
三江源区不同海拔高寒草原土壤养分及化学计量特征
2019-09-25 11:03姜哲浩陈建忠张德罡陈建纲柳小妮
草地学报 2019年4期
姜哲浩, 周 泽, 陈建忠, 张德罡, 陈建纲, 柳小妮
(甘肃农业大学草业学院, 草业生态系统教育部重点实验室, 甘肃省草业工程实验室,中-美草地畜牧业可持续研究中心, 甘肃 兰州 730070)
土壤是影响植物群落物种组成和群落动态的重要因子,土壤养分往往制约着生态系统的演替过程和对环境变化的响应方式[1]。生态化学计量学是研究生态系统生态过程中不同元素之间的平衡关系、植物与土壤相互作用与碳、氮、磷循环的新思路[2]。碳(carbon,C)是植物各种生理生化过程的底物与能量来源,也是衡量生态系统有机物质储量的主要指标[3];氮(nitrogen,N)、磷(phosphorus,P)是植物生长的主要限制因子,并且这种限制作用可以通过植物的氮、磷含量及N/P反映出来[4]。土壤是植物生长的基质,为植物体生长提供养分。全球陆地生态系统大多受到N限制,尤其是土壤C∶N∶P计量学特征对陆地生态系统中碳固定过程具有极强的调控作用[5],因此对土壤生态化学计量特征的研究不可忽视。目前,随着我国学者对该学科关注度的提高,土壤生态化学计量特征的研究也逐渐增多。王健林等[6]研究了青藏高原高寒草原生态系统土壤C/N的分布特征,发现不同草地类型和不同自然地带土壤碳氮比差异显著,而表土层与底土层土壤碳氮比差异也显著;刘颖等[7]研究了川西高寒灌丛草地不同海拔梯度土壤特征,发现磷和海拔高度显著相关,高海拔地区显著高于低海拔地区;李丹维等[8]对太白山不同海拔的土壤进行研究发现海拔、温度和水分显著影响土壤碳、氮、磷化学计量特征。
“三江源”素有“中华水塔”之称,对中国的生态状况及国民经济发展起着重要作用。该区域是研究世界上高海拔生物多样性最理想的地区之一。刘育红等[9]对三江源区高寒草甸植物功能进行了研究,尹亚丽等[10]对该区域高寒草甸土壤微生物进行了研究,魏卫东等[11]通过对退化草地群落特征和土壤因子数据进行冗余分析,探讨了高寒草甸土壤因子与草地退化之间的关系,吴航[12]研究表明该地区植物中碳氮磷含量与海拔高度之间存在相关性。但鲜有对该区域土壤化学计量特征的空间分布格局的研究。三江源地区海拔相对高差较大,气候和植被类型垂直变化分异明显,揭示该区域土壤各组分间的化学计量特征及其相互关系和空间变异,有助于深入认识碳、氮、磷生物地球化学循环的特征。
因此,本文以三江源区不同海拔高度(2 512~4 493 m)高寒草原土壤为研究对象,比较了不同海拔梯度之间土壤养分含量的差异,同时以生态化学计量学方法探讨了三江源区土壤碳、氮、磷养分垂直分布特征,分析了海拔对土壤养分及生态化学计量特征的影响,为三江源区土壤养分供应状况及限制情况提供参考,也可为三江源区生态系统的保护提供重要的理论依据。
1 材料与方法
1.1 研究区概况
研究区位于青海省南部(31°39′~36°19′ N,89°45′~102°23′ E),海拔2 500~4 800 m,属高原大陆性气候,年日照时数2 550~2 760 h,年均温—5.6~—3.8℃,年平均降水262.2~772.8 mm,6-9月降水量占全年降水量的75%,冷季漫长,无明显暖季。草地类型为高寒草原,草层高度25~70 cm,草层盖度56%~95%,土壤类型为高山草原土[13],土壤质地为砂土[14]。样地详情见表1。
1.2 野外调查与取样
2018年8月在研究区进行植被调查和土壤样品采集,在海拔2 500~4 500 m范围以约500 m高差为间隔分别选取5个样地(表1)。每个样地设置3个200 m×200 m的采样区,每个采样区随机设置5个50 cm×50 cm的样方,记录植物种类、草层高度(Grass height,GH)及盖度(Coverage),然后将样方内的植物齐地面刈割,放入样袋,经105℃杀青后,在60℃下烘干至恒重,计算草地植被地上生物量(Above ground biomass,AGB)。去除土层上的枯落物后,采用五点法采集土壤样品,使用土钻采集0~10,10~20,20~30 cm土层的样品。样品经过混合密封后带回实验室,去除土壤动物和植物残体,一份土样测定土壤含水量,另一份自然风干后磨碎,过0.15及2 mm筛用于指标测定。同时,在0~10,10~20,20~30 cm土层采用环刀法(100 cm3)取样,测定土壤容重(Soil bulk density,SBD)。
1.3 样品的处理与测定
土壤含水量(Soil water content,SWC):105℃烘干法;土壤pH值:电位法(水土比为2.5∶1);土壤有机碳(Soil organic carbon,SOC):重铬酸钾氧化-外加热法;土壤全氮(Total nitrogen,TN):凯氏定氮法;土壤全磷(Total phosphorus,TP):酸熔-钼锑抗比色法[15]。
1.4 数据分析
土壤碳氮比(C/N)、碳磷比(C/P)和氮磷比(N/P)均采用质量比。SPSS21.0软件对数据进行One-Way ANOVA统计分析和Duncan极差法进行差异显著性比较,采用Pearson法分析土壤各指标间的相关性,采用QIIME(Version1.7.0)进行不同样本植物多样性指数分析,采用SigmaPlot14.0作图,采用Canoco5.0软件对各海拔梯度间土壤理化和生态化学计量比指标进行主成分分析(Principle Component Analysis,PCA)。
2 结果与分析
2.1 植被变化特征
由表2可知,草地生物量在各海拔梯度间无显著差异,但在高海拔处最高;草层高度在海拔3 005,3 490和3 977 m之间无显著差异,而低海拔2 512 m处,草层高度显著较高(P<0.05);植被盖度在海拔2 512 m处最低,显著低于其他4个海拔梯度(P<0.05);植被Shannon指数在不同海拔间差异显著(P<0.05),并随海拔升高而增加。

表2 不同海拔梯度高寒草原植被特征Table 2 Vegetation characteristics of alpine steppe under different altitudes
注:同列不同小写字母表示不同海拔梯度之间差异显著性(P<0.05)
Note:Different lowercase letters in the same column indicate significant differences between different altitude gradients at the 0.05 level
2.2 土壤pH、含水量和容重变化特征
在不同海拔草地土壤中,随着土层深度的增加,土壤pH和含水量降低,容重则升高(图1),其中,土层深度对pH值的影响显著(P<0.05)。海拔差异对pH值和土壤含水量无明显影响,但对容重有显著影响(P<0.05),表现为随着海拔升高,容重呈先上升后下降的趋势,在3 490 m处最高(1.41 g·cm-3)。
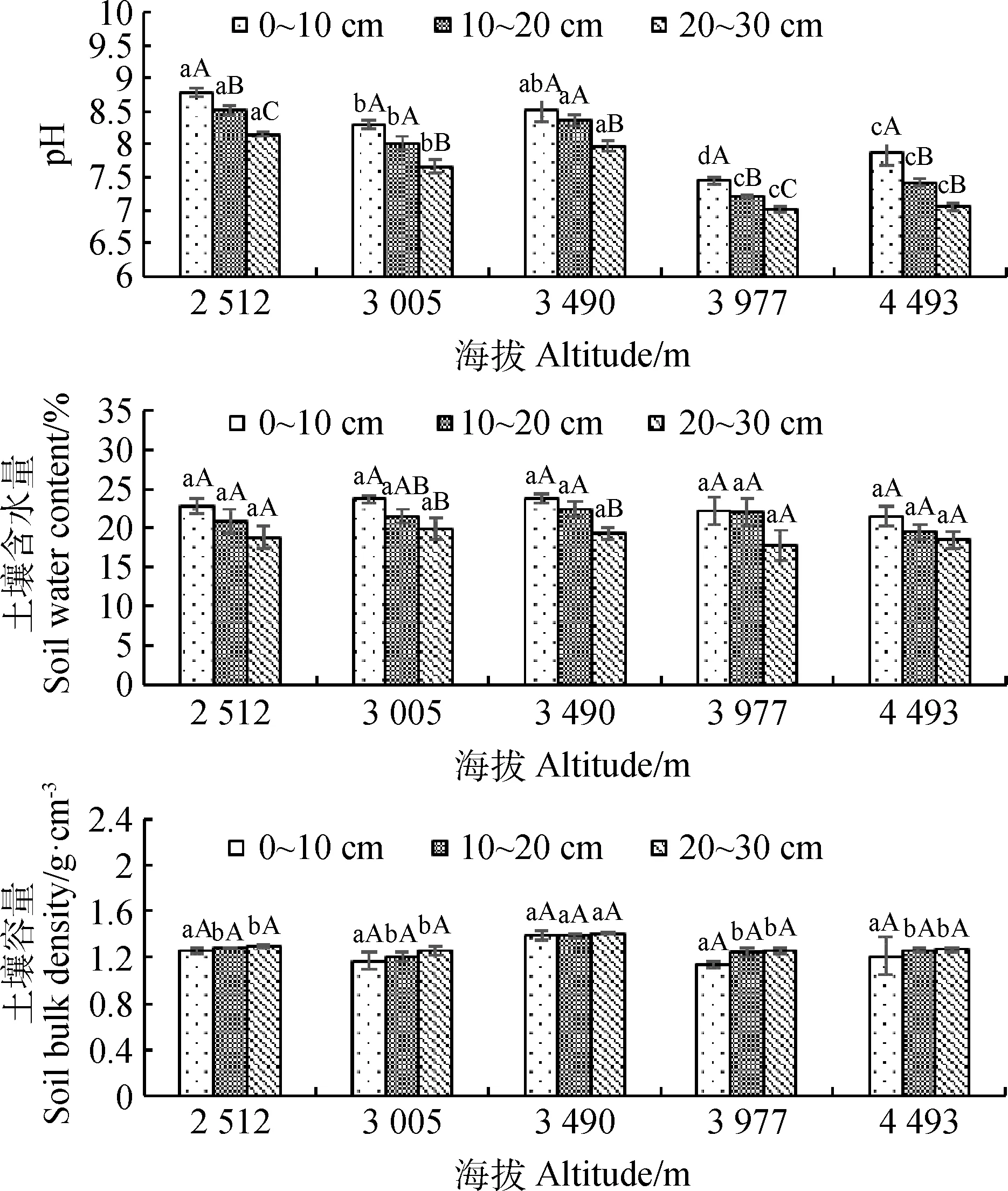
图1 不同海拔高寒草原土壤pH、含水量和容重变化特征Fig.1 Characteristics of soil pH,water content and bulk density of alpine steppe under different altitudes注:误差线为标准误。不同小写字母表示相同土层深度不同海拔梯度差异显著性(P<0.05),不同大写字母表示相同海拔梯度不同土层深度之间差异显著性(P<0.05),下同Note:Error bar is standard error. Different lowercase letters indicate significant differences in altitude gradients between different soil depths at the 0.05 level,Different capital letters indicate significant differences in soil depth between different soil gradients at the 0.05 level,the same as below
2.3 土壤有机碳、全氮、全磷含量变化特征
由表3可以看出,随着海拔的升高,0~30 cm土层中磷含量逐渐升高,表现为4 493>3 977>3 490>3 005>2 512 m。碳和氮含量在海拔2 512~3 490 m呈上升趋势,在3 490 m处达到最高后又逐渐下降,各海拔之间差异显著(P<0.05)。随着海拔的升高,碳和氮含量呈先上升后下降趋势,磷含量呈上升趋势。碳和氮含量在各海拔梯度都呈现出随土层增加而降低的规律,且各土层差异性显著(P<0.05)。碳和氮在表层的聚集现象较为明显,海拔2 512,3 005,3 490,3 977和4 493 m的土壤在0~10 cm土层占整个土壤剖面有机碳的百分比分别为47.98%,42.09%,37.09%,39.74%和41.53%,氮百分比分别为49.75%,44.42%,49.39%,44.98%和50.80%。
2.4 土壤有机碳、全氮、全磷的生态化学计量特征
由表4可知,随着海拔的升高,各海拔间0~30 cm土层的土壤C/N,C/P,N/P均呈现出先增加后减少的趋势,这一变化趋势和土壤碳、氮含量的变化趋势一致。
对不同海拔梯度土壤碳、氮、磷生态化学计量比C/N,C/P,N/P进行分析(表4),其中,土壤C/N变化范围为11.16~35.22。在海拔4 493 m的0~10 cm土层中最低,在3 490 m的20~30 cm土层中最高,增加了3.16倍。随土层的加深,土壤C/N呈增加趋势。
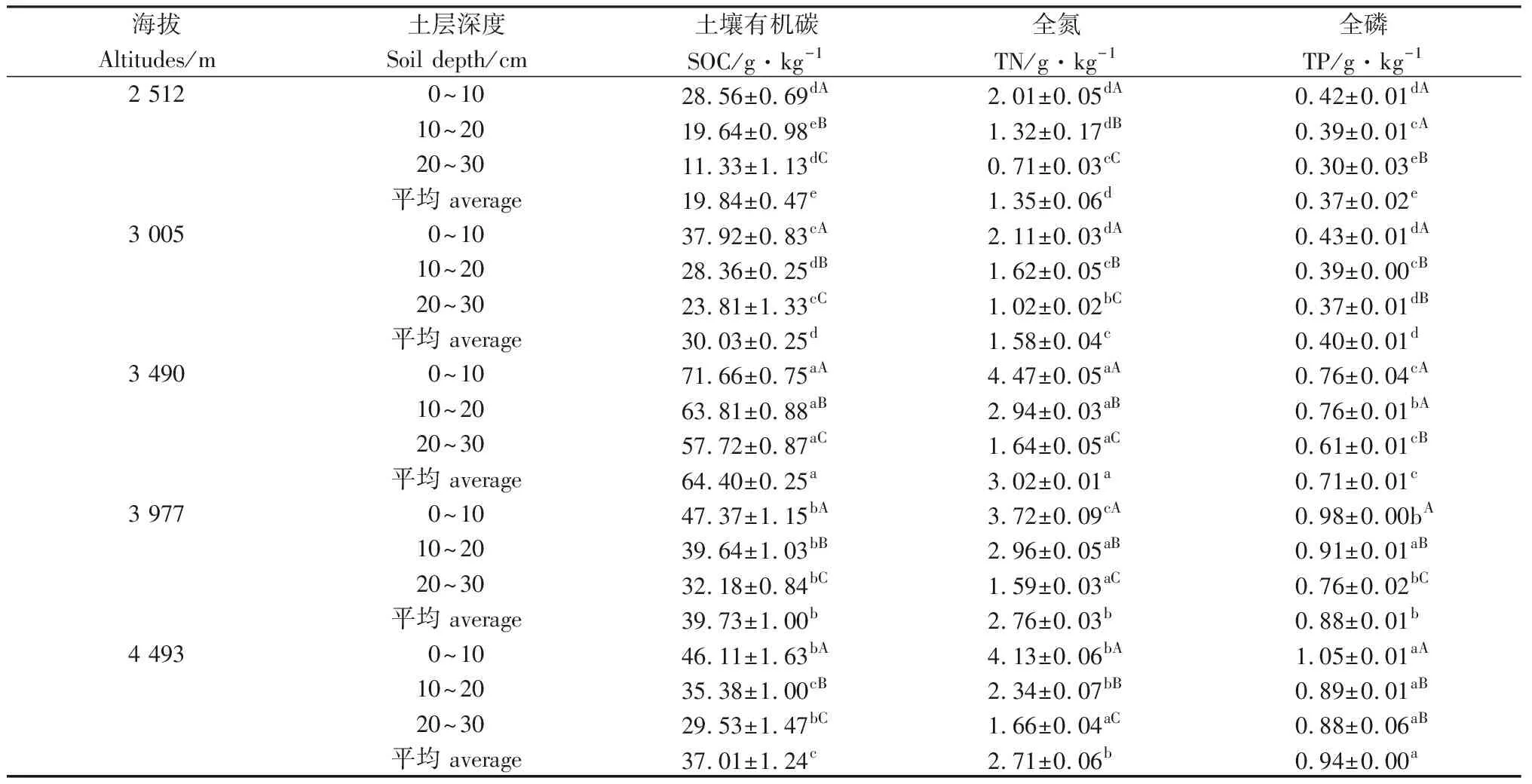
表3 不同土层深度土壤有机碳、全氮、全磷含量Table 3 Content of organic carbon,total nitrogen and total phosphorus in soil with different depths
土壤C/P范围为33.65~94.48。土壤C/P随海拔梯度的增加变化趋势与土壤C/N类似,在3 490 m的20~30 cm土层中最高,在4 493 m的20~30 cm土层中最低。随土层的加深,土壤C/P呈逐渐降低趋势,这与土壤C/N的变化趋势相反,但在3 490 m呈先下降后上升趋势。
土壤N/P的范围为1.89~5.88。N/P随海拔梯度增加先升后降,随土层深度的加深,大致呈下降趋势。在海拔4 493 m的20~30 cm处达到最低值。
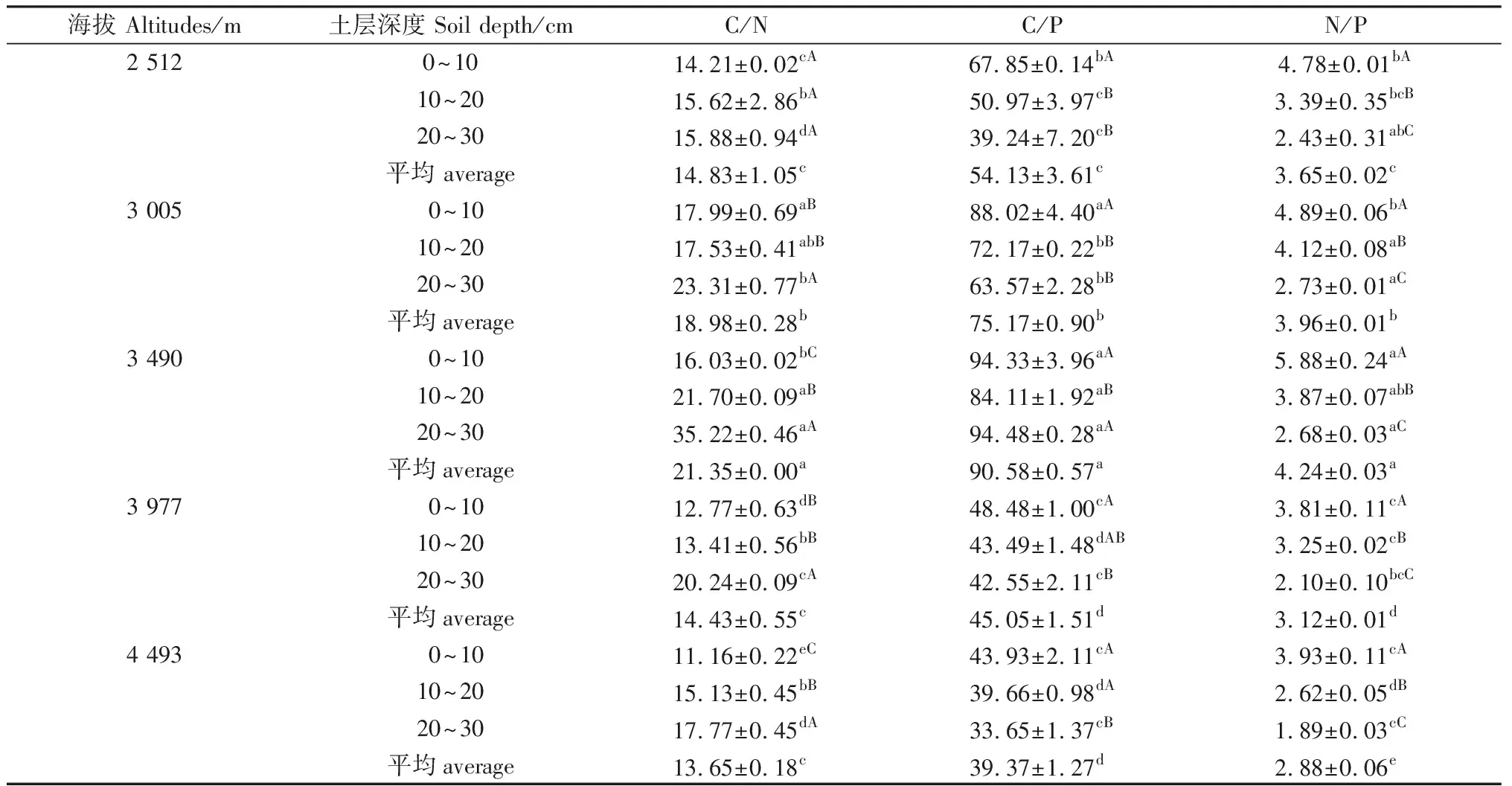
表4 不同土层深度土壤化学计量特征Table 4 Ratios of soil C/N,C/P and N/P in different depths
2.5 土壤理化特性与生态化学计量比的PCA分析
对三江源不同海拔梯度高寒草原土壤理化特性及生态化学计量比之间进行PCA分析,并结合Pearson相关系数分析土壤环境因子间的相互关系。由表5、图2可知,第1,2主成分轴分别贡献了65.2%和31.4%的信息量,累计贡献率达到了96.6%,可以反映出土壤环境的基本信息。从土壤养分含量与海拔之间的关系来看,氮、磷与海拔呈现极显著正相关(P<0.01),而碳与海拔之间的相关性并不明显。从土壤生态化学计量特征与海拔之间的关系来看,N/P和海拔之间极显著负相关(P<0.01)。养分含量之间,氮和碳、磷之间极显著正相关(P<0.01)。对养分含量和化学计量比之间的相关性分析可知,氮与化学计量比之间的相关性不显著;碳与C/N,C/P之间显著正相关(P<0.05),磷与N/P之间显著负相关(P<0.05)。

表5 土壤化学计量特征与海拔之间的相关性分析Table 5 The correlation analysis between soil stoichiometry characteristic and altitude
注:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
Note:*means significant correlation at the 0.05 level;**means extremely significant correlation at the 0.01 level
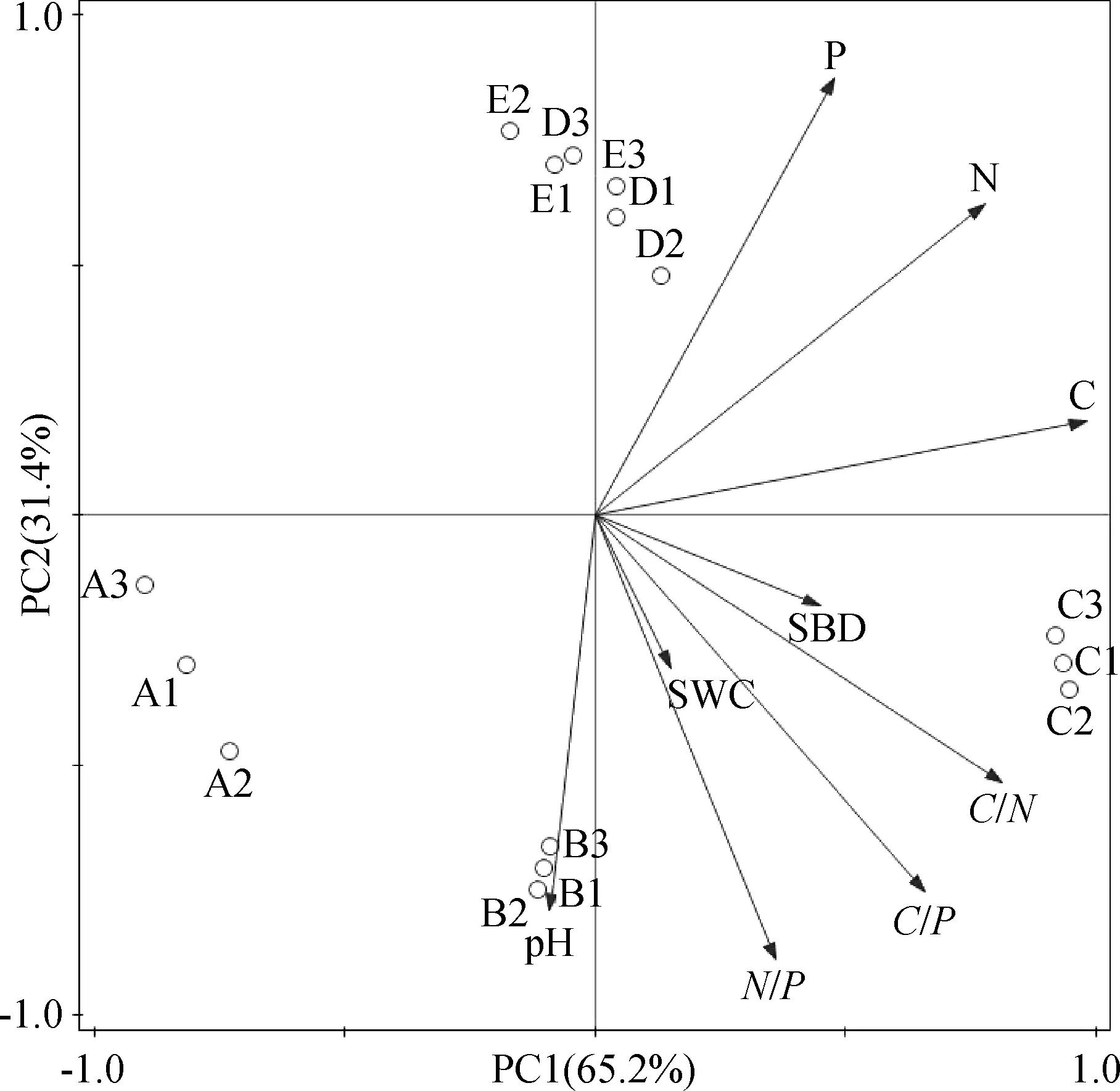
图2 不同海拔梯度土壤环境因子间的主成分分析Fig.2 Principal component analysis of soil environmental factors with different altitude
3 讨论
3.1 高寒草原植被对海拔变化的响应
alpha多样性能够反映出生态系统群落植被的丰富度、均匀度和多样性的整体规律,也可表示群落物种对环境的适应能力和群落受干扰影响程度[16]。本研究中,植物Shannon指数随海拔升高而增大,说明由海拔升高引起的生境变化有利于植物的生长发育,这可从植被盖度和生物量亦随海拔升高而增加这一结果得到进一步佐证,另外,这也可能与高海拔地区人为干扰较少有关,研究区域草层高度随海拔升高而降低,这符合植物在高海拔地区对低温和强太阳辐射的适应机理[17]。
3.2 土壤养分含量对海拔及土层的响应
草地土壤是草地生态系统的重要基础环境,土壤养分的分解与积累受到自然与人为干扰的影响外,还与气候、土壤养分状况和植被类型等外界环境因子息息相关[18-19]。本研究发现,随着海拔的升高,土壤pH值与含水量并没有增加,这与刀静梅等人的研究结果一致[20]。土壤容重随海拔升高而逐渐降低,与马维伟等人的研究结果一致[21],这种变化规律可能与高寒地区太阳辐射和气候温度有关,随海拔升高温度降低,而太阳的辐射量增强,从而导致土壤结构的改变。同样,这种生境变化也导致微生物分解有机质缓慢,易于积累,使土壤容重逐渐降低。植物根系主要分布在土壤表层,较深处的土层根系分布较少,其对土壤的穿插作用变弱[22],因此,形成土壤容重随土层深度增加而增加的结果,这与其他研究者对我国主要草地土壤容重垂直变化特征的相关研究结果一致[23-24]。与土壤容重的变化规律相对应,土壤孔隙度也随土层由浅到深逐渐减小,水分下渗困难,外加植物蒸腾和土壤蒸发等损耗,到达下层的水分越来越少,出现土壤水分随土层深度增加而呈下降趋势,这与李硕等的研究结果一致[25]。
3.3 土壤有机碳、全氮和全磷对海拔及土层的响应
本研究中,高海拔处土壤有机碳和全氮含量明显高于低海拔地区,这是由于海拔升高后,地上生物量增加,微生物活性下降,土壤有机碳氮矿化速率随之减慢所致;且高海拔地区植被根系生命周期较短,死亡后根系为土壤有机质的积累提供有利条件[26]。海拔3 490 m处土壤有机碳和全氮含量最高,可能是因为该海拔物种多样性较为丰富,植被残落物在以真菌为主的微生物作用下进行特殊的腐殖化过程。韦应莉等通过对祁连山土壤微生物研究发现,土壤微生物与有机碳呈显著正相关[27]。土壤全磷含量随海拔的升高呈现上升趋势,这与刘爱琴等研究结果一致[28]。导致全磷含量上升的原因可能是由各海拔区域土壤母质及地上生物量不同而造成的。土壤有机碳是表征土壤肥力的重要参数[29],其含量随着土层深度增加而降低,这与李亚娟等研究结果相一致[30]。随土层深度的加深,各海拔土壤全氮、全磷含量均呈下降趋势,这与前人研究结果相一致[31]。
3.4 土壤生态化学计量比对海拔、植物及土层的响应
植物体内的物质由结构性物质、功能性物质和贮藏性物质组成,碳作为植物各种生理生化过程的底物和能量来源,属于结构性物质,氮和磷是功能性物质,作为蛋白质和遗传物质的重要组成元素[32]。植物碳吸收的途径不同,在自然条件下,一般认为碳不会限制植物生长,氮和磷的变化是影响C/N和C/P的主要因子。本研究中植被盖度与Shannon指数变化规律与土壤N/P变化规律相同,说明植物生长与物种多样性主要受氮素的影响。
土壤生态化学计量比(C/N,C/P,N/P)是衡量土壤质量的重要参数[33],反映了土壤释放氮、磷矿化养分的能力。受气候、地貌、土壤生物和人为干扰等影响,土壤碳氮磷总量变化差异较大[34]。微生物生命活动所需的最佳C/N为25∶1[35],此比值对有机碳转化最有利,小于此比值,有机碳转化较容易,能产生充足的氮素,大于此比值,有机碳转化困难,会导致微生物与植物争夺氮素的现象,但土壤有机碳积累会更加容易[36]。中国土壤的C/N平均值在10~12[37],本研究发现研究区土壤的C/N范围为13.65~21.35,土壤C/N与全N之间相关性不显著,与有机碳呈显著正相关,说明该地区土壤C/N主要受碳控制,且C/N比值小,在其矿化作用之始就能供应给植物所需的有效氮量,有机质分解速率较快[38]。土壤C/P被认为是指示土壤磷元素矿化能力的重要指标,可以衡量土壤有机质矿化释放磷或吸收固持磷的潜力,较高的C/P不利于微生物在有机质分解过程中的养分释放,使土壤中有效磷的含量减少[39],我国土壤C/P平均值为105,变化范围为1.05~177.69[40],本研究发现,各海拔土壤C/P范围为39.37~90.58,低于我国平均值,意味着研究区土壤碳、磷有效性较高。N/P作为判断限制生产力的养分因子指标,将N/P<10和N/P>20作为评价植被生产力受氮或者磷限制的指标[41]。本研究中土壤N/P的范围为2.88~4.24,因此,在不同海拔梯度中,植物生产力主要受氮素的影响。
4 结论
在三江源区,随着海拔的升高,草层高度下降,草地盖度增加,植物Shannon指数增加,土壤中全磷含量逐渐升高,土壤容重、碳、氮、C/N、C/P和N/P含量在海拔3 490 m处达到最高后又逐渐下降,草地生物量、土壤pH与含水量无明显变化规律;随土层加深,土壤pH、含水量、碳、氮、磷、C/P和N/P均有不同程度的下降,土壤容重和C/N呈上升趋势,土壤有机碳和氮含量有明显的表聚现象;土壤氮与碳、磷极显著正相关,海拔与氮、磷极显著正相关、与N/P极显著负相关。高寒草原植物生产力主要受氮素的影响。
猜你喜欢
数学物理学报(2022年1期)2022-03-16
散文诗(2021年22期)2022-01-12
数学物理学报(2021年6期)2021-12-21
数学物理学报(2021年5期)2021-11-19
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
环球时报(2019-05-23)2019-05-23
中国农业文摘-农业工程(2016年5期)2016-04-12
亚热带资源与环境学报(2015年1期)2015-01-22
植物营养与肥料学报(2014年1期)2014-03-11
郑州大学学报(工学版)(2014年6期)2014-03-01
