
浑善达克沙地榆树疏林中木本植物空间格局及种内和种间关系分析
2019-09-12 06:17张志永时忠杰杨晓晖刘艳书
植物资源与环境学报 2019年3期
张志永, 时忠杰, 杨晓晖, 刘艳书, 张 晓,①
(中国林业科学研究院: a. 林业科技信息研究所, b. 荒漠化研究所, 北京 100091)
空间格局是指群落中植物个体在空间中的位置和散布状态,可反映植物之间以及植物与环境之间的相互作用[1-4]。分析物种的空间格局可揭示潜在的生态学信息[5-8];多数生态现象具有尺度依赖性,分析物种的空间格局时,如果仅采用某一固定尺度,其结果将具有一定的局限性[9]。点格局分析以离散的点坐标数据为基础,分析植物在多个尺度下的空间分布特征以及个体间的空间关联性[10-11],该方法很大程度克服了单一尺度分析的局限性[12-13]。
疏林草地约占地球陆地面积的12%[14],其主要特征是在草本植物连续覆盖的地面上稀疏分布着木本植物[15]。在内蒙古的浑善达克沙地中广泛分布着一类天然的温带疏林草地类型,因其乔木树种以榆树(UlmuspumilaLinn.)为主,因而通常被称为榆树疏林,是适应浑善达克沙地半干旱气候的顶极地带性群落[16]。相对于周边的草原生态系统,榆树疏林的生境多样性、生物多样性和结构多样性更高,对维持该区域的生态稳定和生态安全具有重要意义[17]。
对浑善达克沙地植被的研究主要分为3个阶段:在20世纪中期,主要从景观学角度开展浑善达克沙地的植被分布状况探查[18];到20世纪后期,主要在治沙和防护林建设背景下,重点研究小黄柳(SalixflavidaY. L. Chang et Skv.)等灌木在生态修复工程中的作用;而对浑善达克沙地内天然分布的榆树种群的关注则始于21世纪初期,从最初的榆树根系分布特征及生物量研究入手[19],逐渐认识到榆树疏林在空间分布上的异质性[20]。目前,针对浑善达克沙地榆树疏林的研究主要集中在榆树种群结构特征、空间异质性和种子库特征等方面[21-25],而针对榆树疏林对该区域治沙和防护林建设重要性的探索尚不够深入,尤其对包括榆树在内的主要木本种类的空间格局以及种内和种间关系还有待进一步研究。
作者采用点格局分析方法对浑善达克沙地榆树疏林中不同龄级榆树以及主要灌木种类的空间格局和空间关联性进行研究,探究榆树疏林空间格局形成的生态过程,以期加深对该区域木本植物空间分布特征的认识,为浑善达克沙地榆树疏林植被的恢复和保护提供基础数据。
1 研究区概况和研究方法
1.1 研究区概况
浑善达克沙地位于内蒙古高原中东部,地理坐标为北纬41°56′~44°26′、东经112°22′~117°57′。气候和土壤基本概况见文献[26]。
在该沙地的东南部现存有中国面积最大、群落结构最完整的榆树疏林景观[17]。研究区内的主要乔木树种为榆树;灌木种类主要有耧斗菜叶绣线菊(SpiraeaaquilegifoliaPall.)和小叶锦鸡儿(CaraganamicrophyllaLam.)等;草本种类主要有羊草〔Leymuschinensis(Trin.) Tzvel.〕和冰草〔Agropyroncristatum(Linn.) Gaertn.〕等[27]。
1.2 研究方法
1.2.1 样地设置和数据调查 在实地考察的基础上,在浑善达克沙地榆树疏林的东部(克什克腾旗浩来呼热乡附近)、南部(正蓝旗桑根达来镇附近)、西部(正蓝旗宝绍岱苏木附近)和北部(正蓝旗那日图苏木附近)分别选取具代表性的榆树疏林,于2015年8月在每个区域内分别设置1个面积100 m×100 m的样地,共4个样地。各样地的地理信息见表1。
表1 浑善达克沙地4个榆树疏林样地的地理信息
Table 1 Geographical information of four plots ofUlmuspumila-dominated savanna in Otindag Sandy Land
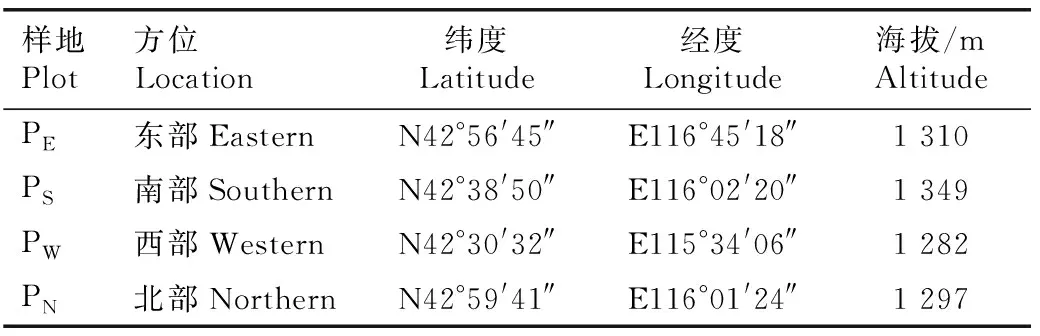
样地Plot方位Location纬度Latitude经度Longitude海拔/mAltitudePE东部EasternN42°56'45″E116°45'18″1 310PS南部SouthernN42°38'50″E116°02'20″1 349PW西部WesternN42°30'32″E115°34'06″1 282PN北部NorthernN42°59'41″E116°01'24″1 297
以样地西南角为原点,以西—东方向为横轴x、南—北方向为纵轴y,采用相邻网格法,将样地划分为25个20 m×20 m的网格,对样地内的所有木本植物进行调查,记录榆树的胸径(DBH)和空间位置;记录灌木种类以及株高较矮、无胸径(植株未达到测量高度)但茎已木质化的榆树幼苗的空间位置。以径级代替龄级,参照彭羽[28]提出的浑善达克沙地榆树龄级和径级拟合方程,将榆树植株划分为成年榆树(DBH≥20 cm)、中龄榆树(5 cm≤DBH<20 cm)、榆树幼树(DBH<5 cm)和榆树幼苗(无胸径)4个等级。
1.2.2 点格局分析 参考文献[4]的方法进行点格局分析。选择成对关联函数g(r)定量研究不同尺度上单一种类的空间格局以及2个种类间的空间关联性。按照公式“g(r)=dK(r)/(dr·2πr)-1”计算g(r),式中,r为空间尺度,K(r)为Ripley’sK函数,dK(r)是K(r)的微分,dr是r的微分。
在应用双变量g12(r)进行分析时,假设乔木层种类(成年树)对灌木层种类(榆树幼苗以及其他主要灌木种类)的生长具有影响,而灌木层种类对乔木层种类无影响,灌木层种类之间可以相互影响,因此,在分析乔木层种类和灌木层种类之间的关联性时,设置乔木层种类个体位置固定,灌木层种类的个体位置随机变化;而在分析灌木层种类之间的关联性时,则设置2个灌木种类的个体位置均随机变化。
1.3 数据处理和分析
采用Programita 2014软件进行数据分析。为精确分析各种类的空间格局以及不同种类之间的空间关联性,同时尽量降低边缘校正对结果的影响,本研究设定的空间分析尺度为0~50 m,步长为1 m。采用完全空间随机(complete spatial randomness,CSR)零模型和异质性泊松过程(heterogeneous poisson process,HP)零模型检验结果的显著性;在此基础上,用Monte Carlo检验拟合199次,产生置信水平为99%的上、下包迹线以检验结果的显著性。
2 结果和分析
2.1 不同龄级榆树和主要灌木种类的数量分析
对浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木的种类和数量进行调查和统计,结果见表2。结果显示:在供试的4个榆树疏林样地中,主要灌木种类均为耧斗菜叶绣线菊和小叶锦鸡儿,且各龄级榆树数量和灌木数量均存在明显差异。
在东部样地中,成年榆树(DBH≥20 cm)、中龄榆树(5 cm≤DBH<20 cm)和榆树幼苗(无胸径)的数量较多,均在60株以上,榆树幼树(DBH<5 cm)数量较少(11株);耧斗菜叶绣线菊和小叶锦鸡儿的数量也较少(分别有16和3株)。在南部样地中,成年榆树和榆树幼苗的数量较多(分别有82和47株),中龄榆树的数量较少(8株),且没有发现榆树幼树;耧斗菜叶绣线菊的数量较多(37株),小叶锦鸡儿的数量较少(4株)。在西部样地中,成年榆树和榆树幼苗的数量较多(分别有65和44株),中龄榆树的数量较少(7株),且没有发现榆树幼树;小叶锦鸡儿的数量较多(55株),耧斗菜叶绣线菊的数量较少(8株)。在北部样地中,成年榆树和榆树幼苗的数量较多(分别有108和89株),中龄榆树和榆树幼树的数量较少(分别有12和10株);耧斗菜叶绣线菊的数量较多(41株),小叶锦鸡儿的数量较少(11株)。
表2 浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的数量比较
Table 2 Comparison on numbers ofUlmuspumilaLinn. at different age classes and main shrub species in four plots ofU.pumila-dominated savanna in Otindag Sandy Land
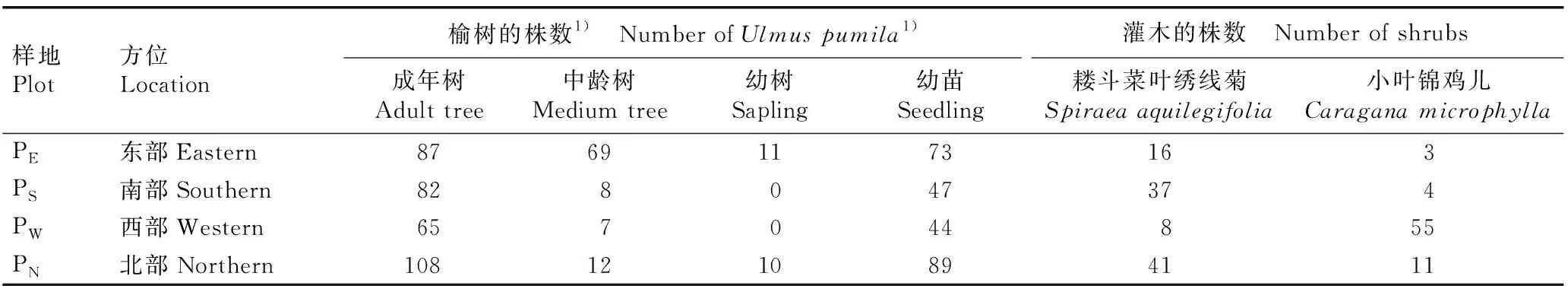
样地Plot方位Location榆树的株数1) Number of Ulmus pumila1)灌木的株数 Number of shrubs成年树Adult tree中龄树Medium tree幼树Sapling幼苗Seedling耧斗菜叶绣线菊Spiraea aquilegifolia小叶锦鸡儿Caragana microphyllaPE东部Eastern87691173163PS南部Southern828047374PW西部Western657044855PN北部Northern1081210894111
1)成年树Adult tree: DBH≥20 cm; 中龄树Medium tree: 5 cm≤DBH<20 cm; 幼树Sapling: DBH<5 cm; 幼苗Seedling: 无胸径Without DBH.
总体上看,浑善达克沙地榆树种群的结构处于不稳定状态,幼苗不能正常生长发育,导致其幼树数量严重不足,甚至缺失,表现为衰退型种群;相较于北部、南部和西部3个样地,东部样地中榆树数量较多,种群结构较合理,而灌木数量则相对较少。
2.2 不同龄级榆树和主要灌木种类的空间格局
2.2.1 空间分布状况 浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类(耧斗菜叶绣线菊和小叶锦鸡儿)的空间分布状况见图1。结果显示:各样地的地形均有不同程度的起伏,可以明显区分出沙丘和丘间地;不同龄级榆树和主要灌木种类均呈现出不同程度聚集分布的空间格局。
2.2.2 基于CSR零模型的空间格局检验 采用CSR零模型对浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间格局进行检验,结果见图2。结果显示:在4个样地中,不同龄级榆树和主要灌木种类在小尺度上呈现显著的聚集分布特征,且随着空间尺度的增加,聚集程度逐渐下降,表现出随机分布的趋势,但在不同样地中不同龄级榆树和主要灌木种类的聚集尺度存在差异。
CSR零模型的检验结果(图2)显示:在东部样地中,成年榆树(DBH≥20 cm)、中龄榆树(5 cm≤DBH<20 cm)和榆树幼苗(无胸径)分别在0~2、0~3和0~8 m尺度内呈现显著的聚集分布。在南部样地中,成年榆树和榆树幼苗分别在0~5和0~3 m尺度内呈现显著的聚集分布,而耧斗菜叶绣线菊则在0~19 m尺度内呈现显著的聚集分布。在西部样地中,成年榆树和榆树幼苗分别在0~2和0~7 m尺度内呈现显著的聚集分布,而小叶锦鸡儿则在0~9 m尺度内呈现显著的聚集分布。在北部样地中,成年榆树和榆树幼苗分别在0~3和0~9 m尺度内呈现显著的聚集分布,而耧斗菜叶绣线菊则在0~6 m尺度内呈现显著的聚集分布。
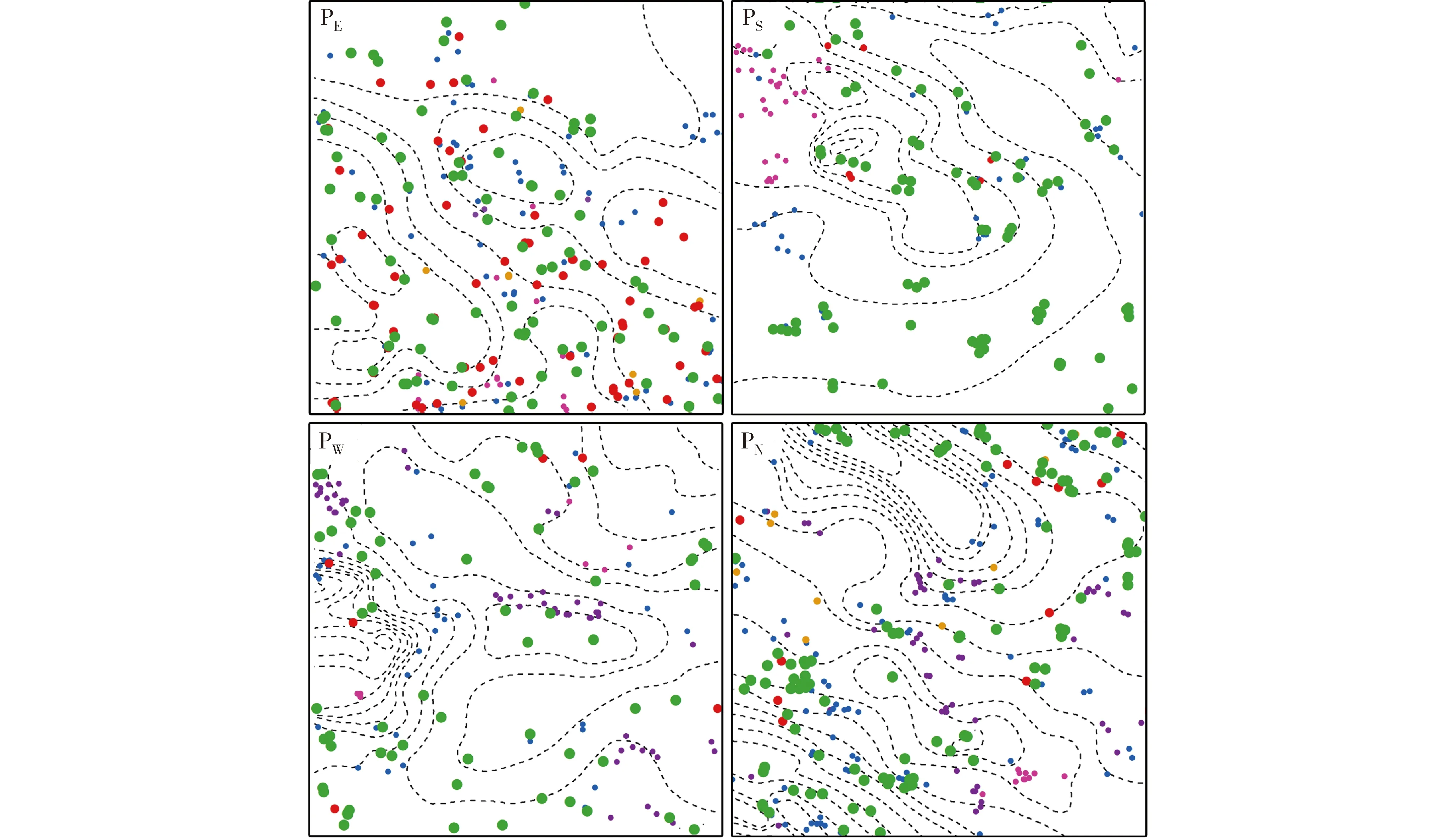
: 成年榆树Adult tree of Ulmus pumila Linn. (DBH≥20 cm); : 中龄榆树Medium tree of U. pumila (5 cm≤DBH<20 cm); : 榆树幼树Sapling of U. pumila (DBH<5 cm); : 榆树幼苗Seedling of U. pumila (无胸径Without DBH); : 耧斗菜叶绣线菊Spiraea aquilegifolia Pall.; : 小叶锦鸡儿Caragana microphylla Lam.; : 等高线Contour line.PE: 东部样地Eastern plot; PS: 南部样地Southern plot; PW: 西部样地Western plot; PN: 北部样地Northern plot.图1 浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间分布状况Fig. 1 Spatial distribution status of Ulmus pumila Linn. at different age classes and main shrub species in four plots of U. pumila-dominated savanna in Otindag Sandy Land
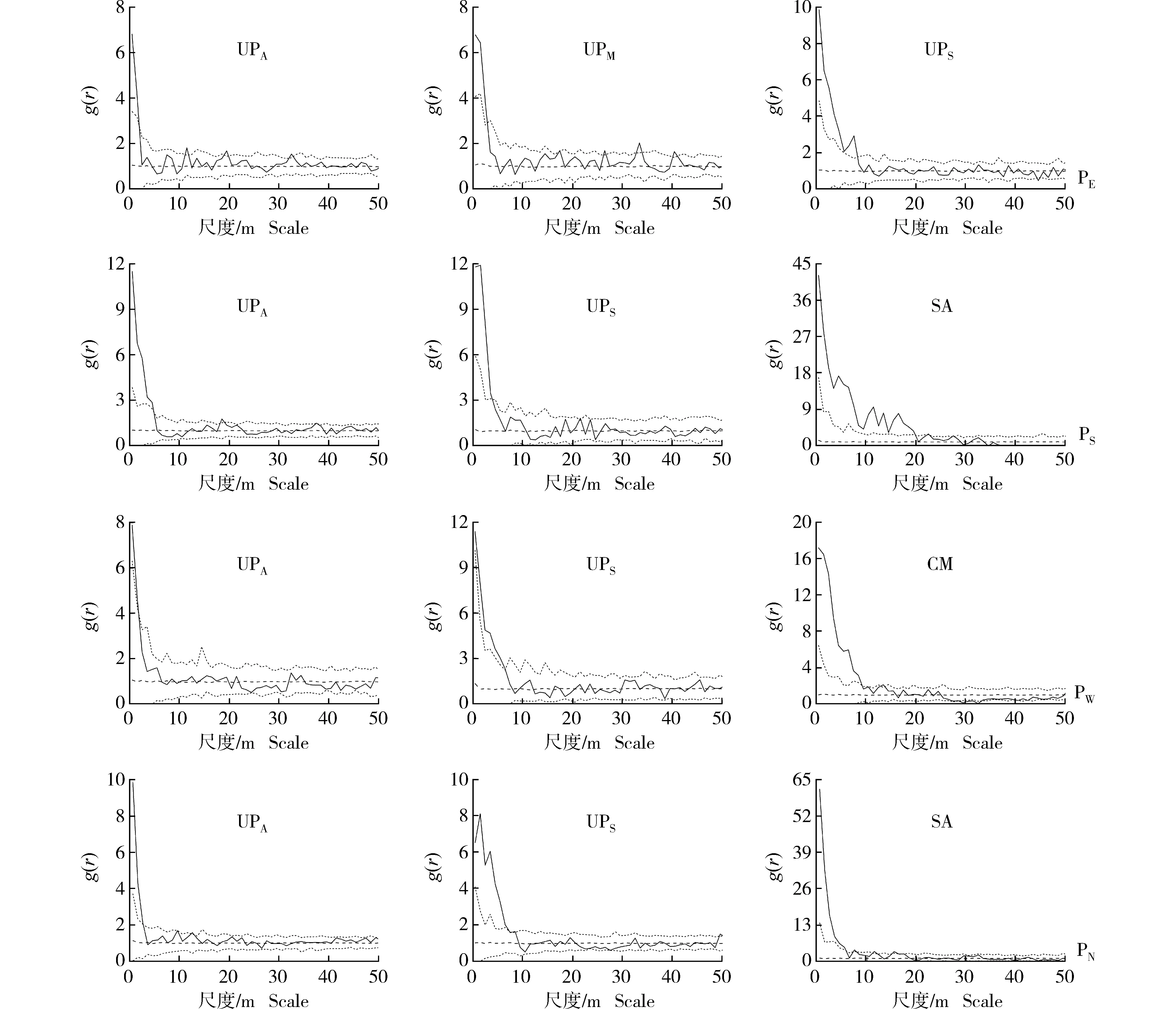
: 由实际数据计算得出的成对关联函数The paired correlation function calculated from the actual data; : 参考线Reference line; : Monte Carlo检验的置信区间(置信水平为99%)The confidence interval of Monte Carlo test (confidence level of 99%).UPA: 成年榆树Adult tree of Ulmus pumila Linn. (DBH≥20 cm); UPM: 中龄榆树Medium tree of U. pumila (5 cm≤DBH<20 cm); UPS: 榆树幼苗Seedling of U. pumila (无胸径Without DBH); SA: 耧斗菜叶绣线菊Spiraea aquilegifolia Pall.; CM: 小叶锦鸡儿Caragana microphylla Lam. PE: 东部样地Eastern plot; PS: 南部样地Southern plot; PW: 西部样地Western plot; PN: 北部样地Northern plot.图2 基于完全空间随机(CSR)零模型的浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间格局Fig. 2 Spatial pattern of Ulmus pumila Linn. at different age classes and main shrub species in four plots of U. pumila-dominated savanna in Otindag Sandy Land based on complete spatial randomness (CSR) null model
2.2.3 基于HP零模型的空间格局检验 采用HP零模型(排除生境异质性的影响)对浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间格局进行检验,结果见图3。结果显示:对比CSR零模型的检验结果,生境异质性对供试样地中各种类的空间分布特征有明显影响。在4个样地中,不同龄级榆树和主要灌木种类的聚集尺度均不同程度减小,对生境异质性的响应程度也不一致,部分龄级榆树和灌木的分布格局由小尺度上的聚集分布转变为所有尺度上的随机分布。
HP零模型的检验结果(图3)显示:在东部样地中,成年榆树、中龄榆树和榆树幼苗分别在0~1、0~2和0~4 m尺度内呈现显著的聚集分布。在南部样地中,成年榆树和榆树幼苗分别在0~3和1~2 m尺度内呈现显著的聚集分布,而耧斗菜叶绣线菊则在0~50 m尺度内呈现随机分布。在西部样地中,成年榆树和榆树幼苗均在0~50 m尺度内呈现随机分布,而小叶锦鸡儿则在0~3 m尺度内呈现显著的聚集分布。在北部样地中,成年榆树在0~2 m尺度内呈现显著的聚集分布,榆树幼苗则在0~50 m尺度内呈现随机分布,而耧斗菜叶绣线菊则在0~1 m尺度内呈现显著的聚集分布。
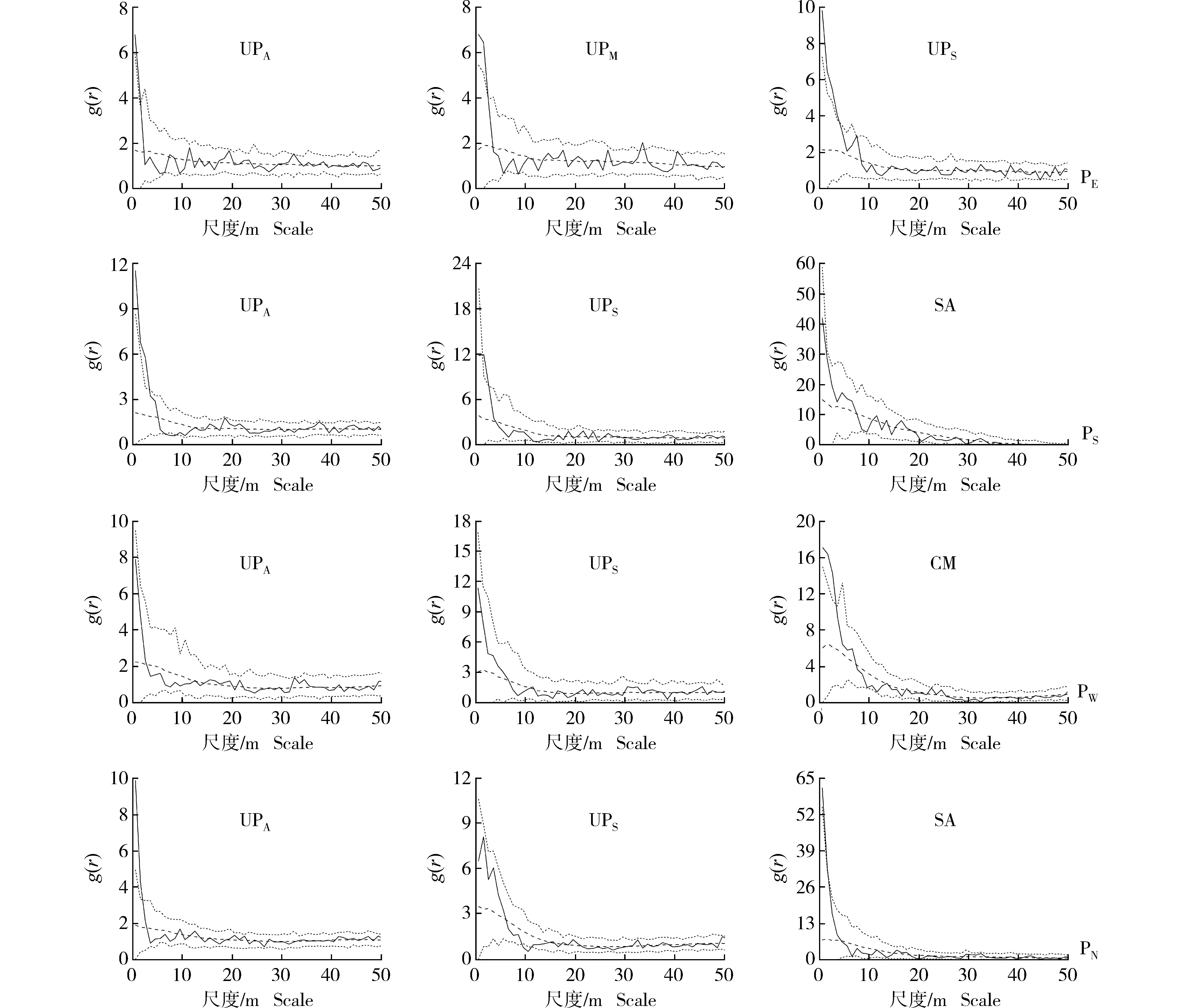
: 由实际数据计算得出的成对关联函数The paired correlation function calculated from the actual data; : 参考线Reference line; : Monte Carlo检验的置信区间(置信水平为99%)The confidence interval of Monte Carlo test (confidence level of 99%).UPA: 成年榆树Adult tree of Ulmus pumila Linn. (DBH≥20 cm); UPM: 中龄榆树Medium tree of U. pumila (5 cm≤DBH<20 cm); UPS: 榆树幼苗Seedling of U. pumila (无胸径Without DBH); SA: 耧斗菜叶绣线菊Spiraea aquilegifolia Pall.; CM: 小叶锦鸡儿Caragana microphylla Lam. PE: 东部样地Eastern plot; PS: 南部样地Southern plot; PW: 西部样地Western plot; PN: 北部样地Northern plot.图3 基于异质性泊松过程(HP)零模型的浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间格局Fig. 3 Spatial pattern of Ulmus pumila Linn. at different age classes and main shrub species in four plots of U. pumila-dominated savanna in Otindag Sandy Land based on heterogeneous poisson process (HP) null model
2.3 不同龄级榆树和主要灌木种类的空间关联性
采用CSR零模型和HP零模型分别对浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类(耧斗菜叶绣线菊和小叶锦鸡儿)的空间关联性进行检验,结果分别见图4和图5。
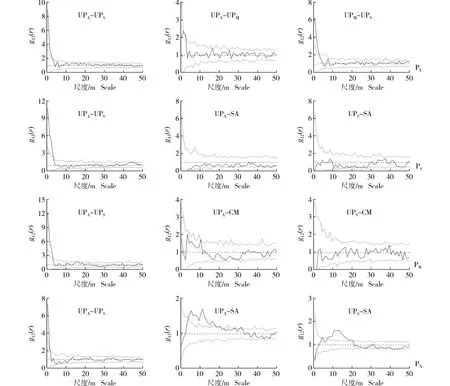
: 由实际数据计算得出的成对关联函数The paired correlation function calculated from the actual data; : 参考线Reference line; : Monte Carlo检验的置信区间(置信水平为99%)The confidence interval of Monte Carlo test (confidence level of 99%).UPA: 成年榆树Adult tree of Ulmus pumila Linn. (DBH≥20 cm); UPM: 中龄榆树Medium tree of U. pumila (5 cm≤DBH<20 cm); UPS: 榆树幼苗Seedling of U. pumila (无胸径Without DBH); SA: 耧斗菜叶绣线菊Spiraea aquilegifolia Pall.; CM: 小叶锦鸡儿Caragana microphylla Lam. PE: 东部样地Eastern plot; PS: 南部样地Southern plot; PW: 西部样地Western plot; PN: 北部样地Northern plot.图4 基于完全空间随机(CSR)零模型的浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间关联性Fig. 4 Spatial associations of Ulmus pumila Linn. at different age classes and main shrub species in four plots of U. pumila-dominated savanna in Otindag Sandy Land based on complete spatial randomness (CSR) null model
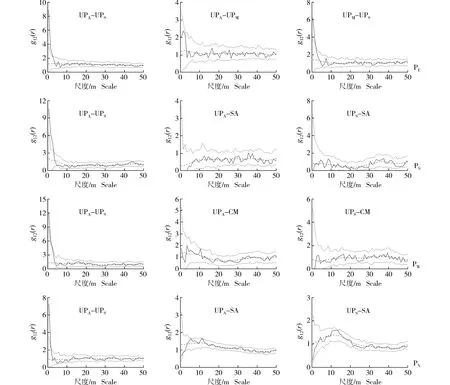
: 由实际数据计算得出的成对关联函数The paired correlation function calculated from the actual data; : 参考线Reference line; : Monte Carlo检验的置信区间(置信水平为99%)The confidence interval of Monte Carlo test (confidence level of 99%).UPA: 成年榆树Adult tree of Ulmus pumila Linn. (DBH≥20 cm); UPM: 中龄榆树Medium tree of U. pumila (5 cm≤DBH<20 cm); UPS: 榆树幼苗Seedling of U. pumila (无胸径Without DBH); SA: 耧斗菜叶绣线菊Spiraea aquilegifolia Pall.; CM: 小叶锦鸡儿Caragana microphylla Lam. PE: 东部样地Eastern plot; PS: 南部样地Southern plot; PW: 西部样地Western plot; PN: 北部样地Northern plot.图5 基于异质性泊松过程(HP)零模型的浑善达克沙地4个榆树疏林样地中不同龄级榆树和主要灌木种类的空间关联性Fig. 5 Spatial associations of Ulmus pumila Linn. at different age classes and main shrub species in four plots of U. pumila-dominated savanna in Otindag Sandy Land based on heterogeneous poisson process (HP) null model
由图4可见:不同龄级榆树以及主要灌木种类间的相互作用关系同样具有较强的尺度依赖性,其中,在东部、南部和西部3个样地中,成年榆树(DBH≥20 cm)与榆树幼苗(无胸径)在0~3 m尺度内呈现显著的正向作用关系;在北部样地中,成年榆树与榆树幼苗在0~2 m尺度内呈现显著的正向作用关系;且随着空间尺度的增加,4个样地中成年榆树与榆树幼苗的正向作用关系均逐渐降低,在绝大多数尺度上呈现相互独立的作用关系。此外,在东部样地中,成年榆树与中龄榆树(5 cm≤DBH<20 cm)在1~2 m尺度内呈现显著的正向作用关系,中龄榆树与榆树幼苗在0~3 m尺度内呈现显著的正向作用关系;在南部样地中,成年榆树和榆树幼苗分别与耧斗菜叶绣线菊在0~50 m尺度内呈现相互独立的作用关系;在西部样地中,成年榆树和榆树幼苗分别与小叶锦鸡儿在0~50 m尺度内呈现相互独立的作用关系;在北部样地中,成年榆树和榆树幼苗分别与耧斗菜叶绣线菊在4~17和7~20 m尺度内呈现显著的正向作用关系。
由图5可见:对比CSR零模型检验结果,在4个样地中,成年榆树与榆树幼苗间的关联性和作用尺度均没有发生变化,成年榆树与中龄榆树、耧斗菜叶绣线菊和小叶锦鸡儿间的关联性也没有发生变化,但是作用尺度发生了一定的变化。在东部样地中,成年榆树与中龄榆树在0~50 m尺度内呈现相互独立的作用关系,而中龄榆树与榆树幼苗则在0~2 m尺度内呈现显著的正向作用关系;在南部、西部和北部3个样地中,成年榆树和榆树幼苗分别与耧斗菜叶绣线菊和小叶锦鸡儿在0~50 m尺度内呈现相互独立的作用关系。
3 讨论和结论
种群结构是反映种群动态变化的重要指标,研究植物的种群结构可以了解其演变的历史,并预测其未来的发展趋势[29-30]。从20世纪90年代开始,浑善达克沙地中的榆树种群逐渐遭到毁坏,种群的结构完整性降低,呈现斑块状的分布特点[31]。本研究结果表明:浑善达克沙地榆树疏林中的榆树种群存在更新不良、出现断层的现象,幼苗在成长为幼树的过程中受到很大的干扰,导致幼树的数量严重不足。究其原因可能有以下2个方面:一方面,在土地利用方式改变和经济发展等多重压力下,过度放牧及人工周期性的机械割草严重制约了榆树幼苗的生长,跨年存活率降低[21];另一方面,在干旱、半干旱环境中,榆树的更新通常发生在降水较多的年份,且免受高温影响、水分充足的生境更有利于其更新,但浑善达克沙地严苛的水热条件阻碍了榆树的正常发育过程[32],同时,目前气候暖干化造成的草地沙化也增加了榆树的更新难度[33]。
自然界中的物种大多呈聚集分布的状态;通常认为,物种在一定尺度上呈聚集分布有利于通过集群效应抵抗不良环境的影响,并在适应环境的过程中对环境进行定向改造,从而促进种群的稳定和繁衍[26]。本研究结果支持物种在特定尺度上呈聚集分布的观点。而多个不同的生态过程会导致物种的聚集分布,这是空间格局具有尺度依赖性的理论基础[9]。种子繁殖是榆树、耧斗菜叶绣线菊和小叶锦鸡儿的重要繁殖方式[34-36],种子成熟后会在重力的作用下飘落到地面,以母树为中心形成聚集分布状态,或者在风力、地面径流以及地面植物阻截等外力作用下形成聚集状态。生境异质性是干旱、半干旱环境中重要的地貌特征。土壤和光照等资源在空间上呈现斑块状分布,资源良好的生境为种子萌发及植物定植创造了有利条件,从而导致物种的聚集分布[6]。本研究结果显示:在排除生境异质性后,所有物种的聚集尺度明显降低,证实了生境异质性对物种的聚集分布影响较大。此外,浑善达克沙地严苛的环境条件还促使物种(尤其是长势较差和地上部死亡的榆树)具有明显的萌蘖繁殖现象,而萌蘖繁殖会在小尺度上造成物种的聚集分布。综上所述,种子扩散限制、萌蘖繁殖以及生境异质性可能是形成榆树疏林中主要木本植物呈聚集分布的重要原因。
种群内部植物个体间的相互关系是影响种群更新的重要因子[37]。本研究中,在2种零模型检验下,不同龄级榆树在小尺度上均呈现显著的正向作用关系,表明成年榆树与榆树幼苗是互相促进的关系,成年榆树可以为其种子萌发和幼苗生长提供适宜的微环境,二者间是“庇护与被庇护”的关系。由于“庇护植物”冠下具备更多的可用水量、更高的土壤养分、更低的放牧强度以及良好的遮光环境,因而,“庇护植物”以其固定的树冠持续为植物种子萌发和幼苗存活提供良好的生境条件[38]。
群落中不同物种间的相互作用是促进群落演替的主要驱动力[39-40]。不同龄级榆树分别与耧斗菜叶绣线菊和小叶锦鸡儿在所有尺度上均呈现相互独立的作用关系,表明浑善达克沙地榆树疏林中各龄级榆树与主要灌木种类在空间上各自占据不同位置,具有不同的生态位,对土壤、水分和光照等环境资源的利用各取所需,未产生直接竞争。
值得注意的是,在目前情况下,虽然浑善达克沙地榆树疏林中灌木的存在未对榆树的生长造成直接影响,但通过对比不同样地间(处于不同水热条件下)的植被组成情况[41-42],可以推断未来如果气候干旱加重,榆树的数量可能持续减少,而灌木植物(如耐旱植物小叶锦鸡儿等)的数量则可能持续增加,灌木种类将有可能最终占据整个疏林草地系统,以榆树为主导的疏林草地可能会逐步退化为以灌丛为主导的疏林草地,从而引起整个疏林草地系统的退化。因此,建议在榆树种群衰退严重的区域,通过建立自然保护区的形式,强化幼苗的生长和抚育,以期恢复该区域内整个榆树疏林系统的活力。
猜你喜欢
河南畜牧兽医(2021年7期)2021-12-10
阅读(中年级)(2021年10期)2021-03-09
诗潮(2019年10期)2019-11-19
阅读与作文(英语高中版)(2019年9期)2019-11-07
故事作文·低年级(2019年10期)2019-10-16
扬子江(2019年3期)2019-05-24
读友·少年文学(清雅版)(2019年12期)2019-04-20
诗潮(2017年2期)2017-03-16
小学阅读指南·高年级版(2009年7期)2009-10-09
小猕猴学习画刊(2006年2期)2006-01-11
