
新分类系统下长蒴苣苔亚科(苦苣苔科)细胞学研究概述
2019-10-16 10:13杨丽华冯晨徐梅珍孙志霞康明
热带亚热带植物学报 2019年5期
杨丽华, 冯晨, 徐梅珍,2, 孙志霞,2, 康明*
新分类系统下长蒴苣苔亚科(苦苣苔科)细胞学研究概述
杨丽华1, 冯晨1, 徐梅珍1,2, 孙志霞1,2, 康明1*
(1. 中国科学院华南植物园, 中国科学院植物资源保护与可持续利用重点实验室, 广州 510650; 2. 中国科学院大学, 北京 100049)
在最新分类系统框架下对长蒴苣苔亚科(Didymocarpoideae)的染色体资料进行了详细的整理和分析,结果表明,长蒴苣苔亚科的细胞学研究仍存在不足,尤其在种级水平上的研究不足25%,且存在一些属的染色体数据空白的现象。在新的分类系统下,一些修订后的属染色体数目表现出一致性或更加具有合理性,但也存在一些属的染色体数目变异仍十分复杂,如汉克苣苔属()和长蒴苣苔属()。基于已有的染色体数据,对长蒴苣苔亚科内一些重要属的染色体进化模式及其对物种分化的影响进行了探讨,推测染色体数目的多倍化及非整倍化进化可能对各类群的物种分化具有重要作用,但需要今后利用基于DNA探针的荧光原位杂交技术并结合分子系统学和基因组学研究才能深入地解析染色体的进化模式及其对物种分化的影响。
苦苣苔科;长蒴苣苔亚科;染色体数目;染色体进化
苦苣苔科(Gesneriaceae)是唇形目(Lamiales)的一个中等大科,约有3 500种,主要分布在新、旧世界的热带和亚热带地区,只有少数物种可以到达温带地区[1]。在分子系统学尚未介入前,最近的基于形态学依据的苦苣苔科分类系统是Burtt等[2]的3亚科系统,即大岩桐亚科(subfam. Gesnerioideae)、木岩桐亚科(subfam. Coronantheroideae)和苦苣苔亚科(subfam. Cyrtandroideae)。随着分子系统学数据的不断充实,Burtt等的分类系统中很多不合理的现象表现出来,如很多类群(包括亚科、族及属的范围)为多系或并系,不能反映各类群间的自然演化关系[3–4]。尤其是苦苣苔亚科,近年来大量的属发生了调整或被取消,Möller等[5]和辛子兵等[6]对我国苦苣苔亚科属级水平的撤销、合并、转置、扩增等修订情况进行了详细的介绍。在综合分子系统学证据后,Weber等[7]又提出了一个新的苦苣苔科分类系统,还是将苦苣苔科分为3亚科:伞囊花亚科(subfam. Sanangoideae)、大岩桐亚科和长蒴苣苔亚科(subfam. Didymocarpoideae),但是这3亚科的范围及内部系统关系显然与Burtt等界定的3亚科具有不小的差异。Weber等的长蒴苣苔亚科在范围上与Burtt等界定的苦苣苔亚科和传统上按地理概念划分的旧世界类群相近,Weber等只是将台闽苣苔族(trib. Titanotricheae)转入了大岩桐亚科,但是长蒴苣苔亚科下的系统划分发生了很大的变化。因此,为了避免概念上的混淆,本文将Weber等的Didymocarpoideae翻译为长蒴苣苔亚科,而没有采纳符龙飞等[8]仍翻译为苦苣苔亚科的观点。按照Weber等的分类系统,长蒴苣苔亚科下面分为2族14亚族(表1),地理分布范围为亚洲、非洲热带地区、欧洲地中海地区及澳大利亚东北部,仅1种间断分布于中、南美洲,热带亚洲为该亚科的分布及多样性中心[7,9]。
植物的染色体数目、大小和形态数据具有重要的分类学意义[10]。Helliard等[11]根据不同染色体基数将狭义的海角苣苔属()分为2亚属: subg.(=16)和subg.(=15),且这2亚属的划分后来得到了更多的细胞学数据[12]和分子数据的支持[13],即使在现今广义的海角苣苔属概念下[14][包括纹房苣苔()、双钟苣苔属()、侧裂苣苔属()、非洲堇属()、齿丝苣苔属()、匍茎苣苔属()、球蒴苣苔属()和顶齿苣苔属()的所有物种],这个基于染色体数据划分的2亚属仍然成立,只有少数几个=14的物种(原非洲堇属)被划入subg.;Weber等[15]综合形态和细胞学资料对广义的长蒴苣苔属()进行了重新调整,将其分为3属:狭义的长蒴苣苔属、广义的汉克苣苔属()和侧裂苣苔属(),其中狭义的长蒴苣苔属仍具有变化多样的染色体数目(但不包括=9),广义的汉克苣苔属染色体数目主要为= 9,而侧裂苣苔属的染色体数目主要为=14。早在1923年Oehlkes首次报道了苦苣苔科植物的染色体资料(,2=32)[16],但是直到20世纪60年代,Ratter和他的同事们才对旧世界的苦苣苔科植物开展了大范围的细胞学工作[17–21], 到20世纪末Kiehn等[22–23]大量报道了旧世界苦苣苔科植物的染色体资料。Möller等[24]对苦苣苔科的细胞学研究概况进行了统计分析,发现该科染色体资料仍偏少,且认为Burtt等分类系统下旧世界类群的染色体基数变化复杂;王印政[25]也对苦苣苔亚科的细胞学研究概况进行了总结。Christie等[26]比较了原唇柱苣苔属()及该属基于分子数据修订后各属的细胞学数据,发现原唇柱苣苔属具有复杂的染色体数目变化,但是被分为5属后,除了狭义的汉克苣苔属外,其他几属的染色体数目变得较为一致。近年来,苦苣苔科的染色体资料仍有新增加, 同时,关于苦苣苔科的分类系统也得到重新调整,而现如今在新的苦苣苔科分类系统下其细胞学研究概况尚不清楚。
WebCyte数据库(http://elmer.rbge.org.uk/webcyte/webcyteintro.php)具有目前最全面的苦苣苔科植物染色体数据,且具有统计分析功能,数据也不断更新[24,27]。CCDB数据库(Chromosome Counts Database, http://ccdb.tau.ac.il/browse/)是一个汇集了众多小型数据库的综合性染色体数目查询数据库[28]。本文综合这两个数据库中苦苣苔科植物的染色体资料,补充近年来相关文献里新增加的染色体数据[29–34],在Weber等的新分类系统下对长蒴苣苔亚科的染色体资料进行全面的总结分析,以期揭示该亚科下各分类阶元(亚科、族、亚族及属)的染色体资料特征;探讨在新的分类系统下各分类阶元的染色体数目变异是否变得更加合理。另外,本文对一些重要属的染色体进化模式及其对物种分化的影响进行简要评述。
1 长蒴苣苔亚科染色体计数
1.1 亚族水平的染色体计数
整个长蒴苣苔亚科共有70属约2 250种(含种下单位)植物, 其中53属539种(含种下单位)报道了染色体数目,属级水平为75.71%,种级水平为23.96%, 说明物种的染色体资料还严重不足。族级水平上,盾座苣苔族(trib. Epithemateae)的4亚族都有染色体数目报道,属级水平为85.71%,种级水平为32.88%; 芒毛苣苔族(trib. Trichosporeae)有2亚族[四轮苣苔亚族(subtrib. Tetraphyllinae)和凹柱苣苔亚族(subtrib. Lito- stigmatinae)]未见染色体数目报道,属级水平为74.60%,种级水平为23.83%。芒毛苣苔族的细胞学研究资料比盾座苣苔族要少,重要原因可能是前者属和种数量要远远大于盾座苣苔族。亚族级水平上,钩毛苣苔亚族(subtrib. Loxoniinae)、盾座苣苔亚族(subtrib. Epithe- matinae)、珊瑚苣苔亚族(subtrib.Corallodiscinae)、细蒴苣苔亚族(subtrib. Leptobaeinae)、肋蒴苣苔亚族(subtrib. Didissandrinae)和肿蒴苣苔亚族(subtrib. Loxo- carpinae)报道染色体数目的物种比例低于30% (表1)。
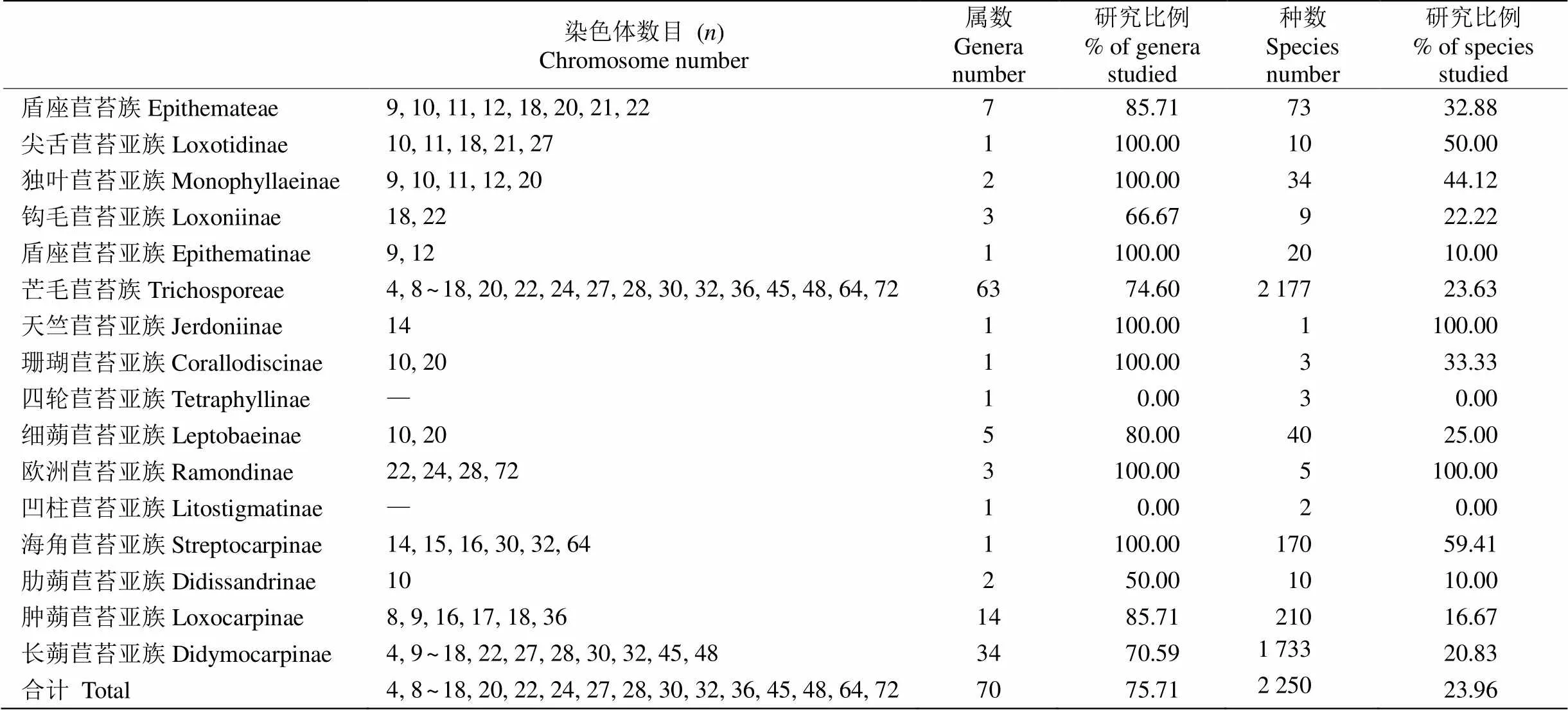
表1 长蒴苣苔亚科的染色体数目和研究概况
1.2 大、中型属的染色体计数
长蒴苣苔亚科含30种以上的大、中型属共有15属,全部为芒毛苣苔族。从表2可见,对这些大、中型属植物染色体数目的研究还存在不足,目前, 只有2属有超过50%以上的物种,即报春苣苔属()和海角苣苔属();有7属报道了染色体数目的物种比例低于15%,即石山苣苔属()、吊石苣苔属()、马铃苣苔属()、半蒴苣苔属()、蛛毛苣苔属()、浆果苣苔属()和红仙苣苔属(),其中浆果苣苔属的物种数量较多, 超过650种,也是苦苣苔科物种数目最多的属[35],因而报道的较少。
1.3 中国苦苣苔科植物的染色体计数
中国是苦苣苔科旧世界类群的重要分布区之一,但是我国对苦苣苔科的细胞学研究起步较晚, 未引起足够重视。Wang等[36]从1998年开始对我国的苦苣苔科植物进行细胞学研究,并率先开展了核型分析。之后,虽然对属级范围进行了广泛的取样分析,但对物种水平的研究还是严重不足。1998-2011年有20属报道了染色体数目,但只有约50种植物[29,31,36–43]。近年来,中国科学院华南植物园对报春苣苔属的细胞学进行了大量的研究,共报道了约100种的染色体数目[32,44–45],同时也对报春苣苔()进行了核型分析[46]。目前, 整个长蒴苣苔亚科尚有17属没有染色体数目资料, 其中8属为我国的特有属,即圆果苣苔属()、凹柱苣苔属()、异唇苣苔属()、异片苣苔属()、扁蒴苣苔属()、双片苣苔属()、圆唇苣苔属()和盾叶苣苔属()。还有一些主要分布在我国的大、中型属的细胞学研究资料严重不足,如吊石苣苔属、石山苣苔属、半蒴苣苔属和马铃苣苔属。
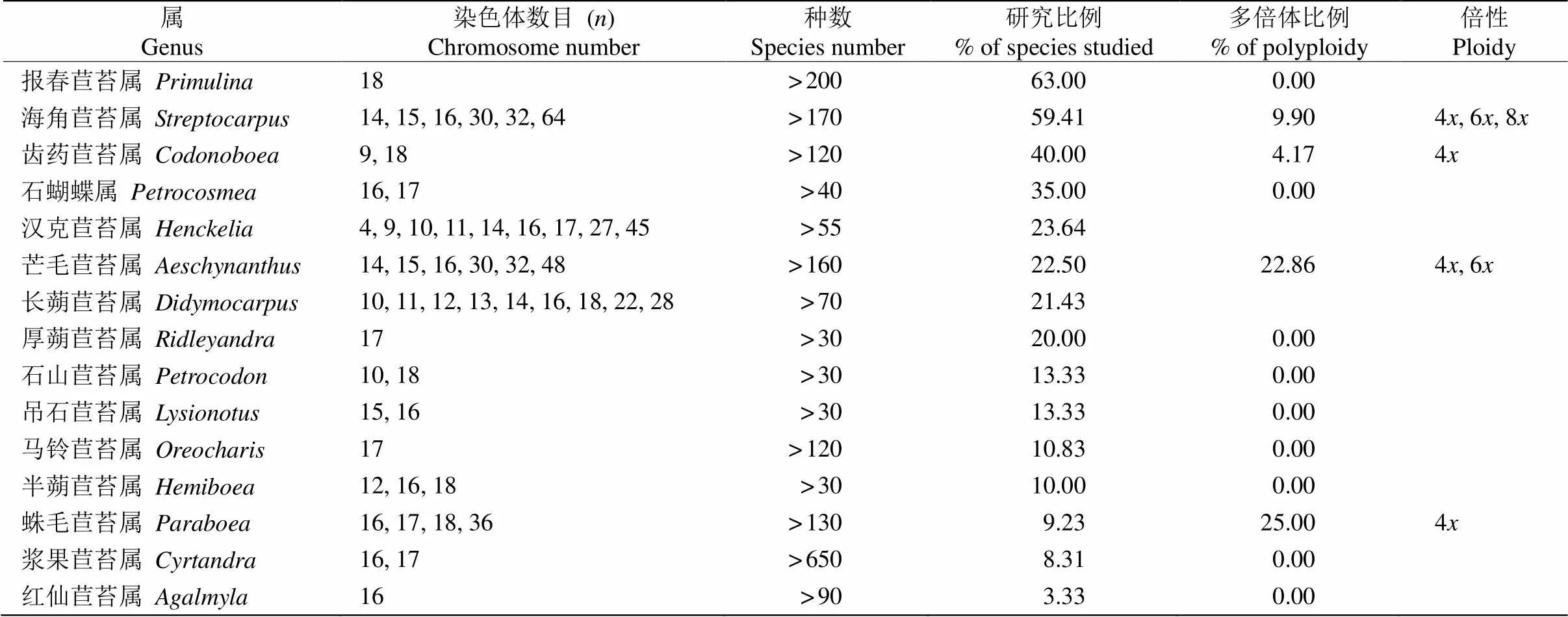
表2 大、中型属(>30种)的染色体数目、多倍体特性
2 长蒴苣苔亚科染色体数目分析
从现有资料看,Weber等分类系统下长蒴苣苔亚科的染色体数目变异十分复杂,但在属级水平上较Burtt等分类系统更具有合理性(表3)。
2.1 盾座苣苔族
Weber等分类系统的盾座苣苔族在范围上与Burtt等和王文采[47]分类系统的尖舌苣苔族(trib. Klugieae)相同。盾座苣苔族属间在形态上差异较大,但其作为1个单系得到了分子系统学数据的支持,且属间的系统关系也得到了较好的解决[48]。依据形态和分子系统学证据,盾座苣苔族进一步可分为4个亚族:尖舌苣苔亚族(subtrib. Loxotidinae)[包括尖舌苣苔属()]、独叶苣苔亚族(subtrib. Monophyllaeinae)[包括异叶苣苔属()和独叶苣苔属()]、钩毛苣苔亚族(subtrib. Loxoniinae)[包括圆果苣苔属()、十字苣苔属(和钩毛苣苔属()]及盾座苣苔亚族(subtrib. Epithematinae)[包括盾座苣苔属()][7]。
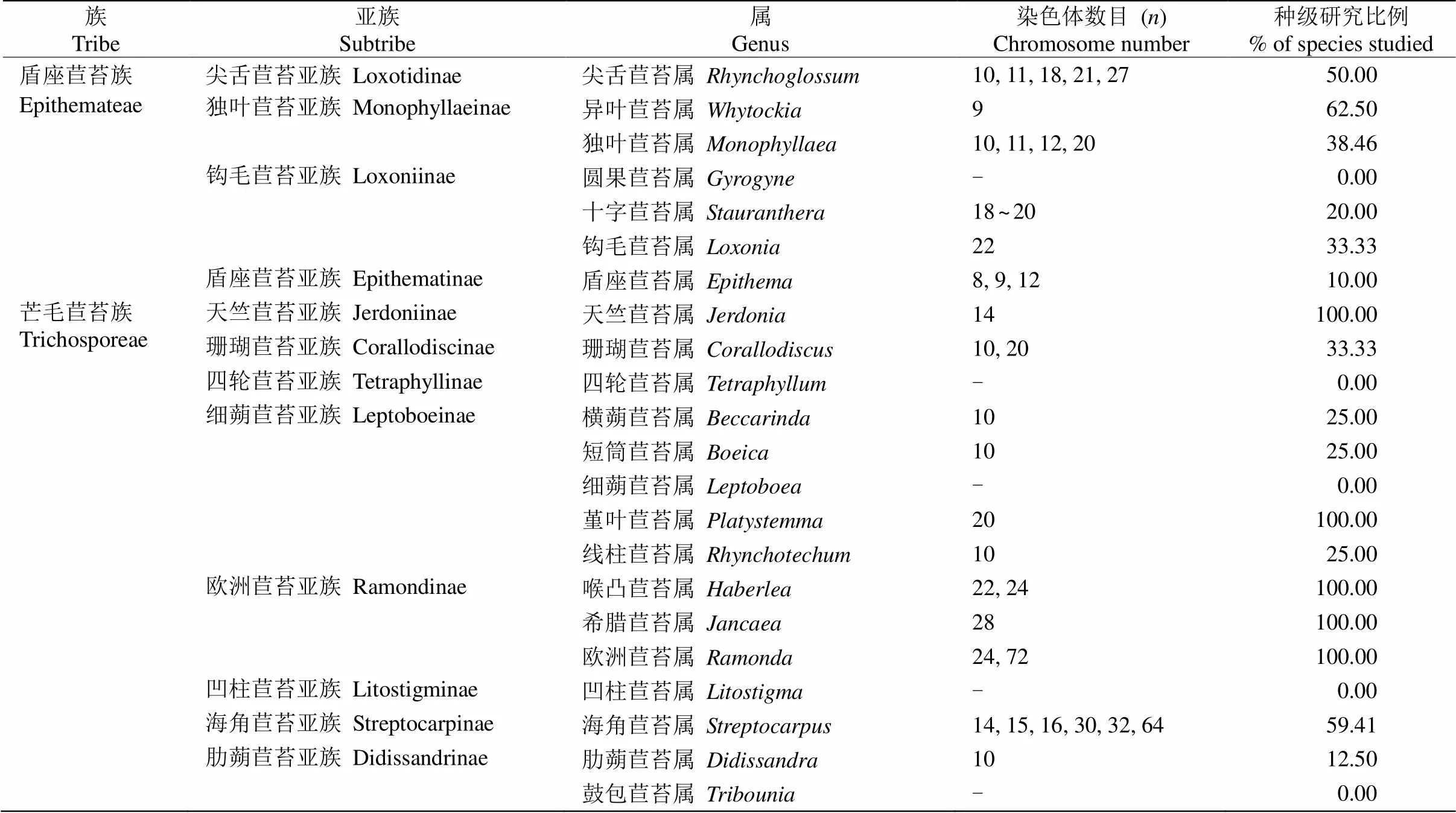
表3 长蒴苣苔亚科的染色体数目和研究概况

续表(Continued)
尖舌苣苔属(=10, 11, 18, 21, 27)、独叶苣苔属(= 10, 11, 12, 20)及盾座苣苔属(=8, 9, 12)的染色体数目变异多样,而异叶苣苔属所有物种的染色体数目较为一致(=9)。钩毛苣苔属(=11)和十字苣苔属(=9, 10)各只有1种报道了染色体数目; 圆果苣苔属尚无染色体数目报道,可以推测盾座苣苔族的染色体基数可能为=9, 10, 11, 12。从染色体数目分布频率来看,=9或10在盾座苣苔族最为频繁。Wang等[36]认为,异叶苣苔属染色体保留了盾座苣苔族中较为原始的性状,因此,=9应该是该族中最原始的基数。
2.2 芒毛苣苔族
Weber等分类系统下的芒毛苣苔族合并了Burtt等分类系统下的浆果苣苔族(trib. Cyrtandreae)、芒毛苣苔族和长蒴苣苔族(trib. Didymocarpeae)的所有物种,但不包括台闽苣苔族。同时,属的范围也发生了大量的调整,Möller等[5]和辛子兵等[6]对我国苦苣苔亚科属级水平上的撤销、合并、转置、扩增等修订情况进行了详细的介绍。另外,基于广泛的分子取样,分布在非洲-马达加斯加的8属植物[纹房苣苔属()、双钟苣苔属()、侧裂苣苔属()、非洲堇属()、齿丝苣苔属()、匍茎苣苔属()、球蒴苣苔属()和顶齿苣苔属()]全部并入了海角苣苔属,致使原本包括9属的海角苣苔亚族(Subtrib. Streptocarpinae)现只含1属[14]。同样基于分子系统学证据,扁冠苣苔属()、套唇苣苔属()、蛛毛苣苔属()和海角苣苔属的范围也进行了重新调整[49]。
扩大的芒毛苣苔族植物染色体数目变异更加多样(表1),包括苦苣苔科中染色体数目最少的物种[斑叶汉克苣苔(), 2=8][26],也包括染色体数目最多的物种(2=144)[30]。虽然整个族的染色体数目变异多样,但是在一些重新调整过的亚族或属的水平上,染色体数目表现出了一致性。珊瑚苣苔亚族和细蒴苣苔亚族植物的染色体数目都为=10 [除堇叶苣苔()和珊瑚苣苔()一些类群发生了多倍化现象导致=20],报春苣苔属的染色体数目为=18,马铃苣苔属的染色体数目为=17, 厚蒴苣苔属的染色体数目为=17。另外,石山苣苔属所有物种的染色体数目严格一致,为=18 (温放未发表数据,私人通信)。作为苦苣苔科中最大的属浆果苣苔属(含超过650种)目前已报道的染色体数绝大多数为=17,仅有3例为=16[50]。蛛毛苣苔属大多数物种的染色体数目为=18,分别只有1例报道=16和=17。石蝴蝶属的染色体数目也较为一致,为=17,只有1例报道为=16[39]。海角苣苔属和芒毛苣苔属的大多数物种染色体数目为=14, 15, 16,但也存在多倍化现象导致=32或64[12,51]。目前,只有汉克苣苔属和长蒴苣苔属在修订后染色体数目变异仍十分复杂,这2属物种的细胞学研究比例都低于25%,汉克苣苔属的染色体数目为=4, 9, 10, 11, 14, 16, 17, 27, 45; 长蒴苣苔属为=10, 11, 12, 13, 14, 16, 18, 22, 28。
3 长蒴苣苔亚科的核型研究
染色体核型分析不仅可为分类学提供更多的信息,而且对了解异源多倍体及认识物种的演化过程都是有帮助的。因为苦苣苔科的染色体普遍较小,通常只有1~2m长[24]。因此,关于苦苣苔科植物的核型研究较少,目前仅有少量我国的物种有详细的核型分析[36–37,40,46,52]。Wang等[36]对尖舌苣苔属进行了全面的核型分析,确认该属的染色体基数=9,其染色体特征在尖舌苣苔族中显示出较原始的性状。孙永健等[52]和杨兴玉等[46]分别对非洲紫罗兰()和报春苣苔的核型进行了分析。Zhou等[40]通过总结及比较分析发现在Burtt等的分类系统下,苦苣苔亚科内各个族内的核型并不一致,尤其是长蒴苣苔族内核型变异复杂。目前相关研究仍太少,限制了在新分类框架下分析讨论核型数据的规律性。在改进制片及染色方法的基础上,对苦苣苔科植物染色体进行更加细致的形态学研究将是今后苦苣苔科植物细胞分类学研究的一个重要方面。
4 长蒴苣苔亚科染色体进化模式及其意义
染色体数目发生多倍化(polyploidy)或非整倍化(dysploidy)变化对植物进化和物种形成具有重要意义[53]。多倍化是植物染色体最常见的进化模式, 被子植物有约35%的物种存在多倍化现象[54]。植物发生多倍化后往往伴随着与祖先种间的生殖隔离, 这样导致快速的物种形成[55]。另外,多倍体通常具有遗传上的优势使其可以占据新的生态位或比祖先种更加具有竞争力[56]。苦苣苔科植物也普遍存在多倍体,尤其是长蒴苣苔亚科[24]。我们从现有的染色体数目可以推测长蒴苣苔亚科2族内都有多倍体的出现,但是多倍体在芒毛苣苔族中更为常见。在海角苣苔属中存在四倍体、六倍体和八倍体(表2),但多倍体只分布在马达加斯加及其附近岛屿,在非洲大陆没有多倍体的报道[12]。海角苣苔属在没有分开为‘非洲支系’和‘马达加斯加支系’之前,多倍体在其属内不同的进化谱系中都存在,这意味着多倍化在属内是平行进化的[24]。芒毛苣苔属()各组都存在多倍体,但是主要出现在芒毛苣苔组(sect.),推测多倍化可能对该组的物种分化具有重要作用[51]。欧洲苣苔属()有3种:、和, 主要分布在巴尔干及利比里亚半岛。Siljak-Yakovlev等[30]的研究表明,和为二倍体(2=2=48),而为六倍体(2=2=144),物种多倍化的同时伴随着基因组大小的加倍,如六倍体的基因组大小约为二倍体的3倍, 因此多倍化可能对该属的物种分化产生重要作用。多倍化同样可以发生在种内不同群体间,珊瑚苣苔()在横断山脉地区的一些群体从二倍体加倍为四倍体,珊瑚苣苔的多倍化阻断了和二倍体间的基因流,对维持种内遗传多样性具有重要意义[34]。
植物染色体非整倍化进化往往由染色体裂变(fission)或融合(fusion)导致染色体数目非整倍性的增加或减少,这种染色体数目的非整倍性变化(dysploidy)不涉及遗传信息的增加或丢失[53]。染色体非整倍化变化在植物中也十分普遍,但染色体非整倍化进化对植物演化的作用相比多倍化的影响被引起的注意仍不足[57]。其实关于多倍化带来的影响还存在不少争论,除了上述有利作用外,多倍化也可能增加多倍体在减数分裂及有丝分裂过程中同源染色体配对和分离的复杂性、改变细胞的物理结构(如多倍体往往需要更大的细胞)、甚至可能导致一些不利的基因结构变化[56]。因此,多倍化有时也可能成为进化上的一条死胡同[58]。Escudero等[57]认为植物染色体非整倍性变化不涉及DNA含量的变化,这可能导致染色体非整倍化进化的谱系相比整倍化进化的谱系在进化历史上可以持续更久。全基因组复制(即多倍化)被广泛认为与被子植物的大爆发相关[59],但Mandáková等[60]认为真正对物种分化起作用的可能是植物基因组在发生复制后伴随着去多倍化的过程,这种去多倍化的过程往往是染色体非整倍性的变化。从现有的染色体数目数据来看,除了一些属的染色体数目较为一致外,大多数属的染色体数目呈非整倍性的变化,或者同时包括整倍性及非整倍性变化,尤其是汉克苣苔属和长蒴苣苔属, 虽然目前这2属都仅有不超过25%的物种有报道染色体数目,但这2属的染色体数目是苦苣苔科中最为复杂的。基于已有的分子系统学证据,Christie等[26]认为汉克苣苔属的染色体数2=18为原始状态,在经历了多次染色体裂变或多倍化后再伴随着融合,从而导致出现2=20, 28, 32, 34的染色体数目,且需要多次直接的融合才能导致2= 8。要想更加详细地探讨一个类群的染色体非整倍进化模式,首先需要通过详细的核型分析确定该类群的染色体基数,然后在稳健的系统发育框架下才能清楚地解析该类群的染色体数目的进化模式,但目前相关研究尚未在苦苣苔科植物中开展。
染色体进化除了数目上整倍性和非整倍变化外,也包括一些结构性的重排,如易位、倒位、插入及缺失等[53]。染色体结构性的重排对物种形成同样具有重要作用[61]。但目前苦苣苔科植物的细胞学研究几乎都仅仅停留在染色体数目的计数上,除了少数研究进行了染色体形态观察外,尚无任何关于染色体结构性的研究,这需要今后利用一些合适的染色方法及荧光标记探针才能清楚地解析染色体结构性重排事件,尤其是对一些近缘种开展染色体结构性重排方面的观察,可以进一步深入的理解染色体重排对物种形成的影响。另外,随着基因组学的发展,全基因组测序也将成为研究染色体结构变化的重要手段[62]。
5 展望
细胞学数据不仅对植物分类有重要参考价值,也对理解植物的起源及进化历史也有重要作用。虽然长蒴苣苔亚科的染色体资料在属级水平上的研究比例超过75%,但是在种级水平上不足25%,尤其是还存在一些重要类群尚无任何染色体数据,如仅分布在我国的一些特有属,即圆果苣苔属、凹柱苣苔属、异唇苣苔属、异片苣苔属、扁蒴苣苔属、双片苣苔属、圆唇苣苔属和盾叶苣苔属。同时,因为苦苣苔科植物的染色体通常较小(1~2m长),导致以前的染色体资料很多存在不准确或不明确,如很多报道的染色体数目为一个范围值。另外,核型研究是确定一个类群的染色体基数的最重要依据, 也对区分一些具有相同染色体数目的近缘种具有参考价值,但是目前在苦苣苔科几乎没有开展这一方面的研究。同时,目前的细胞学研究和分子系统学研究几乎是脱节的,只有整合细胞学及分子系统学数据才能深入的探讨该类群染色体的进化模式。
为进一步提高长蒴苣苔亚科的细胞学研究水平,今后研究重点可以放在以下几方面:1. 在改进压片及染色技术的基础上,进一步补充完善该亚科的染色体资料,同时对以前存在错误或不确定的染色体数据进行改正和核实;2. 采用一些基于DNA探针的荧光原位杂交技术(fluorescenthybrid- dization, FISH),如基于BAC克隆探针荧光原位杂交(BAC-FISH)[63]及基于寡核苷酸探针荧光原位杂交(Oligo-FISH)[64],或基因组分析手段[62],深入分析染色体的形态及结构变化;3. 结合分子系统学手段,将染色体数据标记到系统发育树上,探讨染色体的进化模式,如最近开发出来的ChromEvol软件[65]及ChromoSSE软件[66]可以实现将细胞学数据和分子系统学数据相结合分析。
[1] WEBER A. Gesneriaceae [M]// KADEREIT J W. Flowering Plants: Dicotyledons. Berlin, Heidelberg: Springer, 2004: 63–158. doi: 10. 1007/978-3-642-18617-2_8.
[2] BURTT B L, WIEHLER H. Classification of the family Gesneriaceae [J]. Gesneriana, 1995, 1: 1–4.
[3] WANG X Q, LI Z Y. The application of sequence analysis of rDNA fragment to the systematic study of the subfamily Cyrtandroideae (Gesneriaceae) [J]. Acta Phytotaxon Sin, 1998, 36(2): 97–105. 汪小全, 李振宇. rDNA片段的序列分析在苦苣苔亚科系统学研究中的应用[J]. 植物分类学报, 1998, 36(2): 97–105.
[4] MÖLLER M, CLARK J L. The state of molecular studies in the family Gesneriaceae: A review [J]. Selbyana, 2013, 31(2): 95–125.
[5] MÖLLER M, WEI Y G, WEN F, et al. You win some you lose some: Updated generic delineations and classification of Gesneriaceae: Implications for the family in China [J]. Guihaia, 2016, 36(1): 44–60. doi: 10.11931/guihaia.gxzw201512015.
[6] XIN Z B, FU L F, LI S, et al. The historical changes in the classi- fication of Chinese Gesneriaceae: On the analysis of new taxonomic group qualifiedly published in China and new records of national distribution of this family [J]. Guangxi Sci, 2019, 26(1): 102–117. doi: 10.13656/j.cnki.gxkx.20190307.008. 辛子兵, 符龙飞, 黎舒, 等. 中国苦苣苔科植物的分类系统历史变化——兼论该科植物在我国合格发表的新分类群与国家级分布新记录情况分析 [J]. 广西科学, 2019, 26(1): 102–117. doi: 10.13656/j. cnki.gxkx.20190307.008.
[7] WEBER A, CLARK J L, MÖLLER M. A new formal classification of Gesneriaceae [J]. Selbyana, 2013, 31(2): 68–94.
[8] FU L F, LI S, XIN Z B, et al. The changes of the Chinese names and scientific names of Gesneriaceae in China between Wang’s and Weber’sclassifications for Gesneriaceae [J]. Guangxi Sci, 2019, 26(1): 118–131. doi: 10.13656/j.cnki.gxkx.20190307.003. 符龙飞, 黎舒, 辛子兵, 等. 中国苦苣苔科植物中王文采旧分类系统与Weber新分类系统的名实更替 [J]. 广西科学, 2019, 26(1): 118– 131. doi: 10.13656/j.cnki.gxkx.20190307.003.
[9] LI Z Y. The geographical distribution of the subfamily Cyrtandroideae Endl. Emend. Burtt (Gesneriaceae) [J]. Acta Phytotaxon Sin, 1996, 34 (4): 341–360. 李振宇. 苦苣苔亚科的地理分布 [J]. 植物分类学报, 1996, 34(4): 341–360.
[10] STACE C A. Cytology and cytogenetics as a fundamental taxonomic resource for the 20thand 21stcenturies [J]. Taxon, 2000, 49(3): 451– 477. doi: 10.2307/1224344.
[11] HILLIARD O M, BURTT B L.: An African Plant Study [M]. Pietermaritzburg: University of Natal Press, 1971: 1–171.
[12] JONG K, MÖLLER M. New chromosome counts in(Gesneriaceae) from Madagascar and the Comoro Islands and their taxonomic significance [J]. Plant Syst Evol, 2000, 224(3/4): 173–182. doi: 10.1007/BF00986341.
[13] SMITH J F, KRESGE M E, MÖLLER M, et al. A cladistic analysis ofsequences from representative species ofandsubgeneraand(Gesneriaceae) [J]. Edinb J Bot, 1998, 55(1): 1–11. doi: 10.1017/S0960428600004315.
[14] NISHII K, HUGHES M, BRIGGS M, et al.redefined to include all Afro-Malagasy Gesneriaceae: Molecular phylogenies prove congruent with geographical distribution and basic chromosome numbers and uncover remarkable morphological homoplasies [J]. Taxon, 2015, 64(6): 1243–1274. doi: 10.12705/646.8.
[15] WEBER A, BURTT B L. Remodelling ofand associated genera (Gesneriaceae) [J]. Beitr Biol Pflanzen, 1998, 70(2/3): 293–363.
[16] OEHLKERS F. Entwicklungsgeschichte von[J]. Beih Bot Centralbl, 1923, 39: 128–151.
[17] RATTER J A. Some chromosome numbers in the Gesneriaceae [J]. Notes Roy Bot Gard Edinb, 1963, 24: 221–229.
[18] RATTER J A. A survey of chromosome numbers in the Gesneriaceae of the old World [J]. Notes Roy Bot Gard Edinb, 1975, 33: 527–543.
[19] RATTER J A, PRENTICE H T. Chromosome numbers in the Gesne- riaceae: II [J]. Notes Roy Bot Gard Edinb, 1964, 25(3): 303–307.
[20] RATTER J A, PRENTICE H T. Chromosome numbers in the Gesne- riaceae: III [J]. Notes Roy Bot Gard Edinb, 1967, 27(2): 205–209.
[21] RATTER J A, MILNE C. Chromosome numbers in the Gesneriaceae: IV [J]. Notes Roy Bot Gard Edinb, 1970, 30: 183–187.
[22] KIEHN M, HELLMAYR E, WEBER A. Chromosome numbers of Malayan and other paleotropical Gesneriaceae: I. Tribe didymocarpeae [J]. Beitr Biol Pflanzen, 1998, 70(2/3): 407–444.
[23] KIEHN M, WEBER A. Chromosome numbers of Malayan and other paleotropical Gesneriaceae: II. Tribes Trichosporeae, Cyrtandreae and Epithemateae [J]. Beitr Biol Pflanzen, 1998, 70(2/3): 445–470.
[24] MÖLLER M, KIEHN M. A synopsis of cytological studies in Gesne- riaceae [J]. Edinb J Bot, 2003, 60(3): 425–447. doi: 10.1017/S096042 8603000337.
[25] WANG Y Z. Studies in cytology [M]// LI Z Y, WANG Y Z. Gesne- riaceae Plants in China. Zhengzhou: Henan Science and Technology Press, 2005: 576–600.王印政. 细胞学研究 [M]// 李振宇, 王印政. 中国苦苣苔科植物. 郑州: 河南科学技术出版社, 2005: 576–600.
[26] CHRISTIE F, BARBER S, MÖLLER M. New chromosome counts in Old World Gesneriaceae: Numbers for species hitherto regarded as, and their systematic and evolutionary significance [J]. Edinb J Bot, 2012, 69(2): 323–345. doi: 10.1017/S0960428612000169.
[27] MÖLLER M, PULLAN M. RBGE ‘WebCyte2’: An updated Gesne- riaceae cytology database [DB/OL]. (2016-01-21). http://elmer.rbge. org.uk/webcyte/webcyte.php.
[28] RICE A, GLICK L, ABADI S, et al. The chromosome counts database (CCDB): A community resource of plant chromosome numbers [J]. New Phytol, 2015, 206(1): 19–26. doi: 10.1111/nph.13191.
[29] WANG J C, WANG C C. The genusBlume (Gesne- riaceae) in Taiwan [J]. Taiwania, 2000, 45(4): 355–365.
[30] SILJAK-YAKOVLEV S, STEVANOVIC V, TOMASEVIC M, et al. Genome size variation and polyploidy in the resurrection plant genus: Cytogeography of living fossils [J]. Environ Exp Bot, 2008, 62(2): 101–112. doi: 10.1016/j.envexpbot.2007.07.017.
[31] PUGLISI C, WEI Y G, NISHII K, et al.×(Gesne- riaceae): A natural hybrid from the Shengtangshan Mountains, Guangxi, China [J]. Phytotaxa, 2011, 38: 1–18. doi: 10.11646/phytotaxa.38.1.1.
[32] LIU R R, SKOG L E, LIAO J P, et al. New chromosome counts and their taxonomic implications in(Gesneriaceae) [J].Plant Diver Resour, 2014, 36(1): 13–21. doi: 10.7677/ynzwyj201413044.刘瑞瑞, SKOG L E, 廖景平, 等. 广义报春苣苔属(苦苣苔科)的染色体新计数及其分类学意义 [J]. 植物分类与资源学报, 2014, 36(1): 13–21. doi: 10.7677/ynzwyj201413044.
[33] KOKUBUGATA G, YOKOTA M. The chromosome numbers of four Gesneriaceae species native to Japan [J]. Bull Natl Mus Nat Sci Ser B, 2017, 43(2): 57–61.
[34] ZHOU P, LI J, MÖLLER M. Secondary contact, hybridization and poly- ploidization add to the biodiversity in the Hengduan Mountains, exem- plified by the widespread(Gesneriaceae) [J]. Plant Syst Evol, 2017, 303(5): 587–602. doi: 10.1007/s00606-017-1392-0.
[35] ATKINS H J, BRAMLEY G L C, Clark J R. Current knowledge and future directions in the taxonomy of(Gesneriaceae), with a new estimate of species number [J]. Selbyana, 2013, 31(2): 157–165.
[36] WANG Y Z, GU Z J, HONG D Y. Karyotypes of(Gesne- riaceae) [J]. Acta Phytotaxon Sin, 1998, 36(1): 28–35.
[37] WANG Y Z, GU Z J. Karyomorphology of four species in,and(Gesneriaceae) [J]. Acta Phyto- taxon Sin, 1999, 37(2): 137–142. 王印政, 顾志建. 直瓣苣苔属、筒花苣苔属和吊石苣苔属4个种的核形态学研究[J]. 植物分类学报, 1999, 37(2): 137–142.
[38] LU Y X, SUN X F, ZHOU Q X, et al. Chromosome numbers in ten species in the Gesneriaceae from Yunnan [J]. Acta Bot Yunnan, 2002, 24(3): 377–382. doi: 10.3969/j.issn.2095-0845.2002.03.011.鲁元学, 孙先凤, 周其兴, 等. 云南十种苦苣苔科植物的染色体数目报道[J]. 云南植物研究, 2002, 24(3): 377–382. doi: 10.3969/j.issn. 2095-0845.2002.03.011.
[39] CAO L M, CAO M, TANG X L, et al. Chromosome numbers of 4 species in the Gesneriaceae from Guangxi [J]. Guihaia, 2003, 23(4): 331–333. doi: 10.3969/j.issn.1000-3142.2003.04.012.曹丽敏, 曹明, 唐咸来, 等. 广西苦苣苔科四种植物的染色体数目报道[J]. 广西植物, 2003, 23(4): 331–333. doi: 10.3969/j.issn.1000- 3142.2003.04.012.
[40] ZHOU P, GU Z J, MÖLLER M. New chromosome counts and nuclear characteristics for some members of Gesneriaceae subfamily Cyrtan- droideae from China and Vietnam [J]. Edinb J Bot, 2004, 60(3): 449– 466. doi: 10.1017/S0960428603000349.
[41] JI H, GUAN K Y, LU Y X. Chromosome numbers of eight species in the genus(Gesneriaceae) [J]. Acta Bot Yunnan, 2008, 30 (3): 321–324. doi: 10.3969/j.issn.2095-0845.2008.03.004. 季慧, 管开云, 鲁元学. 石蝴蝶属八种植物的染色体数目报道[J]. 云南植物研究, 2008, 30(3): 321–324. doi: 10.3969/j.issn.2095-0845. 2008.03.004.
[42] ZHAO D K, LU Y X, SHI J F, et al. Seed germination and chromo- some numbers ofand[J]. J Yunnan Agric Univ, 2010, 25(2): 173–177. doi: 10.16211/j.issn.1004- 390x(n).2010.02.019.赵大克, 鲁元学, 石景峰, 等. 吊石苣苔和盾叶粗筒苣苔的种子萌发及染色体数目观察[J]. 云南农业大学学报, 2010, 25(2): 173–177. doi: 10.16211/j.issn.1004-390x(n).2010.02.019.
[43] TAN Y, WANG Z, SUI X Y, et al. The systematic placement of the monotypic genus(Gesneriaceae) based on molecular and cytological data [J]. Plant Diver Res, 2011, 33(5): 465–476. doi: 10. 3724/SP.J.1143.2011.11075.谭英, 王智, 隋学艺, 等. 单种属弥勒苣苔属系统位置研究: 基于分子和细胞学数据[J]. 植物分类与资源学报, 2011, 33(5): 465–476. doi: 10.3724/SP.J.1143.2011.11075.
[44] LIU R R, PAN B, ZHOU T J, et al. Cytological studies ontaxa (Gesneriaceae) from limestone karsts in Guangxi province, China [J]. Caryologia, 2012, 65(4): 295–303. doi: 10.1080/00087114.2012.752920.
[45] KANG M, TAO J J, WANG J, et al. Adaptive and nonadaptive genome size evolution in karst endemic flora of China [J]. New Phytol, 2014, 202(4): 1371–1381. doi: 10.1111/nph.12726.
[46] YANG X Y, LIANG K M, ZHANG X H, et al. Karyotype analysis of an endemic species(Gesneriaceae) [J]. Plant Diver Res, 2012, 34(1): 25–27. doi: 10.3724/SP.J.1143.2012.11126.杨兴玉, 梁开明, 张新华, 等. 国家一级濒危植物报春苣苔核型分析 [J]. 植物分类与资源学报, 2012, 34(1): 25–27. doi: 10.3724/SP.J. 1143.2012.11126.
[47] WANG W T. Gesneriaceae [M]// Flora Rerpublicae Popularis Sinicae, Tomus 69. Beijing: Science Press, 1990: 125–581. 王文采. 苦苣苔科, 尖舌苣苔族[M]// 中国植物志, 第69卷. 北京: 科学出版社, 1990: 125–581.
[48] MAYER V, MÖLLER M, PERRET M, et al. Phylogenetic position and generic differentiation of Epithemateae (Gesneriaceae) inferred from plastid DNA sequence data [J]. Amer J Bot, 2003, 90(2): 321–329. doi: 10.3732/ajb.90.2.321.
[49] PUGLISI C, YAO T L, MILNE R, et al. Generic recircumscription in the Loxocarpinae (Gesneriaceae), as inferred by phylogenetic and morpho- logical data [J]. Taxon, 2016, 65(2): 277–292. doi: 10.12705/652.5.
[50] BÖRGMANN E. Anteil der polyploiden in der flora des Bismarck- gebirges von ostneuguinea [J]. Z Bot, 1964, 52: 118–172.
[51] RASHID M H, JONG K, MENDUM M. Cytotaxonomic observations in the genus(Gesneriaceae) [J]. Edinb J Bot, 2001, 58 (1): 31–43. doi: 10.1017/S0960428601000452.
[52] SUN Y J, CHEN X Q, SUN N, et al. Karyotype analysis of[J]. J Tianjin Agric Univ, 2010, 17(4): 5–8. 孙永健, 陈小强, 孙宁, 等. 非洲紫罗兰的核型分析[J]. 天津农学院学报, 2010, 17(4): 5–8.
[53] de STORME N, MASON A. Plant speciation through chromosome instability and ploidy change: Cellular mechanisms, molecular factors and evolutionary relevance [J]. Curr Plant Biol, 2014, 1: 10–33. doi: 10. 1016/j.cpb.2014.09.002.
[54] WOOD T E, TAKEBAYASHI N, BARKER M S, et al. The frequency of polyploid speciation in vascular plants [J]. Proc Natl Acad Sci USA, 2009, 106(33): 13875–13879. doi: 10.1073/pnas.0811575106.
[55] RIESEBERG L H, WILLIS J H. Plant speciation [J]. Science, 2007, 317(5840): 910–914. doi: 10.1126/science.1137729.
[56] MADLUNG A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools [J]. Heredity, 2013, 110(2): 99–104.
[57] ESCUDERO M, MARTÍN-BRAVO S, MAYROSE I, et al. Karyotypic changes through dysploidy persist longer over evolutionary time than polyploid changes [J]. PLoS One, 2014, 9(1): e85266. doi: 10.1371/ journal.pone.0085266.
[58] STEBBINS G L. Chromosomal Evolution in Higher Plants [M]. London: Arnold, 1971: 1–216.
[59] SOLTIS P S, SOLTIS D E. Ancient WGD events as drivers of key innovations in angiosperms [J]. Curr Opin Plant Biol, 2016, 30: 159– 165. doi: 10.1016/j.pbi.2016.03.015.
[60] MANDÁKOVÁ T, LYSAK M A. Post-polyploid diploidization and diversification through dysploid changes [J]. Curr Opin Plant Biol, 2018, 42: 55–65. doi: 10.1016/j.pbi.2018.03.001.
[61] FARIA R, NAVARRO A. Chromosomal speciation revisited: Rearranging theory with pieces of evidence [J]. Trends Ecol Evol, 2010, 25(11): 660–669. doi: 10.1016/j.tree.2010.07.008.
[62] DOLEŽEL J, VRÁNA J, CÁPAL P, et al. Advances in plant chromo- some genomics [J]. Biotechnol Adv, 2014, 32(1): 122–136. doi: 10. 1016/j.biotechadv.2013.12.011.
[63] SZINAY D, WIJNKER E, van den BERG R, et al. Chromosome evolution intraced by cross-species BAC-FISH [J]. New Phytol, 2012, 195(3): 688–698. doi: 10.1111/j.1469-8137.2012.04195.x.
[64] BRAZ G T, HE L, ZHAO H N, et al. Comparative oligo-FISH mapping: An efficient and powerful methodology to reveal karyotypic and chromosomal evolution [J]. Genetics, 2018, 208(2): 513–523. doi: 10.1534/genetics.117.300344.
[65] GLICK L, MAYROSE I. ChromEvol: Assessing the pattern of chromosomenumber evolution and the inference of polyploidy along a phylogeny [J]. Mol Biol Evol, 2014, 31(7): 1914–1922. doi: 10.1093/molbev/msu122.
[66] FREYMAN W A, HÖHNA S. Cladogenetic and anagenetic models of chromosome number evolution: A Bayesian model averaging approach [J]. Syst Biol, 2018, 67(2): 195–215. doi: 10.1093/sysbio/syx065.
Synopsis of Cytological Studies on Didymocarpoideae (Gesneriaceae) under New Classification System
YANG Li-hua1, FENG Chen1, XU Mei-zhen1,2, SUN Zhi-xia1,2, KANG Ming1*
(1. Key Laboratory of Plant Resources Conservation and Sustainable Utilization, South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650, China; 2. University of Chinese Academy of Sciences,Beijing 100049, China)
Based on molecular phylogenetic evidences, the classification of Gesneriaceae was dramatically changed in recent years. Especially, many genera in the Old Word have been systematically rearranged. However, cytological data of Gesneriaceae have not been re-examined. The cytological data of the Old Word Gesneriads, obtained from WebCyte database, Chromosome Counts Database and related literature, were summarized under the new taxonomic framework of Gesneriaceae. The results showed that the cytological data in Didymocarpoideae was still insufficient. Chromosome number of less than 25% species in this subfamily were reported, and that for many genera have never been reported. Some newly revised genera show uniform somatic chromosome number. However, some genera, such asand, have great variation in chromosome number. Based on these available cytological data, several comments and suggestions on the evolution of chromosome and its influence to species diversification in this family were proposed. It was speculated that both polyploidy and dysploidy of chromosome possibly play important roles to the diversification of some genera. Future works based on new chromosome painting techniques, convinced phylogenetic evidences, and genomics are warranted in studying chromosome evolution of this group.
Gesneriaceae; Didymocarpoideae; Chromosome number; Chromosome evolution
10.11926/jtsb.4082
2019–04–19
2019–07–05
国家自然科学基金委员会与广东省自然科学联合基金项目(U1501211)资助
This work was supported by the Joint Project from the Natural Science Foundation of China-Guangdong Natural Science Foundation (Grant No. U1501211).
杨丽华(1985~ ),男,助理研究员,主要从事苦苣苔科分类及系统发育研究。E-mail: LihuaYang@scbg.ac.cn
E-mail: mingkang@scbg.ac.cn
猜你喜欢
中华养生保健(2022年10期)2022-05-23
小猕猴智力画刊(2021年6期)2021-08-05
特别健康·下半月(2017年10期)2017-10-26
作文大王·低年级(2016年3期)2016-03-11
中国当代医药(2014年23期)2014-09-25
中学理科·综合版(2008年3期)2008-03-07
