
Effect of biochar on grain yield and leaf photosynthetic physiology of soybean cultivars with different phosphorus efficiencies
2019-10-10 06:08ZHUQianKONGLingjianSHANYuziYAOXingdongZHANGHuijunXlEFutiAOXue
Journal of Integrative Agriculture 2019年10期
ZHU Qian, KONG Ling-jian, SHAN Yu-zi, YAO Xing-dong, ZHANG Hui-jun, XlE Fu-ti, AO Xue
Soybean Research Institute, Agronomy College, Shenyang Agricultural University, Shenyang 110866, P.R.China
Abstract This study was conducted with two soybean cultivars, Liaodou 13 (L13, phosphorus (P)-efficient) and Tiefeng 3 (T3,P-inefficient), to investigate the effects of biochar on soybean yield and photosynthetic physiological parameters, at four biochar application rates (0, 1, 5, and 10%, w/w), and two fertilization treatments (0 and 150 kg ha-1). Grain yield, plant biomass, P accumulation, leaf net photosynthetic rate (Pn), chlorophyll index (Chl), nitrogen balance index (NBI), sucrose phosphate synthase (SPS), and sucrose synthase (SS) activities, soluble sugar, sucrose and starch contents, and leaf area duration (LAD) were measured. Biochar had positive effects on Pn, Chl, NBI, SPS, and SS activities, and leaf soluble sugar,sucrose, and starch contents of both genotypes, these effects increased with biochar application rate. L13 benefited more efficiently from biochar than T3 did, as the grain yield of L13 significantly increased by 31.0 and 51.0%, at 5 and 10% biochar,respectively, while that of T3 increased by 40.4 at 10% biochar application rate, as compared with controls. The combined application of biochar and fertilizer boosted the positive effects described, but no difference was found for grain yield in L13 among biochar application rates, while grain yield of T3 continually increased with biochar rate, among which, 1% biochar combined with 150 kg ha-1 fertilizer resulted in T3 yield increment of more than 23%, compared with the application of 150 kg ha-1 fertilizer alone. Altogether, our results indicated that the application of biochar enhanced carbon assimilation in soybean, resulting in increased biomass accumulation and yield. Differences in genotypic responses to biochar highlight the need to consider specific cultivars and biochar rate, when evaluating the potential responses of crops to biochar.
Keywords: biochar, P efficiency, photosynthesis, soybean, yield
1. lntroduction
Inorganic nitrogen (N) and phosphorus (P) fertilizers are essential to ensure adequate plant nutrient supply and maximum crop yield. However, inefficient nutrient recovery from fertilizers occurs globally and prevents crops from achieving desired yields (Galloway et al. 2014). Estimates of overall efficiency of applied fertilizers are up to 50% for N, less than 10% for P, and close to 40% for potassium (K)(Baligar et al. 2001). Lower fertilizer efficiencies are due to significant losses via leaching, run-off, gaseous emission,and fixation of nutrients in the soil (Baligar et al. 2001).These phenomena are affected by soil properties, crop efficiency, climate, and the chemical species of the fertilizer(Hauck 1985; Baligar et al. 1987; Fageria 1992). Fertilizer losses due to plants being unable to successfully absorb them from the soil and use them for biomass accumulation and yield formation can potentially contribute to degradation of soil and water quality, leading to overall environmental degradation (Baligar et al. 2001).
Biochar is produced from pyrolysis of crop residues under limited oxygen conditions and has been widely used as a valuable input for agriculture (Lehmann and Joseph 2015).Biochar has a high surface area and pH, and a strong ability to absorb N, P, and other inorganic ions (Lehmann and Joseph 2009), which reduces the loss of these nutrients from the soil (Lehmann et al. 2011; Hussain et al. 2017).Biochar application to soils alters soil physico-chemical and biological properties, and nutrient cycling (Gul and Whalen 2016); additionally, it increases nutrient bioavailability in soils(e.g., Van Zwieten at al. 2010). The presence of biochar in fertilized soils could foster nutrient availability and enhance the fertilizer use efficiency of plants (Chan and Xu 2009).Therefore, biochar has the potential to improve soil fertility and crop yield (Jeffery et al. 2011; Lehmann and Joseph 2015). Increased crop yield is a generally recognized benefit of biochar application (Hussain et al. 2017); however,it is with drawbacks (Asai et al. 2009; Free et al. 2010;Alburquerque et al. 2014; Ly et al. 2014), and its beneficial effects are highly dependent on the plant species, soil properties, and the efficacy of the biochar itself (type and application rate) (Jeffery et al. 2011; Lehmann et al. 2011).
The genetic and physiological characteristics of plant species also have profound effects on their abilities to absorb and utilize nutrients under various environmental and ecological conditions (Baligar et al. 2001). Nutrient efficient plants are defined as plants that can produce higher yields per unit of nutrient applied/absorbed, compared with other plants grown under similar agroecological conditions(Fageria et al. 2008). Soybean (Glycine max L.) is dependent on P (Sample et al. 1980). Soybean genotypes have been screened for P efficiency in our lab, and P-efficient and-inefficient genotypes have been identified (Li et al. 2004).We previously reported that P-efficient soybeans showed a greater capacity for P absorption, and higher accumulation levels of nutrients than P-inefficient soybeans (Ao et al.2014). Whether biochar application to the soil may improve the productivity of P-inefficient soybeans, or whether the P-efficient soybean genotype may benefit from biochar more efficiently, remains controversial.
Therefore, we studied the responses of the P-efficient and P-inefficient soybean cultivars to biochar at four biochar application rates combined with two fertilizer levels, in a pot experiment. We quantified the effect of biochar application on leaf photosynthetic physiology, and examined the influence of biochar on the grain yield, biomass, and P accumulation in each cultivar. We hypothesized that: (1)biochar application would improve plant growth of both soybean genotypes (P-efficient and P-inefficient); (2)upon biochar application, the increased productivity of the P-efficient genotype would be more efficient and economical than that of the P-inefficient genotype.
2. Materials and methods
2.1. Soil and biochar
Biochar used for the experiment was produced from pyrolysis of rice husks by Liaoning Jinhefu Agricultural Development Co., Ltd., China. The pyrolysis conditions were 450°C for 1 to 2 h, and the detailed method can be found in a China patent (publish number: CN102092709 B). Biochar pH was measured by a Metter-Toledo pH meter in deionized water with a biochar:water ratio of 1:5. Soil samples from the 20-cm top layer in the research site at Shenyang Agricultural University (41°82´N, 123°57´E), China, were used for analyses. After removal of roots and visible plant residues,the soil samples were air-dried. Properties of the soil and biochar used to prepare the pots are shown in Table 1 and were determined before sowing, according to conventional laboratory protocols as described by Lu (2002).
2.2. Experiment description
A pot experiment was carried out under open-field conditions in 2016 at Shenyang Agricultural University, Liaoning Province, China. Two soybean genotypes with different P efficiencies - the P-efficient soybean Liaodou 13 (L13)and the P-inefficient soybean Tiefeng 3 (T3) - were used;both genotypes were previously evaluated for their ability to tolerate P stress (Li et al. 2004). The experiment was laid out in a fully randomized block design with three replicates.The experiment included eight treatments, including biochar application at four rates: 0, 1, 5, or 10% (biochar/dry soil, w/w), and fertilizer (Diammonium phosphate, DAP:N-P-K=18-46-0) application at 0 and 150 kg ha-1.

Table 1 Properties of the soil (0-20 cm) and biochar used in the experiment
The soil was thoroughly mixed with biochar or fertilizer;15 kg of each mixture were packed into plastic pots (30 cm,i.d., by 32 cm height) before sowing. All pots were then watered using drip irrigation to maintain soil moisture content at ~70% of the field water-holding capacity to prevent drought stress from occurring. Each pot contained two plants under the same treatment and was considered as one experimental unit.
2.3. Plant sampling and measurement
During the soybean growing cycle, the middle leaflet of the upper 3rd trifoliate leaf of each plant was used to measure the photosynthesis parameters at the 4th node stage (V4),full anthesis (R2), full pod (R4), and grain-filling stages(R6) (Fehr et al. 1971). Net photosynthetic rate (Pn) was measured with a portable photosynthesis system (LI-6400,LI-COR Inc., Lincoln, NE, USA) equipped with CO2and temperature control modules. The light intensity was set at 1 000 μmol (photon) m-2s-1, which was the light-saturation point for soybean by the measurement of light-response curves. The leaf temperature was kept at 25 to 30°C, relative humidity was 60-65%, CO2concentration of 400 μmol (CO2)mol-1, and air flow of 500 μmol s-1.
The chlorophyll index (Chl) and nitrogen balance index(NBI) was measured by a rapid nondestructive optical device(Dualex 4, Force-A, Orsay, France). After the determination of these traits, the upper 3rd leaves were collected, frozen in liquid nitrogen immediately, and stored at -80°C for evaluation of leaf physiology.
Leaf area per plant was measured at every growth stage by the hole-punch method (Williams and Ruckelshaus 1993)to calculate the leaf area duration (LAD) according to the equation:
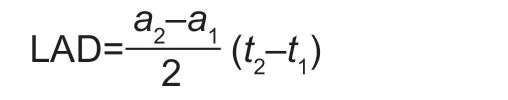
Where, t1and t2are the pervious and later sampling time;a1and a2are the values of the ratio of leaf area to ground area at the sampling times t1and t2, respectively.
At maturity stage (R8), plants were harvested to determine grain yield and plant biomass. The above-ground biomass per plant was measured after oven drying at 85°C for 72 h. The apparent harvest index (AHI) was calculated as grain mass/above-ground plant biomass. Samples were partitioned into individual plant fractions and finely ground and passed through a 2-mm sieve to determine P content by the vanadium-molybdenum yellow colorimetric method,and was used with dry weights to mathematically derive the extent of P accumulation in the plant tissues (Ao et al. 2014).
2.4. Enzyme extraction and assays
Leaves (approximately 3 g fresh mass) were ground in a chilled mortar with 25 mL grinding medium containing 25 mmol L-1Hepes-NaOH (pH 7.5), 5 mmol L-1MaCl2,0.5 mmol L-1ethylene diamine tetraacetic acid (EDTA), 2%PEG-20, 3 mmol L-1dithiothreitol (DTT), and 1% bull serum albumin (BSA). The brei was pelleted by centrifugation at 38 000×g for 10 min. An aliquot of the supernatant was desalted on a 3-mL Sephadex G-50 column (equilibrated with 25 mmol L-1Hepes-NaOH (pH 7.5), 5 mmol L-1MaCl2,and 0.5 mmol L-1EDTA) prior to enzyme assay. Sucrosephosphate synthase (SPS, EC 2.4.1.14) activity was assayed at 37°C, the reaction mixtures (70 μL) contained 50 mmol L-1Hepes-NaOH (pH 7.5), 15 mmol L-1MgCl2,1 mmol L-1EDTA, 5 mmol L-1NaF, 6 mmol L-1uridine diphosphate glucose (UDPG), 4 mmol L-1fructose-6-P,20 mmol L-1glucose-6-P, and 20 μL of crude enzyme.Reaction mixtures were incubated for 30 min at 37°C and incubation was terminated with 70 μL of 5 mol L-1NaOH.Tubes were placed in boiling water for 10 min to destroy any unreacted fructose or fructose-6-P. After cooling, 1 mL of a mixture of 0.14% anthrone in 13.8 mol L-1H2SO4was added and incubated at 40°C in a water bath for 20 min.Colour development of cooled solutions was measured at 620 nm and the SPS activity was calculated. The assay for the sucrose synthase (SS, EC 2.4.1.13) in the synthetic direction was the same, except that fructose replaced fructose-6-P.
2.5. Carbohydrate measurement
Fresh leaf samples (0.1 g) were extracted at 80°C for 1 h in 25 mL of 80% ethanol (v/v) and the supernatant analyzed for soluble sugar and sucrose contents. The residue was boiled for 3 h in 10 mL of 2% HCl (v/v) and the supernatant analyzed for starch content according to the method of Zeng et al. (2014). The soluble sugar was quantified by the anthrone colorimetry method (Xu et al. 2016), while sucrose content was determined through a modified phenol-sulphuric acid method (Yu et al. 2004).
2.6. Statistical analysis
The data were analyzed using fully factorial analysis of variance (ANOVA) and the means were compared using Duncan’s test at P<0.05. A three-way ANOVA was used to test the effects of biochar addition, fertilizer addition,and soybean cultivar. A general response trend across the biochar gradient was also examined using analysis of co-variance (ANCOVA, with biochar rate as the covariate).The relationships between grain yield and photosynthetic physiological parameters were assessed using Pearson’s correlation coefficients. All data were analyzed using SPSS Software version 20.0 (SPSS Inc., Chicago, USA), and Figs.were drawn using SigmaPlot (version 12.5, Systat Software).
3. Results
3.1. Effect of biochar and fertilizer on grain yield and plant biomass
Plant biomass, P accumulation, and grain yield were significantly affected separately by cultivar, fertilizer, and biochar rate of application (Table 2). The number of seeds per plant and AHI were also influenced by biochar application. There was a significant cultivar×fertilizer interaction for L13, but not for T3, which resulted in higher grain yield (30.1%), biomass (43.7%), and P accumulation(15.4%) for L13 compared with T3.
There was also a significant cultivar×biochar interaction for AHI, as 10% biochar application unexpectedly induced a decline in AHI for L13, but not for T3. Nevertheless,biochar application had a positive effect on other measured parameters. Further, significant fertilizer×biochar, and cultivar×fertilizer×biochar interactions for P accumulation and grain yield were detected. P accumulation and grain yield of both soybeans increased with increasing biochar single application rate, but, when combined with fertilizer,there were no differences in the effects of different biochar rates for L13, while grain yield and P accumulation still increased in T3 with increasing biochar application rate(ANCOVA, P<0.05). Thus, biochar was the primary factor responsible for the yield increases experienced by T3 plants.
The grain yield of L13 significantly increased by 31.0 and 51.0%, at 5 and 10% biochar application, respectively, while that of T3 significantly increased by 40.4% at 10% biochar application. Similarly, the biomass of L13 significantly increased by 36.0 and 63.0%, at 5 and 10% biochar,respectively, while that of T3 increased by 44.0% at 10%biochar. Accordingly, significant increase in P accumulation after single biochar application reached 60.1% for L13,and up to 43.2% for T3 at the highest biochar application rate. When biochar was co-applied with fertilizer, meangrain yield and P accumulation in T3 increased by 53.3 and 45.4%, respectively, compared to single fertilizer application.Interestingly, the addition of 10% biochar produced a higher grain yield, biomass, and P accumulation in T3 than a single fertilizer application. However, the combined application of fertilizer and the highest rate of biochar was consistently associated with the largest increase in plant growth.

Table 2 Effects of biochar and fertilizer on yields and yield-related parameters of two soybean cultivars with different phosphorus efficiencies1)
3.2. Effect of biochar and fertilizer on leaf photosynthetic traits
Net photosynthetic rate (Pn)Leaf Pnwas affected by both cultivar and biochar applications, and by fertilizer at the R4 and R6 stages. L13 showed significantly higher Pnthan T3 did at V4 and R6 growth stages. At R6 stage, L13 retained a high level of Pn, while T3 showed a sharp decline in Pn,compared to the R4 stage (Fig. 1). Significant interactions between three or each two factors were observed at the V4 stage, but not at any other stages. The positive effect of biochar on Pnfor L3 was the highest at the R4 stage, as it significantly increased by 15.2, 13.4, and 22.6% at 1, 5,and 10% biochar, respectively. Meanwhile, the Pnof T3 significantly increased by 21.7 at 1% biochar application,compared to the control at the R6 stage.
Leaf chlorophyll index (Chl)The Chl was affected by cultivar, biochar, and fertilizer application. L13 showed significantly higher Chl than T3 at later growth stages (R4 and R6). Fertilizer application resulted in an increment in leaf Chl at the V4 and R6 stages, while biochar produced significantly positive effects on Chl at all stages under study (Fig. 2). The leaf Chl of both soybean cultivars under study increased with increasing biochar rate at the V4 stage, and the largest increases were 17 and 41%for L13 and T3, respectively, at 10% biochar application,compared to 0% biochar (control). There was a significant cultivar×biochar×fertilizer interaction for leaf Chl at the R6 stage, with a peak in L13 under the co-application of the highest biochar and fertilizer rates. Additionally, the Chl of L13, but not T3, significantly increased with biochar and fertilizer applications, but no increase in Chl value was observed when biochar was applied in combination with fertilizer, compared to fertilizer application alone (Fig. 2).
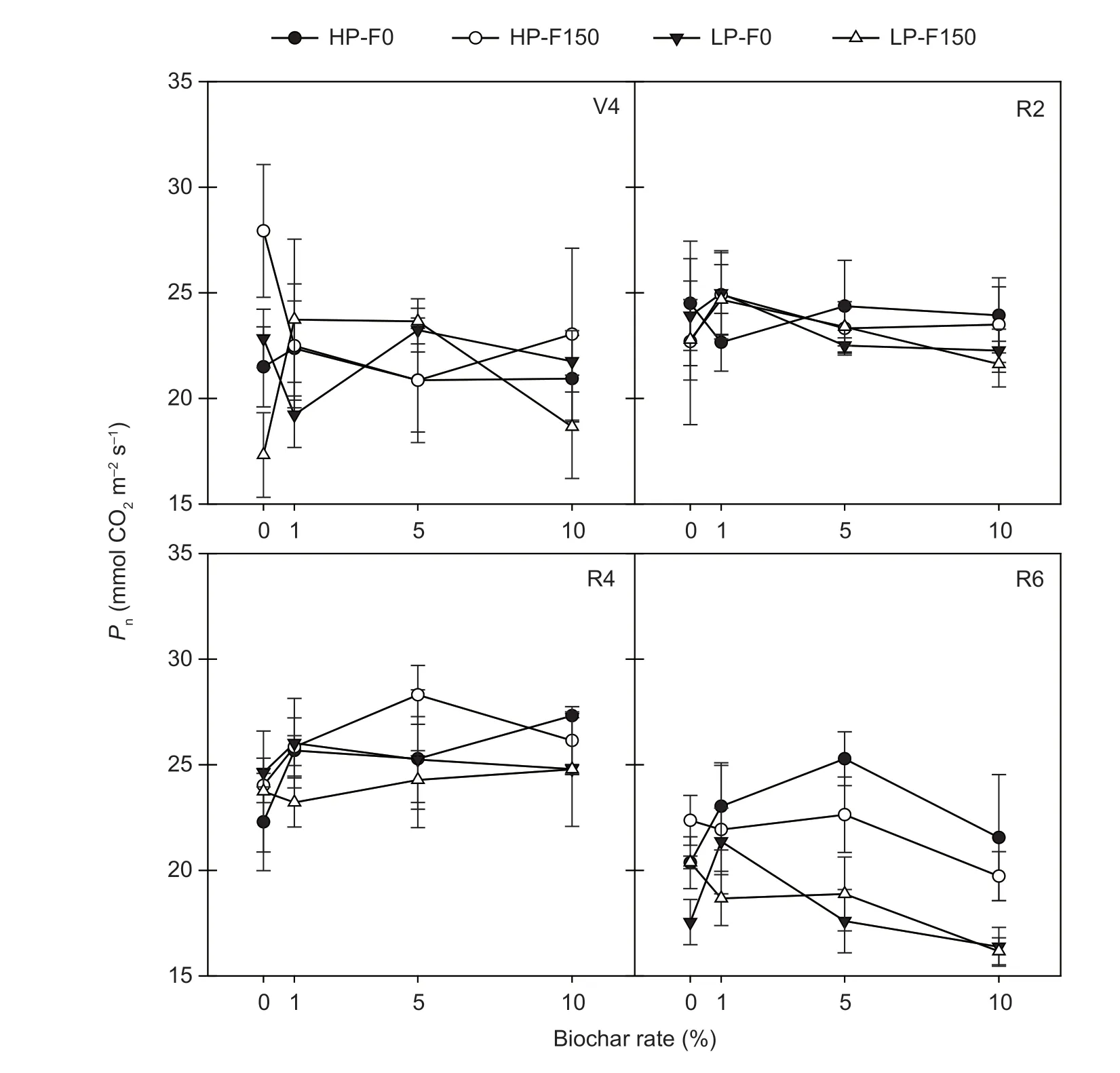
Fig. 1 Effect of biochar and fertilization application on net photosynthetic rate (Pn) of soybeans with different P efficiencies at different growth stages (mean value±SD, n=3). HP, P-efficient soybean Liaodou 13 (L13); LP, P-inefficient soybean Tiefeng 13 (T3); F0,no fertilizer; F150, 150 kg ha-1 diammonium phosphate (DAP) application. V4, R2, R4, and R6, 4th node stage, full anthesis, full pod, and grain-filling stages, respectively.
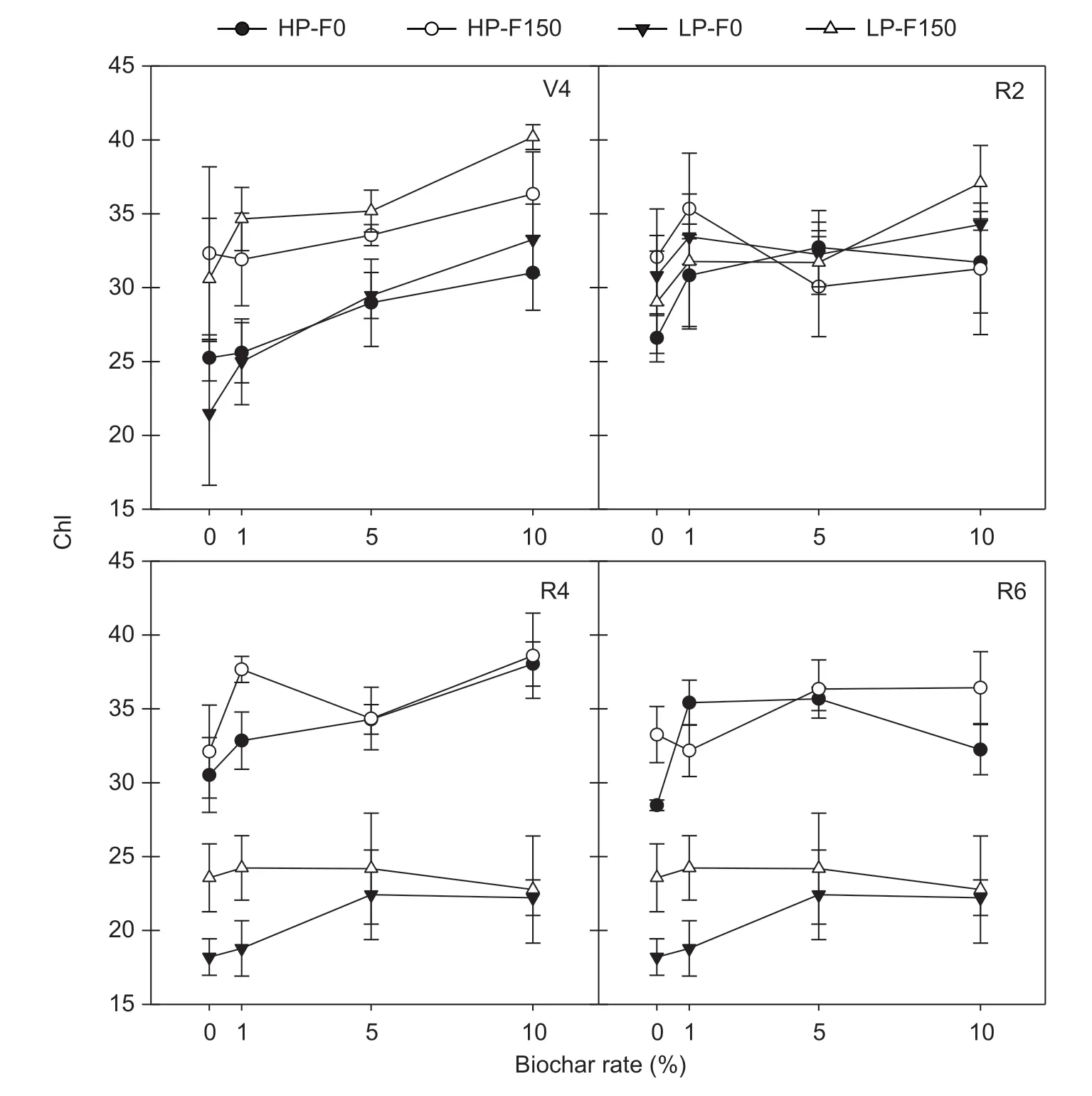
Fig. 2 Effect of biochar and fertilization on leaf chlorophyll index (Chl) of soybeans with different P efficiencies at different growth stages (mean value±SD, n=3). HP, P-efficient soybean Liaodou 13 (L13); LP, P-inefficient soybean Tiefeng 3 (T3); F0, no fertilizer;F150, 150 kg ha-1 diammonium phosphate (DAP) application. V4, R2, R4, and R6, 4th node stage, full anthesis, full pod, and grain-filling stages, respectively.
Nitrogen balance index (NBl)Significant differences were observed for NBI between the two soybean cultivars, as L13 showed much higher NBI than T3 at most stages measured,except V4 (Fig. 3). Fertilizer application resulted in a significant increment of NBI at the V4 stage, while biochar showed positive effects on NBI at all stages. There was a significant cultivar×fertilizer interaction at the V4 stage, as L13 displayed higher NBI than T3 did without fertilizer, but lower with fertilizer application (Fig. 3). The highest value of NBI was observed at 10% biochar application, irrespective of fertilizer, at V4 and R2 stages, which was significantly higher than the corresponding values for 0% biochar controls. The increment in NBI decreased with increase in biochar rates at later growth stages.
Leaf area duration (LAD)LAD was affected by cultivar,fertilizer, and biochar. L13 displayed obviously higher LAD than T3 in growth periods of V4-R2 and R2-R4 (Fig. 4).Both biochar and fertilizer applications had positive effects on LAD of the two soybean cultivars throughout all growth stages measured. Moreover, there was a significant cultivar×biochar×fertilizer interaction for LAD in all three growth periods, and the peak was observed in L13 at the combined application of 10% biochar and fertilizer at V4-R2 and R2-R4 growth stages, and under combined application of 5% biochar and fertilizer at R4-R6 stages in T3.

Fig. 3 Effects of biochar and fertilization on leaf nitrogen balance index (NBI) of soybeans with different P efficiencies at different growth stages (mean value±SD, n=3). HP, P-efficient soybean Liaodou 13 (L13); LP, P-inefficient soybean Tiefeng 3 (T3); F0, no fertilizer; F150,150 kg ha-1 diammonium phosphate (DAP) application. V4, R2, R4, and R6, 4th node stage, full anthesis, full pod, and grain-filling stages, respectively.

Fig. 4 Effect of biochar and fertilization on leaf area duration (LAD) of soybeans with different P efficiencies at different growth periods (mean value±SD, n=3). HP, P-efficient soybean Liaodou 13 (L13); LP, P-inefficient soybean Tiefeng 3 (T3); F0, no fertilizer;F150, 150 kg ha-1 diammonium phosphate (DAP) application. V4-R2, R2-R4, and R4-R6, soybean growth periods of branching to blooming, blooming to podding, and podding to grain-filling, respectively.
LAD significantly increased with 5% biochar in the growth period of V4-R2, and with 10% biochar in the R2-R4 stages.During R4-R6 stages, LAD of L13 significantly increased at all rates of biochar application, while LAD of T3 significantly increased with 10% single biochar application, compared with each 0% biochar treatment. When biochar and fertilizer were co-applied, the LAD of T3, but not L13, significantly increased with 5 and 10% biochar, compared to single fertilizer application, in the growth periods of R2-R4 and R4-R6 stages.
3.3. Effect of biochar and fertilizer on leaf enzymatic activity and carbohydrate content
Sucrose phosphate synthase (SPS) and sucrose synthase (SS)Soybean leaf SPS and SS activities were affected by cultivar, fertilizer, and biochar throughout all the measured growth stages (Table 3 and Appendix A). The application of biochar resulted in an increase in SPS and SS activities, and there was a significant cultivar×biochar interaction for both enzyme activities. In L13, the activity of SPS was significantly higher at all biochar addition rates than at the 0% biochar treatment, except the 1% biochar rate at the R6 stage. Meanwhile, the SPS and SS activities in T3 were increased only at the highest (10%) biochar application rate at most stages, except V4. In addition, there were significant fertilizer×biochar and cultivar×fertilizer×biochar interactions for SS activity at the V4 stage, as the 10%single biochar application significantly reduced the SS activity in T3.
Total soluble sugar, sucrose, and starch contentsThe total soluble sugar content in soybean leaves was significantly affected by cultivar (Table 4), with T3 showing much higher total soluble sugar content than L13 at the V4-R4 growth stages, although it was lower at the R6 stage (Appendix B). Biochar had significant effects on leaf soluble sugar content for both soybean cultivars in the reproductive stages (R2-R6 stages). There were significant fertilizer×biochar, and cultivar×fertilizer×biochar interactions at the R2 stage, as total soluble sugar content increased at 5 and 10% single biochar applications in both soybean cultivars compared with controls; and increased in L13 with co-application of biochar and fertilizer compared with single fertilizer treatments. Conversely, total soluble sugar content significantly decreased in T3 when biochar was applied in combination with fertilizer, compared with corresponding single biochar rate. At other stages, no significant interactions between cultivar, biochar, and fertilizer were found, as total soluble sugar content increased with increasing biochar application rate. The only difference was that the total soluble sugar content in L13 significantly increased at all biochar rates, while it increased in T3 only at R4 stage at 10% biochar application.
Leaf sucrose content was much higher in T3 than in L13, while starch content was higher in T3 than in L13 at R2 stage, but much lower than in L13 at the R6 stage. The contents of sucrose and starch were significantly affectedby biochar application throughout the reproductive stages(R2-R6), and increased with the addition of fertilizer to some extent (Appendix B). Leaf sucrose and starch contents increased as biochar application rate increased, and displayed significant increments at 10% biochar application rate, as compared to controls.

Table 3 Effects of biochar, fertilizer, and cultivar on the activity of sucrose phosphate synthase (SPS) and sucrose synthase (SS)in soybean leaves

Table 4 Effect of biochar, fertilizer, and cultivar on carbohydrate content in soybean leaves1)
3.4. Relationships between yield and photosynthetic traits
Soybean leaf photosynthetic traits significantly influenced grain yield. Pearson’s correlation analysis (Table 5)showed that yields of both soybean cultivars were positively correlated with leaf photosynthetic traits, mainly at the early growth stages (Chl, NBI, and SPS activities at V4 stage),and at R6 stage (SPS and SS activities, sucrose content).In addition, yield and LAD were positively correlated in the R2-R4 stages for both soybean cultivars studied.
4. Discussion
4.1. Effects of biochar, and interaction of biochar and fertilizer on soybean yield
In general, the application of inorganic fertilizer improves crop yield. However, fertilizer use efficiency depends on the crop type. In this study, biomass and grain yield of L13 significantly increased by 49.6 and 47.0% with fertilizer application, respectively, while only a 6.3% increase in biomass and a 4.1% increase in grain yield were observed in T3. These results were mainly caused by the different nutrient efficiencies of each of the two soybean cultivars.L13 demonstrated a greater ability to absorb nutrients from soils by a larger root length and greater root vitality than shown by T3 (Ao et al. 2008). Our previous study indicated that regardless of P level (normal, sufficient or deficient), whole plant or specific organ P accumulation by the P-efficient soybean plant were all higher than those by the P-inefficient plant; this observation confirmed a greater capacity for P nutrient absorption in P-efficient soybean genotypes (Ao et al. 2014). Accordingly, in this study,P accumulation per plant in L13 significantly increased by 20.9% with fertilizer application relative to unfertilized controls, but no difference in P accumulation was found between fertilizer levels for T3. These differences enhanced the difference in plant harvest between L13 and T3, resulting in much higher grain yield, biomass, and P accumulation in L13 than in T3, upon fertilizer application.
Nevertheless, a positive effect of biochar was observed on plant growth in both soybean cultivars, which verified our first hypothesis. The positive effect of rice husk-derived biochar on yield was also found in rice (Haefele et al.2011) and maize (Mekuria et al. 2014). Previous studies reported increases in crop yield due to increased P uptake after the application of biochar (Major et al. 2010; Uzoma et al. 2011; Hussain et al. 2017). Furthermore, higher yield always resulted from high plant biomass induced by biochar application (Steiner et al. 2007). In this study, we found that the increase in grain yield was a result of an increase in plant biomass and P accumulation, as well as in seeds per plant, which together contributed to the increase in grain yield. An increase in P accumulation has several impacts on photosynthesis, including affecting energy transfer across the thylakoid membrane (Foyer and Spencer 1986),influencing several key enzymes involved in the Calvin cycle(Jacob and Lawlor 1992), and affecting electron transfer(Preiss 1984) and carbohydrate metabolism (Zhang et al.2014).
Biochar application rate affected grain yield in contrasting soybean cultivars for different P efficiencies. Higher biochar rate induced more plant growth, which was consistent with the result of Gul and Whalen (2016). The difference inresponses to biochar rate between the two soybean cultivars tested here was that the P-efficient soybean showed a significant increase in grain yield at a relatively lower biochar application rate (31.3% at 5% biochar) than the P-inefficient soybean (40.4% at 10% biochar). Therefore, a much higher yield-increment of L13 was achieved at the same biochar rate than that of T3. This result was consistent with the response of biomass and P accumulation to biochar rates.Further, it suggested that the P-efficient soybean benefited from biochar application more efficiently and economically than the P-inefficient one.

Table 5 Pearson’s correlation coefficients between grain yield and photosynthetic parameters in soybean cultivars
Soybean grain yield was affected by the interaction of biochar and fertilizer. The influence of biochar application on soybean growth was more pronounced when applied in combination with fertilizer. The combined application of inorganic fertilizer and biochar resulted in sustained crop yield, which was consistent with Brown et al. (2011). Similar results were reported for maize (Yamato et al. 2006), rice,and sorghum (Chrishtoph et al. 2007; Steiner et al. 2007).However, the increment in grain yield observed for L13 decreased as biochar rate increased, when combined with fertilizer. This was explained by the decrease in AHI at high biochar application rates combined with fertilizer. In contrast, grain yield, as well as biomass and P accumulation in T3 increased with increasing biochar rate irrespective of fertilization. This was all due to the inefficacy of single fertilizer application. Furthermore, we found that the reaction of a single application of 10% biochar, as well as 1% biochar with 150 kg ha-1fertilizer, resulted in T3 yield increases greater than 23%, compared with that at the application of 150 kg ha-1fertilizer only. This indicated that biochar applications could remedy with the low use of fertilizer by the P-inefficient soybean cultivar, thereby increasing plant productivity.
4.2. Effects of biochar and interaction of biochar and fertilizer on soybean photosynthetic physiology and biochemistry
Photosynthesis is an important physiological process to crop productivity, contributing to 90-95% of plant yield(Du et al. 1999). Previous studies indicated that biochar increased the photosynthetic rate and water use efficiency of potato and grape (Akhtar et al. 2014; Baronti et al. 2014).Xu et al. (2015) further showed that biochar increased leaf photosynthetic rate and the maximum electron transport rate in peanut. In this study, Pnwas also increased in soybeans.Biochar application resulted in enhancement of soybean leaf Chl and NBI across growth stages. Chl reflects the relative content of chlorophyll in leaves, which is correlated with N content, and NBI represents N level (Padilla et al.2014). Photosynthesis was positively correlated with leaf N content (Wright et al. 2004), and the increase in Chl and NBI directly accelerated photosynthetic capacity, thus increasing Pn. LAD also increased with biochar application,enabling plants to intercept more light during plant growth(Vivian et al. 2013). In general, higher LAD promotes dry matter accumulation and higher yield (Kumudini et al.2001). Further, an increase in LAD during the middle and late growth stages of soybean also has a significant impact on yield (Chu 1988). In this study, soybean yield and LAD were positively correlated, and the addition of biochar increased LAD during the reproductive stages for both soybean cultivars under study, which promoted leaf carbon assimilation (Yu et al. 2004), thereby increasing plant biomass accumulation and grain yield (Yin et al. 2012;Bruun et al. 2014).
Furthermore, we observed genotypic variation in photosynthetic traits, with L13 showing much higher Pn,Chl, NBI, and LAD values than T3, especially at the late growth stages. Specifically, during R4-R6, L13 maintained increased Chl and LAD with biochar or fertilizer applications,while T3 did not. In the case of the combined application of biochar and fertilizer, there were no differences in the effect of biochar application rates on Chl or LAD for L13, but there were in the case of T3, thus demonstrating that biochar acted as a primary factor for stimulating the growth of T3 plants.
Biochar and fertilizer applications resulted in increased SPS and SS activities and carbohydrate content in functional leaves during the reproductive stages of both P efficient soybeans, which might promote a feedback type of regulation of sugar signals in photosynthesis (Paul and Foyer 2001).SPS and SS are both involved in sucrose synthesis and their combined effect under biochar application stimulated high production of sucrose, which then increased total soluble sugar content. Since sucrose is the main product of photosynthesis, we speculate that biochar enhanced carbon assimilation during photosynthesis may provide more carbohydrates for plant metabolism, which would increase yield (Schnyder 1993). Soluble sugars, which are the primary products of photosynthesis in higher plants, are the building blocks of macromolecules that control plant growth and development (Gibson 2000). The enrichment in soluble sugars induced by biochar and fertilizer in soybean functional leaves, may act as a trigger that controls osmotic pressure at the cellular level (Cha-um et al. 2009). Moreover, starch is the main source of carbohydrate (Halford et al. 2011) and the end-product of a metabolic pathway that is linked to the Calvin-Besson cycle via plastid phosphoglucose isomerase(Bahaji et al. 2014), which is closely linked with P content in plants (Zhang et al. 2014). The over-accumulation of starch is accompanied by enhanced up-regulation of SS and other enzymes involved in sugar metabolism (Bahaji et al. 2014). Therefore, the increase in starch in soybean leaves upon biochar application would benefit the storage of photosynthetic products, ultimately contributing to grain yield (Bahaji et al. 2014).
5. Conclusion
Biochar had positive effects on soybean growth, irrespective of cultivar differences in P efficiency. The positive effects of biochar on leaf photosynthetic traits, including the increase in leaf Chl, NBI, and LAD, and the enhanced activity of SPS and SS, altogether increased leaf carbohydrate content,and P accumulation, ultimately resulting in increased yield and biomass accumulation. A higher rate of biochar application promoted higher soybean productivity, and the P-efficient cultivar benefited from biochar application more than the P-inefficient cultivar. The combined application of inorganic fertilizer and biochar maintained crop yield, and a synergistic effect of biochar and fertilizer improved crop yield in the P-inefficient soybean. Application of biochar alone at an optimal rate or in combination with inorganic fertilizer could improve photosynthesis and plant growth for better grain yield in soybean cultivars with different P efficiencies,thereby resulting in much less environmental pollution.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31271643), the Program for Liaoning Excellent Talents in University, China (LJQ2015097), the Natural Science Foundation of Liaoning Province, China(20170540809), and the National Key R&D Program of China (2017YFD0101306-04).
Appendicesassociated with this paper can be available on http://www.ChinaAgriSci.com/V2/En/appendix.htm
 Journal of Integrative Agriculture2019年10期
Journal of Integrative Agriculture2019年10期
- Journal of Integrative Agriculture的其它文章
- Application of virus-induced gene silencing for identification of FHB resistant genes
- Dynamic changes of root proteome reveal diverse responsive proteins in maize subjected to cadmium stress
- Strategies to enhance cottonseed oil contents and reshape fatty acid profile employing different breeding and genetic engineering approaches
- Maize/peanut intercropping increases photosynthetic characteristics,13C-photosynthate distribution, and grain yield of summer maize
- Rhizosphere soil bacterial community composition in soybean genotypes and feedback to soil P availability
- lnheritance of steroidal glycoalkaloids in potato tuber flesh