
砂粒粒径对日本囊对虾存活、生长及行为特性的影响
2019-09-18 10:29王庚申谢建军许文军
水产科学 2019年5期
王庚申,谢建军,施 慧,何 杰,汪 玮,许文军
( 1.浙江省海洋水产研究所,浙江 舟山 316021; 2.浙江省海水增养殖重点实验室,浙江 舟山 316021 )
日本囊对虾(Marsupenaeusjaponicus)俗称花虾、车虾、竹节虾等,隶属甲壳纲、十足目、游泳亚目、对虾科、囊对虾属,在中国、日本、东南亚、澳大利亚、非洲及红海等海区均有分布,是我国重要的虾类养殖品种[1-2]。其味道鲜美,营养丰富,市场价格高,且具生长快、耐低温、耐干露等特点,养殖潜力巨大,深受消费者和养殖业者的喜爱。我国20世纪80年代开展全人工养殖以来,沿海各省市均有养殖,并形成一定的规模[3]。据统计,2017年全国日本囊对虾养殖面积2.57×104hm2,产量5.25×104t,取得了较好的经济和社会效益[4]。近年来,日本囊对虾养殖技术不断得到完善和提高,池底覆砂的工厂化养殖和高位池精养模式已成为主要养殖模式[5]。
底质是养殖生态系统的重要组成部分,不仅能够提供营养、稳定水质及改变水环境,同时也可以作为养殖生物和微生物生长繁殖的场所[6]。在工厂化和高密度精养模式下,养殖环境与对虾的自然生境差异较大,养殖密度增加会限制对虾特有的一些行为,从而导致对虾部分行为的缺失或异常。动物的行为和功能与其生存、适应环境的关系十分密切,行为的缺失或异常会影响动物对环境变化的调节与整合,也就无法通过其特有的行为来维持体内平衡,致使机体产生应激反应,添加底质则可以中和由于养殖密度增加而带来的不良影响[7-9]。对虾绝大部分时间与底质接触,因此,底质的选择在对虾分布、寻食、躲避攻击和栖息地选择等过程中均至关重要[10-12]。当前,国内外对日本囊对虾的研究主要涉及群体差异[13-16]、生理生化[17-18]、争胜行为[19-20]、病害防治[21-23]、免疫与营养[24-25]、遗传育种[26-27]等,对其底质选择等基础生物学研究较少。日本囊对虾有很强的潜砂习性,对底质要求较高,一些学者的研究仅涉及不同底质类型对日本囊对虾存活和生长的影响[28-29],而有关砂粒粒径对日本囊对虾影响的研究尚未见报道。刘勇克[30]研究了三疣梭子蟹(Portunustriuberbuculatus)在不同砂粒粒径底质中的潜砂频率;徐永健等[31]研究表明,砂粒粒径对梭子蟹幼蟹的摄食行为和生长特性均有明显的影响。本研究通过在实验室设置不同规格砂粒底质,研究砂粒粒径对日本囊对虾存活、生长及行为特性的影响,探讨日本囊对虾对砂粒粒径的选择和偏好,以期更加全面地了解日本囊对虾的行为习性,为日本囊对虾的精养模式提供理论参考和依据。
1 材料与方法
1.1 试验材料
试验于2016年7月22日—8月12日在浙江省海洋水产研究所试验场开展,所用试验虾取自试验场日本囊对虾精养塘,平均体长(2.76±0.18) cm,平均体质量(0.27±0.03) g,于室内水泥池中暂养7 d。暂养期间,微充气,日换水量30%,每日定时过量投喂对虾配合饲料,及时清除残饵和粪便。试验砂购于市场,预先经不同规格的筛网筛选,并用高锰酸钾消毒,清水冲洗,晾干后使用。
1.2 试验设计
试验容器为160 L塑料水槽,共设3个砂粒粒径组:细砂组(粒径<0.5 mm)、中砂组(粒径1~2 mm)、粗砂组(粒径>2.5 mm),砂层厚度为10 cm,以无砂组为对照组,每组3个平行。暂养结束,每组放入规格一致的健康日本囊对虾幼虾40尾,适应24 h后开始正式试验。试验周期为20 d。
1.3 试验指标统计与分析
1.3.1 生长指标
成活率/%=n2/n1×100%
特定生长率/%·d-1=(lnm2-lnm1)/t×100%
蜕皮频率/次·(尾·d)-1=n3/(n0×t)
式中,n1和n2分别为试验开始和结束时试验虾的数量
(尾),m1和m2分别为试验开始和结束时试验虾的湿质量(g),n3为试验时间t(d)内试验虾的蜕皮次数(次),n0为试验虾尾数(尾)。
红军在进入川西北地区之初,就这样争取少数民族的支持并使之投身革命斗争,从封建压迫下解放出来的问题,进行了仔细的研究,显示出中央对这一问题的极大关注,意识到了群众工作的重要性。
1.3.2 行为特性
寻食虾比率/%=n4/n0×100%
运动率/%=n5/n0×100%
攻击频率/次·(尾·h)-1=n6/(n0×t)
潜砂时间/s=t1/n
式中,n0为试验虾数量(尾),n4为寻食虾数量(尾),n5为处于运动状态下的试验虾数量(尾),n6为时间t(h)内试验虾发生攻击行为的次数(次),t1为试验虾潜砂消耗的时间之和(s),n为潜砂总次数(次)。
1.4 数据处理
使用SPSS 20.0分析数据,利用单因素方差分析和Duncan多重比较法检验不同处理组间的差异,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 对体色和表观形态的影响
通过观察发现,试验第4 d,无砂组日本囊对虾体色出现明显变化,浅绿色或透明,与正常虾黄褐色不同,但活力良好、摄食正常;随后在第7~10 d,3个砂粒底质组个体也出现体色改变。至试验结束,无砂组对虾体色改变比例最高,其次是粗砂组和中砂组,细砂组最低(P<0.05)。
表观形态变化定义为日本囊对虾的第二触角是否断裂,以第二触角完整比例计。经统计,细砂组日本囊对虾第二触角完整比例最高,分别是中砂组、粗砂组和无砂组的129.10%、141.75%和357.00%。多重比较结果表明,中砂组和粗砂组差异不显著(P>0.05),其他组之间差异显著(P<0.05)。

表1 不同处理组日本囊对虾体色和表观形态的比较 %
注:当对虾第二触角长度≥体长时,视为完整.同行不同上标字母表示差异显著(P<0.05).下同.
2.2 对生长指标的影响
日本囊对虾的成活率、特定生长率和蜕皮频率见表2。试验第4 d,无砂组日本囊对虾出现死亡,而中砂组、粗砂组和细砂组分别在第6 d、第8 d和第9 d出现死亡个体;第10 d后,各组试验虾的死亡频率明显增加,但日死亡数量≤3尾。细砂组成活率最高,显著高于其他组(P<0.05);无砂组成活率最低,与粗砂组差异不显著(P>0.05)。
各组日本囊对虾的特定生长率差异显著,细砂组特定生长率最高,显著高于无砂组(P<0.05),但与中砂组和粗砂组无显著差异(P>0.05)。无砂组的特定生长率最低,分别是细砂组、中砂组和粗砂组的77.23%、80.00%和80.41%。
日本囊对虾第1次蜕皮在试验第3 d,分别为粗砂组和无砂组,中砂组和细砂组分别在第4 d和第5 d;第5 d各组出现第1次蜕皮高峰,随后2 d蜕皮虾数量快速下降,直至第9~10 d和15~16 d再次出现蜕皮高峰。至试验结束时,无砂组蜕皮频率最高,分别是细砂组、中砂组和粗砂组的242.52%、238.00%和165.48%,显著高于其他组(P<0.05),但细砂组、中砂组和粗砂组之间差异不显著(P>0.05)。

表2 不同处理组日本囊对虾生长指标的比较
2.3 对行为特征的影响
各处理组日本囊对虾的行为特征见表3。投喂前,各组试验虾的运动率在无砂组中最高,显著高于3个砂粒底质组(P<0.05)。寻食虾比率则在粗砂组中最低,显著低于细砂组和无砂组(P<0.05),但与中砂组无显著差异(P>0.05)。攻击频率在无砂组最高,显著高于3个砂粒底质组(P<0.05);中砂组最低,与粗砂组无显著差异(P>0.05)。
投喂后,无砂组运动率显著高于3个砂粒底质组(P<0.05),而在砂粒底质组中,粗砂组运动率显著高于细砂组(P<0.05),但与中砂组差异不显著(P>0.05)。寻食虾比率则在细砂组中最高,显著高于其他3组(P<0.05);中砂组和无砂组之间差异不显著(P>0.05)。无砂组试验虾攻击频率最高,粗砂组最低,与中砂组之间差异不显著(P>0.05)。
相比之下,各砂粒组投喂后对虾的运动率低于投喂前,而投喂后攻击频率和寻食虾比率则高于投喂前。投喂后细砂组、中砂组、粗砂组和无砂组的运动率分别是投喂前的15.66%、46.38%、44.83%和55.34%,而寻食虾比率分别是投喂前的485.19%、462.22%、541.80%和330.76%,攻击频率分别是投喂前的292.16%、319.14%、239.43%和335.92%。
不同砂粒底质对日本囊对虾的潜砂时间影响显著,细砂组潜砂时间最长,显著高于其他组(P<0.05)。观察还发现,试验虾在3种砂粒底质中的埋栖深度不同,细砂组日本囊对虾完全埋栖或仅露出第一触角,而中砂组日本囊对虾会露出整个额角或1/3头胸甲,粗砂组日本囊对虾则露出大于1/2或整个头胸甲。
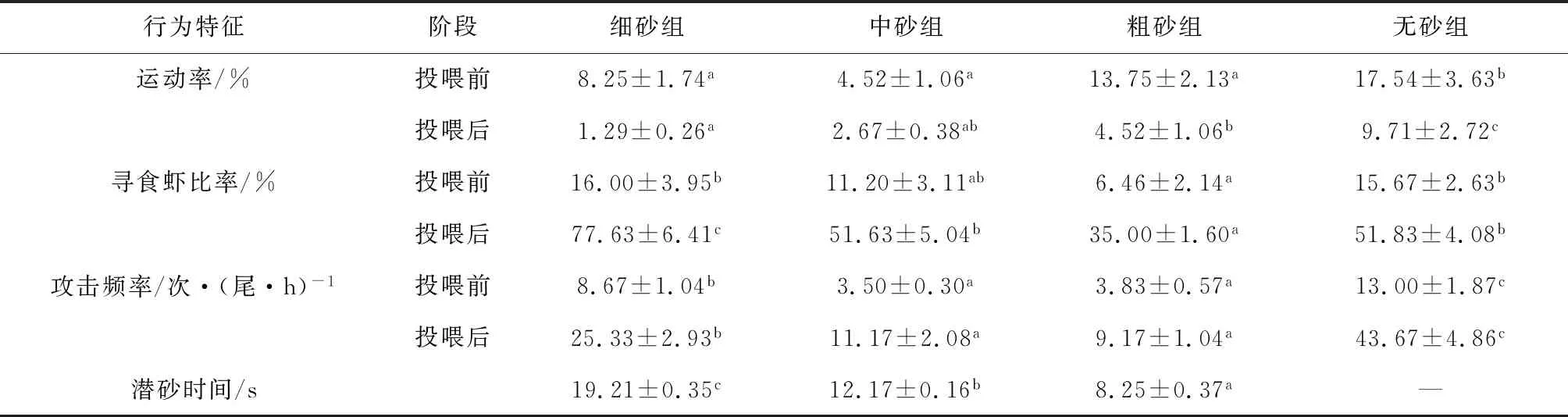
表3 不同砂质组日本囊对虾行为特征的比较
3 讨 论
3.1 砂粒粒径对日本囊对虾体色和表观形态的影响
对虾的甲壳中含有虾青素,煮熟或氧化后变为红色,对虾体色主要决定于虾青素的含量[32]。虾青素是一种类胡萝卜素,一般不能在动物体内合成,只能在体内积累[33]。有研究表明,虾青素的含量与光照及水环境密切相关[34],当对虾受光照刺激时间增加时,虾青素含量提高,对虾体色即表现出适应性变化。本研究中,无砂组对虾的体色改变比例高于3个砂粒底质组,与罗氏沼虾(Macrobrachiumrosenbergii)幼虾[35]在无底质组出现明显的体色改变一致。3个砂粒底质组中,细砂组对虾体色改变比例最低,与粗砂组和中砂组差异显著。由本研究结果可知,不同粒径砂粒的添加影响了日本囊对虾的潜砂行为,改变了其受光照刺激的时间,从而引起体色的适应性变化,即明显的体色改变。表观形态变化定义为第二触角的断裂,通常是由对虾的争胜行为所致。当缺少底质遮蔽物(无砂)时,日本囊对虾争胜行为发生频繁,其附肢残缺比例随之升高。3个砂粒底质组中,第二触角完整比例随着砂粒粒径的增大而降低,说明试验虾在细砂中埋栖程度最深,减少了争胜行为的发生。综合试验结果可知,添加底质对日本囊对虾的体色和表观形态均有显著影响,其中细砂组对虾体色改变比例最低,而第二触角完整比例最高,这表明细砂更有利于保持日本囊对虾正常的体色和表观形态。
3.2 砂粒粒径对日本囊对虾生长指标的影响
目前砂粒粒径对甲壳类生长和存活的研究报道较少,徐永健等[31]研究表明,砂粒粒径对单体筐养三疣梭子蟹的质量增加和成活影响显著,细砂粒组和混合砂粒组成活率、体质量增长率和蜕皮次数均显著高于粗砂粒组和无砂组。本研究中,无砂组日本囊对虾的蜕皮频率高于各砂粒组,而特定生长率则明显低于各砂粒组,这可能与无砂组试验虾受长时间光照使蜕皮周期缩短有关,而蜕皮过程需要消耗大量的能量,频繁蜕皮可能会影响对虾的生长[36-37]。3个砂粒组中,细砂组成活率和特定生长率均最高,蜕皮频率则最低,其中成活率显著高于中砂组和粗砂组,而特定生长率和蜕皮频率无显著差异。由本研究结果可知,添加砂粒可明显提高日本囊对虾的成活率和特定生长率,这与小褐虾(Crangonuritai)[38]的研究一致。在不同砂粒组中,随着砂粒粒径的增大,日本囊对虾成活率下降。这可能是因为砂粒满足了日本囊对虾的潜砂特性,同时为其活动及蜕皮提供了隐蔽场所;当砂粒粒径过大时,对虾埋栖深度变浅,易暴露,砂底的遮蔽作用减弱,导致对虾成活率下降。
3.3 砂粒粒径对日本囊对虾行为特征的影响
行为是生物应对外界环境变化的表观反应,在外界环境变化时进行调节和整合,以保证生命活动的正常进行[39]。而底质是对虾主要的生长和栖息环境,在养殖生态系统中具有重要作用,与对虾的摄食、攻击、潜砂等行为特征密切相关。臧维玲等[28]试验观察发现,日本囊对虾幼虾有底质组的摄食积极性及摄食频率均低于无底质组。张沛东等[6]也指出,中国明对虾(Fenneropenaeuschinensis)在有底质组的寻食虾比率和运动率均高于无底质组,攻击频率则低于无底质组。本研究发现,投喂后各组试验虾的运动率明显低于投喂前,而投喂后的攻击频率和寻食虾比率则高于投喂前,这说明日本囊对虾摄食积极,导致运动虾数量下降,同时在摄食中争胜行为发生频率更高。投喂前后,日本囊对虾在无砂组的运动率、攻击频率均显著高于3个砂粒底质组,而寻食虾比率则低于细砂组,但高于中砂组和粗砂组,与中国明对虾[6]的研究不同,可能与日本囊对虾更强的潜砂习性有关,也可能是因为试验所设置的底质不同。在砂粒底质组中,投喂前试验虾的运动率无明显规律,投喂后运动率随着砂粒粒径增大而升高,而寻食虾比率和攻击频率在投喂前后均随着砂粒粒径的增大而下降,说明日本囊对虾在细砂组摄食积极性更高,但由寻食而引起的攻击行为也更频繁。
本研究还发现,日本囊对虾的潜砂时间随着砂粒粒径的增大而减少,埋栖深度则随着砂粒粒径增大而变浅,这与臧维玲等[28]观察日本囊对虾幼虾[平均体长(1±0.1) cm]在细砂中易潜入,在2~3 mm砂粒中不潜入的结果不同。分析可能是由于试验虾规格差异导致对砂粒粒径的适应性不同,一般认为,随着对虾规格的增大,潜砂能力增强,对砂粒粒径的适应能力也增强。
本研究结果表明,砂粒底质的添加更有利于日本囊对虾生长与存活,保持正常的体色和表观形态,同时适宜的底质也能促进其行为的调节,如促进潜砂、提高摄食积极性等。综合比较而言,3种砂粒粒径底质中,日本囊对虾更偏好砂粒粒径<0.5 mm的细砂。另外,本研究所选取的试验虾为单一规格,因此在将来的研究中,进一步探讨不同规格试验虾对砂粒粒径的选择,可以更加全面地为日本囊对虾养殖生产提供参考。
猜你喜欢
海洋通报(2022年2期)2022-06-30
安徽建筑(2021年11期)2021-11-12
济南大学学报(自然科学版)(2021年2期)2021-03-04
新少年(2020年10期)2020-10-30
Medical Gas Research(2019年4期)2020-01-07
上海航天(2018年3期)2018-06-25
启迪与智慧·教育版(2018年12期)2018-02-26
中国房地产业·下旬(2017年12期)2017-07-09
天津农业科学(2015年5期)2015-05-30
Beijing Review(2013年10期)2013-12-06
