。【拟解决的关键问题】采集新疆阿克苏地区五团果园苹果、库尔勒香梨和新疆杨的腐烂病病样,分离纯化,通过科赫氏法则验证后进行交互接种。将菌株的β-tubulin和EF-1α基因序列在GenBank查找并下载同源序列,采用邻结法(Neighbor-joining,NJ)构建系统发育树,进行序列分析。分析新疆果园微生态环境下不同寄主间腐烂病菌是否存在交互侵染情况。
1 材料与方法
1.1 材 料
1.1.1 供试菌株
表1 供试腐烂病菌株信息
Table 1 Information of rot pathogen strains tested
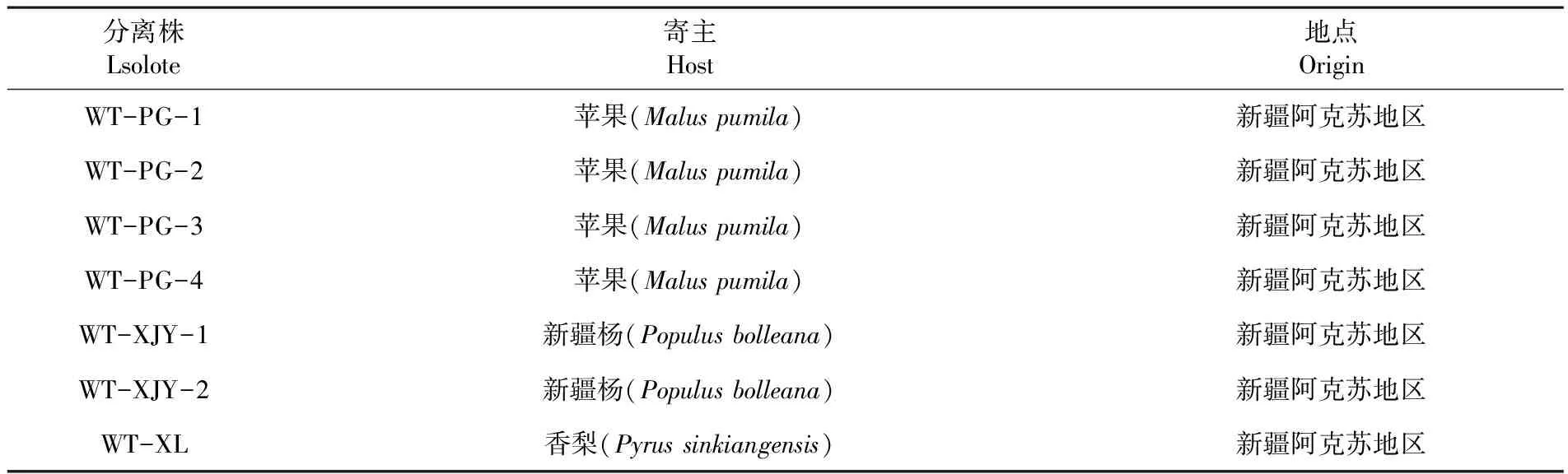
分离株Lsolote寄主Host地点OriginWT-PG-1苹果(Malus pumila)新疆阿克苏地区WT-PG-2苹果(Malus pumila)新疆阿克苏地区WT-PG-3苹果(Malus pumila)新疆阿克苏地区WT-PG-4苹果(Malus pumila)新疆阿克苏地区WT-XJY-1新疆杨(Populus bolleana)新疆阿克苏地区WT-XJY-2新疆杨(Populus bolleana)新疆阿克苏地区WT-XL香梨(Pyrus sinkiangensis)新疆阿克苏地区
1.2 方 法
1.2.1 室内交叉接种
将经过科赫氏法则验证的菌株接种于PDA(Potato Dextrose Agar)平板25℃培养3 d,接种方法参考严海璘等[19]略有改进。在塔里木大学校内分别采集苹果、新疆杨和库尔勒香梨1~2年生枝条,将采集来的枝条用枝剪成长度为10 cm的小枝条。先用纯净水清洗2~3次,再用蒸馏水清洗2~3次,待供试枝条水分干之后,用75%酒精擦拭消毒。将16×2(cm)消毒盘用处理枝条的方法进行处理后,在消毒盘上铺上2层纱布,将其置于无菌操作台用紫外灯灭菌5 min。将5 mm打孔器沾酒精置于酒精灯火焰上进行消毒,在处理过的枝条上进行烫伤打孔,将培养3 d的菌株用5 mm打孔器打成菌饼接种于枝条上,置于消毒盘,重复10次。向消毒盘加入适量的无菌水,用保鲜膜进行封口。将其置于25℃培养箱光照/黑暗(12 h/12 h)培养,每天观察发病情况。
1.2.2 菌丝体的培养与收集
将暗培养3 d的菌株用打孔器取5~6个菌饼放置于含有50 mL PD的150 mL的锥形瓶中,置于25℃摇床160 r/min培养5 d。用无菌处理的纱布过滤菌块,用无菌水冲洗3次,再用灭菌的滤纸吸干水分,经冷冻干燥后,保存于-80℃超低温冰箱中备用。
1.2.3 菌丝体基因组总DNA的提取
采用改进的CTAB法提取菌株基因组总DNA[20-21]。将保存于-80℃超低温冰箱的菌体拿出,加入液氮研磨成粉,加入提前65℃预热的2×CTAB,迅速剧烈震荡,65℃水浴30 min。加入500 μL氯仿(CHCl3)和500 μL的Tris饱和酚,剧烈震荡混匀。将其置于4℃冷冻高速离心机12 000 r/10 min。取上清与新的离心管,加入1 mL氯仿(应等于上清体积),剧烈震荡混匀。将其置于4℃冷冻高速离心机12 000 r/10 min(中间白色层基本消失时可进行沉淀)。取上清至新的离心管中,加入1 mL异丙醇(沉淀DNA)和100 μL的乙酸钠(辅助沉淀),温和颠倒混匀。置于-20℃沉淀2 h,之后置于室温放置5 min,12 000 r/10 min,弃上清。加入1 mL 75%酒精并将沉淀从管底弹起,使其浮与酒精中。12 000 r/10 min,弃上清(超净工作台吹10 min)。加入适量Tris-EDTA缓冲液,-20℃保存。
1.2.4 EF-1α和β-tubulin基因的PCR扩增
采用EF-1α的通用引物728F(5'-CATCGAGAAGTTCGAGAAGG-3')和986R(5'-ACTTGAA
GGAACCCTTACC-3')扩增EF-1α基因片段[22-23];采用通用引物Bt2a(5'-GGTAACCAAATCGG
TGCTGCTTTC-3')和Bt2b(5'-ACCCTCCAGTGTAGTGACCCTTGGC-3')扩增β-tubulin基因片段。表2
EF-1α反应条件为94℃预变性4 min;94℃变性30s,55℃退火45s,72℃延伸30s,40个循环;最后,72℃延伸10 min,4℃保存;β-tubulin反应条件为94℃预变性3 min,94℃变性30s,61℃退火30s,72℃延伸 1 min,共38个循环,接着72℃延伸10 min,最后并于4℃保存。扩增产物经1%琼脂糖进行电泳检测后,由上海生工科技股份有限公司进行测序。
表2 PCR扩增体系
Table 2 PCR amplification system

反应液Reaction Content体积Volume(μL)5'Primer1.03'Primer1.0Taq MasterMix5.0Template1.0ddH2OTotal volume17.025.0
1.2.5 EF-1α和β-tubulin基因的序列对比和系统发育树构建
将各个菌株的EF-1α和β-tubulin基因的序列提交至NCBI中GenBank的BLAST程序与已知序列进行对比并下载同源序列。用DNAMAN中Sequence程序的multiple sequence Alignment进行对比,并用MEGA 7.0.26[24]中的Align DNA分析进行人工调整,删除两端不齐的碱基,采用邻接法(Neighbor-joining,NJ)构建系统发育树,全面分析系统发育树的各分支序列。
2 结果与分析
2.1 枝条交互侵染接种
研究表明,来源于苹果、新疆杨和库尔勒香梨的腐烂病病菌在进行交互侵染时,均能够引起发病。其中寄主为库尔勒香梨的腐烂病菌侵染新疆杨枝条发病最早,在接种第1 d出现病斑,侵染苹果的在第2 d出现病斑。寄主为新疆杨的腐烂病菌的腐烂病侵染库尔勒香梨和苹果枝条发病次之,均在第3 d出现病斑。寄主为苹果的腐烂病菌侵染库尔勒香梨枝条发病较晚,在第4 d出现病斑,侵染新疆杨枝条在第3 d出现病斑。发病率100%的为苹果腐烂病菌侵染库尔勒香梨枝条(B)、杨树腐烂病菌侵染库尔勒香梨枝条(D)和库尔勒香梨腐烂病菌侵染苹果枝条(F)。发病率80%的为杨树腐烂病菌侵染苹果枝条(C)。发病率70%的为苹果腐烂病菌侵染新疆杨枝条(A)。新疆杨枝条接种腐烂病菌后病斑为暗褐色。库尔勒香梨枝条接种腐烂病菌后,病斑初期为暗黄色,后期在病斑周围呈黑色。苹果枝条接种腐烂病菌后病斑为暗黄色。图1

注:A:苹果腐烂病菌侵染新疆杨枝条;B:苹果腐烂病菌侵染库尔勒香梨枝条;C:杨树腐烂病菌侵染苹果枝条;D:库尔勒香梨腐烂病侵染新疆杨枝条;E:杨树腐烂病菌侵染库尔勒香梨枝条;F:库尔勒香梨腐烂病菌侵染苹果枝条
Note: A: Xinjiang Poplar branch infected by Apple canker; B: Fragrant Pear branch infected by Apple canker; C: Apple infected branch by Xinjiang Poplar canker; D: Xinjiang Poplar branch infected by Fragrant Pear canker; E: Fragrant Pear branch infected by Xinjiang Poplar canker; F: Apple branch infected by Fragrant Pear canker
图1 室内交互侵染结果
Fig. 1 The result of the indoor interactive infection
2.2 菌株总DNA的提取及检测
运用改良的CTAB法提取菌株的总DNA结果表明,经1%琼脂糖进行电泳检测后,条带清晰,所提DNA可用于下步实验。图2

图2 DNA提取结果
Fig. 2 The result of the DNA
2.3 β-tubulin和EF-1α基因的PCR扩增
β-tubulin的PCR扩增产物经1%琼脂糖进行电泳检测后,结果显示,条带清楚,大小约为280 bp。图3
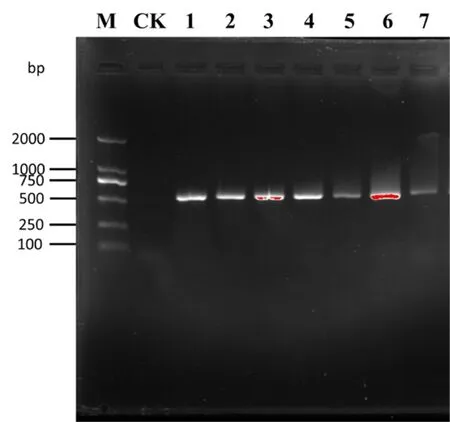
图3 供试菌株β-tubulin扩增结果
Fig. 3 The result of the PCR amplification products of β-tubulin sequences
2.4 EF-1α基因的PCR扩增
EF-1α的PCR扩增产物经1%琼脂糖进行电泳检测后,结果显示,条带清楚,大小约为500 bp。图4

图4 供试菌株EF-1α扩增结果
Fig. 4 The result of the PCR amplification products of EF-1α sequences
2.5 β-tubulin和EF-1α序列
将各个菌株的β-tubulin和EF-1α基因的序列提交至NCBI中GenBank的BLAST与已知序列进行对比并下载同源序列。
将下载的同源性较高的16个β-tubulin序列与分离的7个菌株的β-tubulin序列,利用MEGA 7.0.26软件分析。β-tubulin序列分析显示主要分为三大类并且三大类的延展度大于99。其中来源于苹果、新疆杨和库尔勒香梨的腐烂病β-tubulin序列与各自同源序列聚为一支。来源于苹果和新疆杨的腐烂病菌的病原为Valsamalivar,其中来源于苹果的腐烂病菌的病原为Valsamalivar.mail,来源于新疆杨的腐烂病菌的病原为Valsamalivar.pyil。来源于库尔勒香梨的腐烂病菌的病原为valsasordida。苹果树腐烂病菌的8个同源序列寄主都为苹果,而新疆杨腐烂病菌的5个同源序列中寄主有梨和苹果。库尔勒香梨腐烂病菌的3个同源序列寄主都为杨树。表3,表4
将下载的同源性较高的10个EF-1α序列与分离的7个菌株的EF-1α序列,利用MEGA 7.0.26软件分析。EF-1α序列分析显示主要分为三大类并且三大类的延展度大于99。EF-1α序列分析结果显示苹果树腐烂病菌的5个同源序列寄主都为苹果,而新疆杨腐烂病菌的3个同源序列都为苹果。库尔勒香梨腐烂病菌的2个同源序列寄主都为杨树。
表3 供试菌株β-tubulin序列的同源序列
Table 3 Homologous sequence of the β-tubulin sequence of tested strains

分离株Isolate同源序列Homologous sequence相似度SimilarityWT-PG-1,WT-PG-2, WT-PG-3,WT-PG-4JQ900342、JQ900344、JQ900348、JQ900349JQ900352、JQ900355、JQ900357、KT934371超过 98%WT-XJY-1,WT-XJY-2 JQ900358、JQ900364、JQ900366、KT934372、KT93436999%WT-XLKT590410、KT934375、KT934376超过 96%
表4 供试菌株EF-1α序列的同源序列
Table 4 Homologous sequence of the EF-1α sequence of each isolate

分离株Isolate同源序列Homologous sequence相似度SimilarityWT-PG-1,WT-PG-2, WT-PG-3,WT-PG-4JQ900314、JQ900317、JQ900319、JQ900320JQ900323超过97%WT-XJY-1,WT-XJY-2KX273798、KX273819、KX27381099%WT-XLJX438549、TX438550超过 96%
在基于β-tubulin序列和EF-1α序列构建的系统发育树中,7株不同寄主来源的菌株主要分为两大支分别是Valsamalivar和valsasordida,其中苹果的腐烂病菌为Valsamalivar.mail,寄主为新疆杨的为Valsamalivar.pyil。其中寄主为苹果和新疆杨的腐烂病菌同属一大支,相较于寄主为库尔勒香梨的腐烂病病菌亲缘关系近。图5,图6
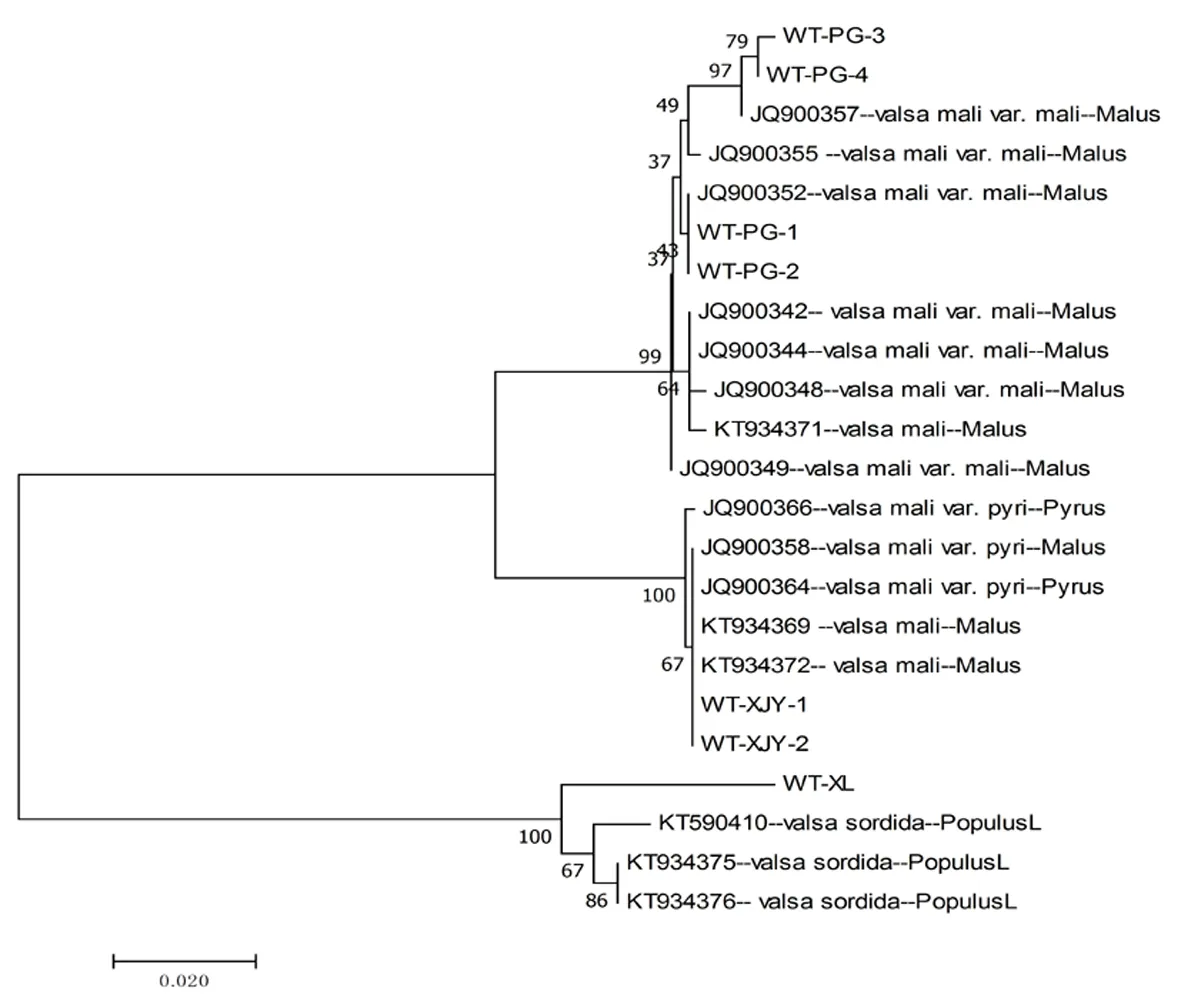
图5 β-tubulin序列构建的系统发育树 (NJ)
Fig. 5 Phylogram using β-tubulin equences data
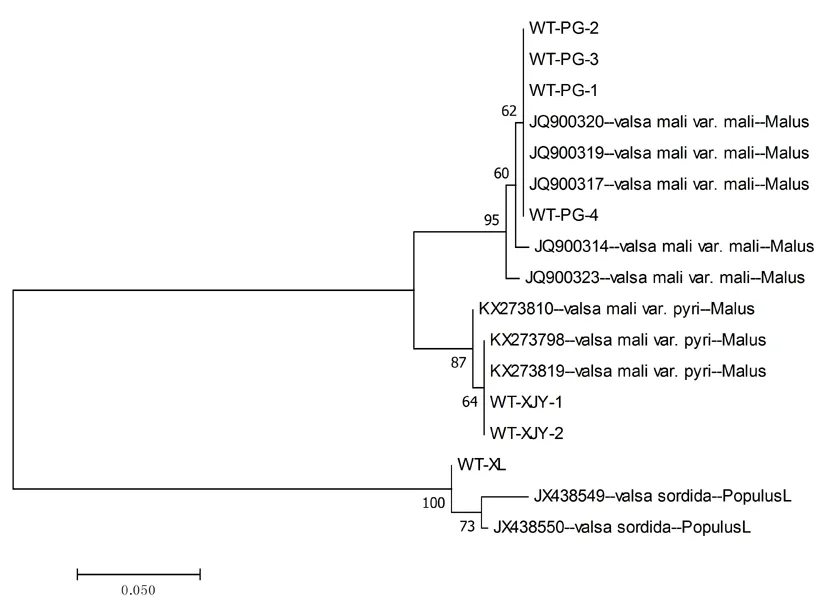
图6 EF-1α序列构建的系统发育树 (NJ)
Fig. 6 Phylogram (NJ) constructed by EF-1 α sequence data
3 讨 论
研究以新疆苹果园为微生态环境,研究不同寄主间腐烂病菌是否存在交互侵染。在新疆独特的绿洲农业中形成了多种林木共存的特点,在一定程度上增加了不同寄主间腐烂病交互侵染的频率。在对采集来的苹果、新疆杨和库尔勒香梨树腐烂病病样进行分离纯化后,经过科赫氏法则验证,进行交互接种,发现均可成功侵染。除了室内接种分析外,还利用腐烂病菌株的β-tubulin基因和 EF-1α基因序列在NCBI下载同源序列,利用邻结法进行建树对腐烂病菌进行分析发现,寄主为苹果的腐烂病菌的同源序列寄主为苹果,但寄主为新疆杨的同源序列的寄主有苹果和梨,寄主为库尔勒香梨的同源序列寄主为杨树,结合交互侵染实验可以证明这腐烂病菌在苹果、新疆杨和库尔勒香梨之间可交互侵染。有研究表明不同寄主来源的腐烂病菌在不同的寄主之间存在交互侵染[25-26]。唐俊煜等[11]研究了枣园不同林木腐烂病之间的亲缘关系,结果显示不同寄主的腐烂病可能存在交互侵染。与研究结果一致。交互侵染不仅存在于林果腐烂病之间,在其他作物间也有发生。有研究表明,马铃薯和向日葵的大丽轮枝菌也存在交互侵染[27]。引起小麦、玉米、水稻、谷子和高粱等作物纹枯病这5种禾谷类作物均可交互侵染[28]。西瓜与棉花的枯萎病菌、小麦与水稻的霜霉病、玉米与高粱的丝黑穗菌之间存在交互侵染[29-31]。Phoulivong S W等[32]从辣椒、咖啡、龙眼、芒果、木瓜和辣椒的果实中分离出炭疽菌菌株,进行了交互侵染,结果发现均能够进行交互感染。O.M.Olanya等[33]在马铃薯种植地的毛龙葵(Solanumsarrachoides)上发现了马铃薯晚疫病(Phytophthorainfestans(Mont.)deBary),研究发现马铃薯晚疫病在毛龙葵与马铃薯上存在交互侵染。Mark L.Farman[34]研究发现马唐、水稻或狗尾草的灰斑病分离株不会引起黑麦草(Loliumperenne)灰斑病流行,引起黑麦草灰斑病是由P.grisea种群引起的。
这些存在交互侵染的作物首先在地理位置上来说相邻种植,而且在遗传距离上来说也较近。进而造成这些作物在发生一些病害时会存在交互侵染。在新疆南疆地区果园栽培模式下,尤其在阿克苏地区,苹果果园、库尔勒香梨园和红枣园临近种植的现象十分普遍,个别果园甚至存在多个树种共存的现象,而杨树作为行道树和果园农田防护林随处可见。由于新疆杨耐旱成活率高的特点,进而果园大都以新疆杨为防护林。所以在一定程度上增加了林果树木腐烂病的交互侵染。研究发现腐烂病菌在苹果、新疆杨和库尔勒香梨之间存在交互侵染,因此,发生腐烂病的果园可通过周边防护林向未发生的腐烂病果园传播。在进行果园腐烂病防治时,应该充分考虑存在交互侵染的现象,把果园周围防护林也纳入防治范围。当周围果园腐烂病严重时,要加强防范意识,提前做好预防措施。新建果园时应该更加关注病害之间的交互侵染的问题,使果园潜在病害发生率降到最低,以达到增产增收的目的。研究在室内条件下,研究了林果树腐烂病菌在苹果、新疆杨和库尔勒香梨可交互侵染,但是其在田间的侵染行为尚需验证和其流行规律也需要进一步深入研究。
4 结 论
4.1 室内利用枝条接种交互侵染发现来源于苹果、库尔勒香梨和新疆杨的腐烂病病菌均能够互相侵染成功。其中寄主为库尔勒香梨的腐烂病菌侵染新疆杨枝条发病最早、寄主为新疆杨的腐烂病菌的腐烂病侵染库尔勒香梨和苹果枝条发病次之、寄主为苹果的腐烂病菌侵染库尔勒香梨枝条发病较晚。进行室内枝条接种还发现有4组发病率为100%,其余两组均在70%以上。
4.2 寄主为苹果的腐烂病菌的同源序列寄主为苹果,但寄主为新疆杨的同源序列的寄主有苹果和梨,寄主为库尔勒香梨的同源序列寄主为杨树。而且这7株不同寄主来源的菌株主要分为两大支分别是Valsamalivar和valsasordida,其中寄主为苹果和新疆杨的腐烂病菌同属一大支,相较于寄主为库尔勒香梨的腐烂病病菌亲缘关系近。
4.3 室内条件下林果树木腐烂病菌在苹果、新疆杨和库尔勒香梨之间存在交互侵染。
猜你喜欢
中国果业信息(2022年4期)2022-11-22
今日农业(2022年16期)2022-09-22
基层中医药(2022年4期)2022-07-22
汉字汉语研究(2021年2期)2021-08-30
今日农业(2020年15期)2020-12-15
汉字汉语研究(2019年2期)2019-08-27
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
农产品市场周刊(2014年35期)2014-12-18
