
基于温度梯度的带壳鲜花生热风干燥收缩模型研究
2019-09-24 00:42卢映洁任广跃张乐道凌铮铮
食品与机械 2019年8期
卢映洁 任广跃,2 段 续,2 张乐道,2 凌铮铮
(1. 河南科技大学食品与生物工程学院,河南 洛阳 471023;2. 粮食储藏安全河南省协同创新中心,河南 洛阳 471023)
目前,中国对刚收获的花生常用的干燥方法除自然晾晒外,还采用热风、热泵、微波等机械干燥方法[1-3],极大提高了干燥效率,但快速干燥的均匀性较差,干燥品质无法得到保障。带壳鲜花生本身为复杂的多孔双层结构,且在干燥过程中可能会发生收缩,故不考虑结构变化和收缩情况,难以有针对性地对带壳鲜花生进行机械干燥。
大多数农产品属于可变形的植物基多孔介质[4],在干燥过程中会有一定的体积收缩,而收缩变形是影响其干燥品质与效率的重要因素之一。Seerangurayar等[5]对比了3种太阳能干燥方式对大枣收缩率的影响,通过显微结构分析,发现强制对流太阳能干燥的组织变形最小,干燥效果最优。白竣文等[6]研究了大野芋薄层干燥特性,发现干燥温度和切片厚度均对大野芋收缩率有显著影响,采用较低的干燥温度和较厚的切片厚度能够提高收缩率,减少收缩。Aprajeeta等[7]指出在62 ℃干燥条件下,马铃薯片的收缩率随含水率呈线性变化,其径向尺寸减小约35%,且收缩会随热量和质量的同时传递而发生变化。李建欢等[8]发现在热风干燥过程中,澳洲坚果的果壳收缩量由内至外逐渐增大,收缩不均匀,且收缩量随含水率的降低呈非线性增长趋势。Sagar等[9]对果蔬对流干燥收缩模型的研究进行了综述,对比分析了不同果蔬材料的收缩模型。陈良元等[10]研究表明,切片茄子的体积收缩影响干燥传质过程,收缩导致水分扩散内部阻力增大,在未考虑干燥收缩对动力学影响时,水分有效扩散系数会被明显高估。物料收缩与其干燥过程中热质传递及应力应变机制密切相关[11],引入收缩模型并分析其在干燥过程中结构的变化可对干燥机理进行更深入的探讨。然而,关于收缩对带壳鲜花生干燥特性影响的研究未见报道。
试验拟采用热风干燥对带壳鲜花生进行研究,考察不同温度(40,50,60 ℃)对带壳鲜花生干燥收缩特性的影响,并建立相应的数学模型,采用扫描电镜(scanning electron microscope, SEM)等从微观结构、空隙量、内部温度等干燥收缩特性分析干燥过程中收缩对带壳鲜花生的影响,以期为规模化控制干燥制定合理的干燥工艺和品质研究提供理论依据。
1 材料与方法
1.1 材料与仪器
带壳鲜花生:购于河南洛阳丹尼斯超市;
电热鼓风干燥箱:101型,北京科伟永兴仪器有限公司;
扫描电镜:TM3030plus型,日立高新技术公司;
电子天平:A.2003N型,上海佑科仪器仪表有限公司;
数控笔式温度计:KT300型,欧达时科技(香港)有限公司。
1.2 方法
1.2.1 原料预处理 在试验前挑选大小均匀成熟饱满的花生,清除杂质、泥沙,用自封袋封装于4 ℃冰箱中保存备用。采用GB 5009.3—2016测定带壳鲜花生、鲜花生壳、鲜花生仁的初始干基含水率分别为0.736,0.931,0.545 g/g。
1.2.2 试验设计 设定温度(40,50,60 ℃),预热30 min,将带壳鲜花生在网状托盘(25 cm×25 cm,筛孔直径为5 mm)上平铺一层(约500 g)。因风速(≤2 m/s)对带壳鲜花生体积收缩的影响不显著,故仅考虑热风干燥温度对带壳鲜花生热风干燥收缩特性的影响。取两盘花生同步进行干燥,一盘每隔1 h测定样品的质量,快速测量后放回,另一盘每隔1 h取30个样品进行留样保存,此盘取出的样品不再放回。干燥至安全水分(10%)停止试验。每组试验重复3次。
1.2.3 干基含水率和水分比的计算 带壳鲜花生的干基含水率和水分比分别按式(1)、(2)计算:
(1)
式中:
X——干基含水率,g/g;
mt——t时刻物料的质量,g;
m——物料干燥至恒重时的质量,g。
(2)
式中:
MR——水分比;
Xo——初始干基含水率,g/g;
Xt——t时刻干基含水率,g/g;
Xe——平衡时刻干基含水率,g/g。
1.2.4 SEM分析 在带壳花生下半部分饱满处进行取样,固定于样品台,利用SEM检测干燥处理后的花生壳、花生仁表面的微观结构,电镜放大倍数200倍。
1.2.5 空隙量的测定 为更好地反映带壳鲜花生在热风干燥过程中的收缩变化,采用注水法测定壳内空隙。在带壳花生的顶端开一小裂缝,用5 mL针头注射器吸取蒸馏水,缓慢注入花生壳内,直至水溢出花生壳,记录注水量,记为空隙量(cm3)。每组试验取20个样品测量,结果取平均值。
1.2.6 内部温度的测定 为反映带壳鲜花生的花生壳与花生仁之间的收缩差异,对带壳鲜花生内部温度进行测量。采用笔式温度计对内部温度进行实时测量,记录读数。每组试验取5个样品进行测量,结果取平均值。
1.2.7 收缩比与收缩速率的计算 采用排沙法测体积[12],分别按式(3)、(4)计算样品体积相对收缩比和收缩速率。
(3)
式中:
SR——收缩比;
Vt——任意t时刻的体积,m3;
V0——初始体积,m3。
(4)
式中:
W——收缩速率,h-1;
SRt——t时刻收缩比;
SRt+Δt——t+Δt时刻收缩比;
Δt——时间间隔,h。
1.2.8 收缩模型的选择 在理想的收缩条件下,物料体积的减少等于除去液体的体积,因此常用收缩比与水分比的函数来拟合收缩[13-15]。试验根据花生的自身特性,选取以下4种收缩模型(见表1)。

表1 4种常见收缩模型†Table 1 Four common contracting model
†k1~k5分别表示模型常数。
1.3 数据处理
采用Origin 8.5对试验数据进行线性/非线性拟合,并分析其拟合度;使用DPS 7.05对试验数据进行显著性分析。
2 结果与分析
2.1 温度对带壳鲜花生热风干燥收缩特性的影响
从图1(a)可知,随着热风干燥温度升高,花生壳的收缩比逐渐减小,当干燥温度为40,50,60 ℃时,其干燥至平衡时的收缩比分别为0.857,0.819,0.777。温度提高20 ℃,收缩比减小8.0%。由图1(b)可知,带壳鲜花生热风干燥的体积收缩速率具有短暂的升速阶段,然后进入降速阶段,最后基本为恒速阶段。水分是支撑花生壳组织饱满的重要物质,干燥前期,带壳鲜花生主要为自由水的减少,大部分热量传递给花生中的水分,以提高花生中水分的蒸发温度,随着花生壳中水分温度的逐渐升高,花生壳水分蒸发量逐渐增大,形成水分梯度,内部扩散速率也逐渐加快,水分迁移速度快,体积变化明显,收缩速率高。随着干燥的进行,后期内部主要为结合水,而结合水不易散失,收缩速率自然降低。随着温度的升高,收缩速率曲线越陡峭,由于干燥温度是影响干燥效果的主要因素[18]。干燥温度越高,水分流失越快,收缩越明显,速率越快。
由图2(a)可知,随着热风干燥温度的升高,花生仁的收缩比逐渐减小,当干燥温度为40,50,60 ℃时,其干燥至平衡时的收缩比分别为0.695,0.659,0.624。温度升高20 ℃,收缩比减小7.1%,说明温度越高,花生仁的收缩程度越大。由图2(b)可知,花生仁的收缩速率曲线呈先升高再降低的趋势,且温度越高,收缩越快。可能是在干燥前期,花生壳作为保护屏障,阻挡热量进入壳内,壳内温度较低;随着干燥的进行,壳内温度逐渐升高,导致含水量较高的花生仁水分散失加快,组织收缩明显。在干燥后期,花生壳内温度稳定,花生仁水分含量也较低,组织结构收缩程度小,收缩速率变缓。

图1 不同温度下带壳鲜花生花生壳的收缩曲线及收缩速率曲线
Figure 1 Contraction curve and contraction rate curve of peanut shell of fresh shelled peanuts under different temperatures
对比花生仁与花生壳的收缩曲线和收缩速率曲线可知,相同条件下,花生仁的收缩比比花生壳的收缩更加明显,说明花生壳与花生仁的收缩非同步进行。花生壳干燥初期有较为剧烈的收缩,中后期持续缓慢收缩,但花生仁在干燥初期收缩速率缓慢,干燥中期收缩剧烈,可能是热风干燥时,花生壳先于花生仁接触到高温环境,先失水收缩,待花生壳失去一部分水分后,花生仁才发生收缩。花生壳与花生仁绝干时干基含水率分别为0.931,0.545 g/g,花生壳的干基含水率高于花生仁,收缩程度却不及花生仁,可能是由于花生仁较厚,可发生形变的范围大;花生壳主要成分为粗纤维素(65.7%~79.3%)和半纤维素(10.1%)[19],干物质重量轻,通道大,可包容的水分较多,有良好的网状结构,而干燥使得通道中的水分散失,结构更为致密,但并不会导致网状结构的坍塌,故不易发生形变,而花生仁中主要成分为脂肪(44%~45%)、蛋白质(24%~36%)[20]等,细胞中的水分散失后,无交联的网状细胞结构作为支撑,容易发生形变。针对干燥过程中花生壳优先花生仁干燥且收缩比小于花生仁的情况,可以考虑变温干燥或联合干燥提高干燥效率,改善干燥品质。
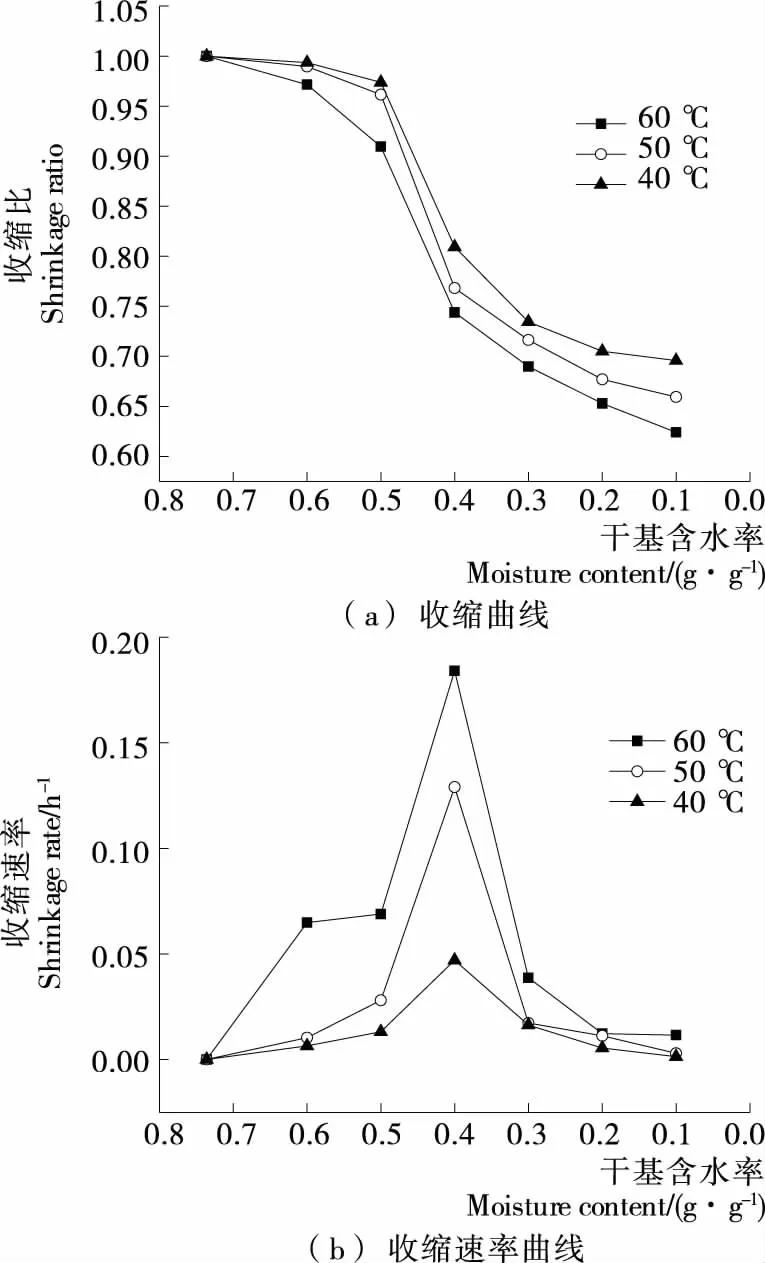
图2 不同温度下带壳鲜花生花生仁的收缩曲线及收缩速率曲线
Figure 2 Contraction curve and contraction rate curve of peanut kernel of fresh shelled peanuts under different temperatures
2.2 温度对带壳鲜花生空隙量的影响
由图3可知,同一温度下,随着干燥的进行,花生壳与花生仁之间的空隙逐渐增大,为二者共同收缩所致。干燥初期,花生壳为主要收缩对象,且是花生壳内部结构的收缩,空隙量较小;随着干燥进行,花生仁开始收缩且收缩较为严重,空隙量快速增大。另外,花生壳与花生仁之间出现的空隙可能会形成内外压力梯度差,并增加花生仁水分迁移路径,从而影响干燥效率。在40,50,60 ℃干燥至终点时,其空隙量分别为0.93,1.05,1.25 cm3。干燥温度越高,空隙量越大且空隙量的曲线越陡,表明温度对空隙量的影响较为显著。从侧面反映温度能引起收缩,温度越高,收缩越严重,产生的空隙越大。
2.3 温度对带壳鲜花生内部温度的影响
由图4可知,随着干基含水率的降低,带壳鲜花生内部温度逐渐升高,干燥温度越高,内部升温速度越快。在40,50,60 ℃干燥至终点时,其内部温度分别为33.8,41.8,45.5 ℃。在较高干燥温度下,内部温度基本呈直线上升趋势。结合图2可知,花生仁的收缩主要发生在干基含水率为0.4 g/g左右,表明花生壳对花生仁有保护作用,能减缓热传递,壳外温度远大于壳内温度,故花生壳先于花生仁收缩。在60 ℃干燥至终点时,内部温度仍有上升趋势,说明花生壳的保护作用较强,在干燥时间范围内外部温度不能完全穿过花生壳加热花生仁,从而影响干燥效率。因此,在保证花生仁收缩不明显的前提下,可以采取前期高温后期低温的变温干燥或不同干燥方式的联合干燥来优化带壳鲜花生的干燥工艺。
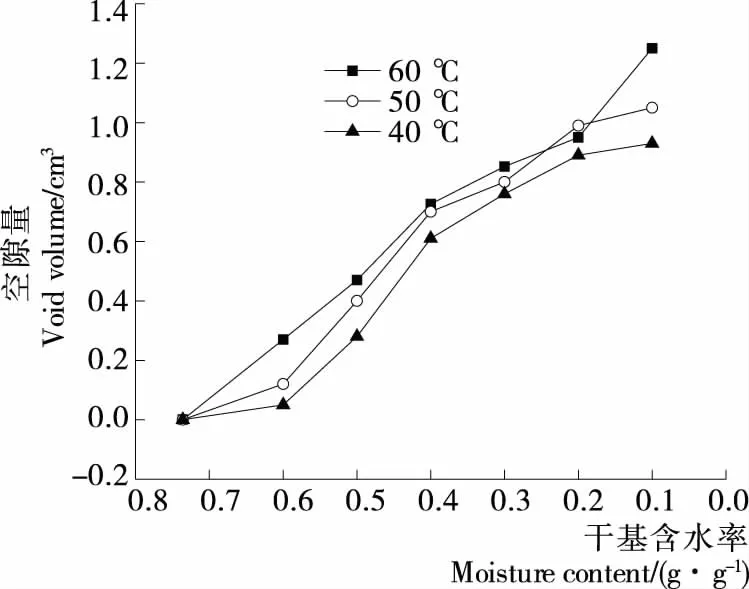
图3 不同温度下带壳鲜花生空隙量的变化Figure 3 Changes of void volume of fresh shelled peanuts under different temperatures

图4 不同温度下带壳鲜花生内部温度的变化Figure 4 Changes of internal temperature of fresh shelled peanuts under different temperatures
2.4 温度对带壳鲜花生微观结构的影响
在干燥过程中,花生壳与花生仁内部结构也会发生变化。为确定热风干燥在不同温度下对花生内部结构的影响,使用SEM对不同温度干燥处理过程中的花生壳与花生仁的表面进行观察,结果如图5、6所示。
由图5可知,对比相同温度下不同干基含水率的微观结构,花生壳表面最初为排列规律的蜂窝状结构,网孔边界清晰,排列相对规则。随着干燥的进行,花生仁的蜂窝状结构快速变形,结构排列杂乱,疏水通道变窄,从而导致内部花生仁的水分不易扩散至外界,影响干燥效率。对比不同温度下相同干基含水率的带壳鲜花生的孔隙结构,发现较高的干燥温度能加快水分散失,高温使得花生壳在干燥前期网状结构严重收缩,而40 ℃时花生壳的网状结构变化较为缓慢。干燥至平衡时,60 ℃的结构比40 ℃ 的结构变形更为剧烈。从侧面反映了干燥温度越高,花生壳的体积收缩越明显。由图6可知,新鲜花生仁的表面结构呈现出饱满蓬松、似“玉米粒”的构造。随着干燥的进行,花生仁表面结构逐渐收缩变形,排列杂乱,细胞壁出现褶皱、收缩、卷曲现象,说明花生仁的细胞内主要为水分,当水分逐渐迁移后,剩下的大部分物质为脂肪和蛋白质,两种物质导致花生仁比花生壳的收缩更为明显,与图2结果一致。且干燥温度的升高使花生仁的细胞脱水速度加快,导致细胞结构变化明显,收缩加快。
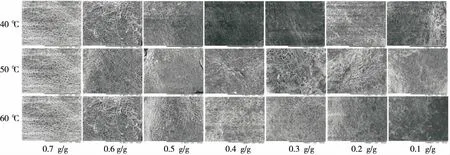
图5 不同温度下带壳鲜花生花生壳的SEM图Figure 5 SEM of peanut shells of fresh shelled peanuts under different temperatures

图6 不同温度下带壳鲜花生花生仁的SEM图Figure 6 SEM of peanut kernel of fresh shelled peanuts under different temperatures
结合图5、6可知,干燥至0.4 g/g左右时,花生壳的微观结构几乎完全变形,而此时对应的花生仁表面结构变形程度较为剧烈,说明干燥至0.4 g/g左右时,可视为干燥中的一个转折点,花生壳大部分水分在干燥初期丢失,先于花生仁完成干燥。
2.5 带壳鲜花生热风干燥体积收缩模型的建立
对50 ℃条件下带壳花生与花生仁热风干燥的收缩比进行分析,选取4个收缩模型(表1)用Origin 8.5对其进行非线性拟合,得出相应的R2(相关系数)、RSS(残差平方和)、X2和模型系数,见表2。R2越大,RSS及X2越小,数据拟合结果越好。通过对比分析4种模型的R2、X2、RSS可知,在50 ℃条件下,花生壳的Quadratic模型R2最大,为0.990 1,X2为3.375 1×10-5,RSS为1.350 0×10-4,数值较小,且Quadratic模型的表达式更为简便,故选择Quadratic模型作为花生壳的最优收缩模型;花生仁的Vazquez模型R2值最大,为0.967 5,X2为0.002 3,RSS为0.004 5,数值较小,故选择Vazquez模型为花生仁的最佳收缩模型。两种模型能较为准确地反映带壳鲜花生在热风干燥过程中的体积收缩特性,为带壳鲜花生的热风干燥规模化控制提供理论依据。
2.6 带壳鲜花生热风干燥体积收缩模型的验证
选取不同干燥温度(40,50,60 ℃)条件下的试验值和最终模型预测值进行验证比较,结果如图7、8所示。
由图7可知,试验值与模型预测值的吻合程度较高,说明模型的拟合程度较好。对不同温度下试验值与Quadratic模型预测值进行相关性分析,40,50,60 ℃条件下的花生壳试验值与预测值相关性分别为0.99,0.83,0.99(P<0.01),说明不同温度下试验值与预测值都呈极显著的正相关。结果表明试验所建立的模型准确可靠,能够预测花生壳在干燥过程中收缩比随水分比的变化规律。

表2 各干燥收缩模型的统计分析结果Table 2 Statistical analysis results of each dry contraction model

图7 不同温度下花生壳试验值与Quadratic模型预测值比较
Figure 7 Comparison between test values at different air temperatures and predicted values of Quadratic model

图8 不同温度下花生仁试验值与Vazquez模型预测值比较
Figure 8 Comparison between test values at different air temperatures and predicted values of Vazquez model
由图8可知,试验值与模型预测值的吻合程度较高,说明模型的拟合程度较好。对不同温度下试验值与Vazquez模型预测值进行相关性分析,40,50,60 ℃条件下的花生仁试验值与预测值相关性分别为0.97,0.95,0.94(P<0.01),说明不同温度下试验值与预测值都呈极显著的正相关。结果表明试验所建立的模型准确可靠,能够预测花生仁在干燥过程中收缩比随水分比的变化规律。
3 结论
在试验温度范围内,较高的干燥温度会对带壳鲜花生的干燥品质产生一定的影响。选取较低的干燥温度,干燥进程缓慢,壳内温度变化幅度小,花生壳与花生仁的微观结构变化程度小,能减小带壳鲜花生的收缩,保证花生的干燥品质。花生壳先于花生仁收缩,但收缩程度不及花生仁,可能是花生壳纤维结构的收缩不及花生仁细胞结构的收缩。花生壳与花生仁之间的空隙随着干燥的进行而逐渐增大,可能会形成内外压力梯度差并增加花生仁水分迁移路径,且为了减少收缩,可以考虑变温干燥或联合干燥工艺。
对不同收缩模型分析表明,花生壳采用Quadratic收缩模型拟合程度较高,花生仁采用Vazquez模型拟合程度较高,两者能很好地反映带壳鲜花生热风干燥过程中体积收缩的变化。因此,该研究结果能更准确地预测带壳鲜花生热风干燥过程中干燥特性的变化规律,进一步保证带壳鲜花生的干燥品质。
猜你喜欢
动漫界·幼教365(小班)(2021年11期)2021-03-23
小天使·聪聪画刊(2020年11期)2020-09-10
启蒙(2020年2期)2020-02-12
中国油脂(2020年1期)2020-01-14
满族文学(2019年5期)2019-10-08
江苏大学学报(自然科学版)(2018年6期)2018-11-15
领导决策信息(2017年14期)2017-06-21
中国粮油学报(2016年5期)2016-01-23
小学生导刊(中年级)(2014年11期)2014-11-20
