
不同降雨年型下反枝苋和大豆光合特征的比较
2019-09-10 07:22孙彦坤陈睿李静鲁萍李琦白雅梅张险峰肖同玉李亦奇
江苏农业学报 2019年3期
孙彦坤 陈睿 李静 鲁萍 李琦 白雅梅 张险峰 肖同玉 李亦奇





摘要: 为揭示不同降雨年型下C4外来杂草与C3作物的竞争机制,以C4外来杂草反枝苋和C3作物大豆为研究对象,利用盆栽试验,设置3种降雨年型(高雨量、正常雨量和低雨量),比较单种和混种模式下两种植物的气体交换参数、光合色素含量和比叶面积的差异。结果显示,在相同降雨年型和栽培模式下,反枝苋的净光合速率、气孔导度、蒸腾速率及光合水分利用效率均高于大豆;反枝苋在三种降雨年型苗期均保持较高的净光合速率,不受降雨年型影响,但大豆净光合速率受降雨年型的影响显著;在低雨量年型下反枝苋光合水分利用效率最大,大豆光合水分利用效率最小。两种植物的这些差别很可能是反枝苋成功入侵东北大豆田的重要原因。
关键词:反枝苋;大豆;降雨年型;光合特性;叶绿素含量;比叶面积
中图分类号:S459
文献标识码:A
文章编号: 1000-4440(2019)03-0554-10
人类进入工业文明后,大量化石燃料的使用使大气中温室气体浓度升高,加剧温室效应,地球表面温度的变化也使降雨格局发生变化,主要表现为降雨总量的改变、单次降雨的波动以及降雨时间间隔的变化[1]。已有研究结果表明,降雨格局变化对C,和C4植物的生长动态具有不同影响[2-5]。Monson等发现干旱的冬天和潮湿的夏天促进C4植物的生长,而潮湿的冬天和干旱的夏天则促进C3植物的生长[6]。
据统计,全世界18种恶性杂草中有14种是C4植物,86种粮食作物中81种是C3植物[7]。尽管关于C。杂草和C,作物之间相互作用的研究报道有很多[8-11],但比较C4外来杂草和C3本地作物光合特性对降雨格局变化响应机制的研究还甚少[12]。
反枝苋(Amaranthus retrofiexus)是原产地为美洲的一种典型C4杂草,常分布于道路两旁、农田和荒野[13]。反枝苋的入侵使大豆(Glycine max)、玉米(Zea mays)、小麦(Triticum aestivum)等作物的产量明显减少[13]。反枝苋可以入侵到许多C3作物的农田,大豆是原产于中国东北的重要C3作物,大豆田也是反枝苋经常入侵的农田[13]。
近年来,关于不同降雨格局影响农业生态系统的研究主要针对作物,且主要是研究不同降雨年型下作物的出苗率、水分利用效率、生长速率、产量和栽培优化模式[14-16]。目前为止,针对降雨年型变化下外来杂草和作物竞争机制的研究却不多[17]。
本试验以C4外来杂草反枝苋和C3本地作物大豆为研究材料,利用盆栽试验的方法,比较不同降雨年型和竞争模式下两种植物的气体交换参数、光合色素含量和比叶面积的变化规律,探索两种植物对降雨格局变化的响应及竞争机制,为揭示外来杂草在全球气候变化背景下在农田中的入侵过程提供理论参考。
1 材料与方法
1.1 试验地点和材料
2016年在东北农业大学塑料大棚内进行盆栽试验,试验地为寒温带大陆性气候,无霜期大气温度平均值是3.78℃,全年降雨总量为400 - 600mm[18]。盆栽所用黑土的理化性质为:有机质含量53.53 g/kg,全氮含量5.50 g/kg,全磷含量1.03g/kg,速效钾含量162.26 mg/kg,碱解氮含量173.13 mg/kg,速效磷含量34.67 mg/kg,土壤pH值7.35。
盆栽试验的塑料盆半径为14 cm,高度为21cm,塑料盆的左右两侧各有3个孔(孔直径0.5 cm,距离盆底2 cm),盆底放有直径为29 cm、高度为3cm的托盘(以便在浇水量大时,收集从盆底溢出的水,待盆内土壤中水蒸发后再归还盆中,以保证每次试验处理的准确性),每盆装土8.3 kg。在黑土中施入尿素(氮含量46. 4%)、过磷酸钙(P2 05,50%)、硫酸钾( K70,50%),施用水平均为50 kg/hm2,经计算每盆氮、磷、鉀肥施用量分别为0.664 g、1.925g、0.616 g。
反枝苋种子2015年秋采于哈尔滨市香坊农场,大豆品种为东农62号。
1.2 试验设计
试验为双因素(降雨年型和竞争模式)随机区组设计。3种降雨年型为高雨量年型、正常雨量年型和低雨量年型。其中正常年型的降雨量为415mm,是近30年来哈尔滨市6月至9月降雨量的平均值,约为全年降雨量的80%(黑龙江省气象台资料);高雨量年型的降雨量为498 mm,高于正常年型20%;低雨量年型的降雨量为322 mm,低于正常年型20%。2种竞争模式:种内竞争,即每盆种植相同植物2株,2株大豆或2株反枝苋;种间竞争,即每盆种植2种植物各1株,1株大豆和1株反枝苋。试验共设置4个区组,每个区组包含每个处理7个重复。
通过人工浇水模拟不同降雨年型。试验于6月1日开始,9月28日结束,持续时间为120 d,每隔ld浇水1次。依据正常年型的降雨量变化规律计算每次降雨量,降雨量变化规律采用一元二次方程y= -0. 007 lx2+0. 431 9x+2. 515 2模拟,x为浇水累计次数,y为降雨量。然后用降雨量乘以盆上表面积得到每次的浇水量,准确称量后倒入塑料喷壶中模拟降雨。高雨量年型处理每次浇水量比正常年型多20%,低雨量年型处理每次浇水量比正常年型少20%,降雨模拟见图1。为减少水蒸发量与周围环境中水蒸发量的差别,试验在透光性良好的塑料薄膜遮雨棚中进行,四周塑料薄膜卷起1.5 m高以保证通风状况良好。盆栽试验播种时间为2016年5月20日,10 d后按试验设计定苗。
1.3 测定指标与方法
1.3.1 气体交换参数的测定2016年6月23-29日、7月23-31日、8月24-31日,在天气晴朗的上午8:00-11:00,每个处理取4盆植物,随机选取健康植株顶部成熟叶片,用Ciras-2便携式光合仪测定气体交换参数。测定时设置光合仪的叶室光照度、CO,浓度、温度、空气相对湿度分别为1700μmol/( m2.s)、380 μmol/mol、25℃和70%。测定反枝苋和大豆叶片的净光合速率(Pn)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(E),并计算光合水分利用效率(WUE),WUE=净光合速率(Pn)/蒸腾速率(E)。
1.3.2 光合色素含量的测定2016年6月24日、7月24日、8月24日,测定两种植物的光合色素含量,每个处理测定4盆植物,4次重复。采集植株顶部完全展开的第1片叶(大豆称取0.2g,反枝苋称取0.3g),参照李合生等的方法[19]测定叶绿素含量。
1.3.3 比叶面积的测定 2016年6月24日、7月24日、8月24日,每个处理取4盆植物,将每株植物的叶片全部摘下放人封口袋中,带回实验室,用Li-3100C台式叶面积仪测定植株总叶面积,在65 cC电热鼓风干燥箱中烘干后称质量。比叶面积计算公式:比叶面积=叶片面积/叶片干质量[20]。
1.4 数据分析
采用SPSS 19.0统计分析软件分析试验数据。通过三因素重复测量方差分析,检验降雨年型、竞争模式、生长时期和它们的交互作用对两种植物的气体交换参数、光合色素含量、比叶面积的影响显著性;用Duncan氏多重比较方法,分析反枝苋和大豆的各项参数在相同测定时期不同降雨年型或相同降雨年型不同测定时期之间各项指标的差异;通过独立样本t检验,分析反枝苋与大豆的各项参数在相同降雨年型和相同竞争模式下的差异。采用OriginPr0 8.5绘图软件作图。
2 结果与分析
2.1 反枝苋和大豆净光合速率对不同降雨年型的响应
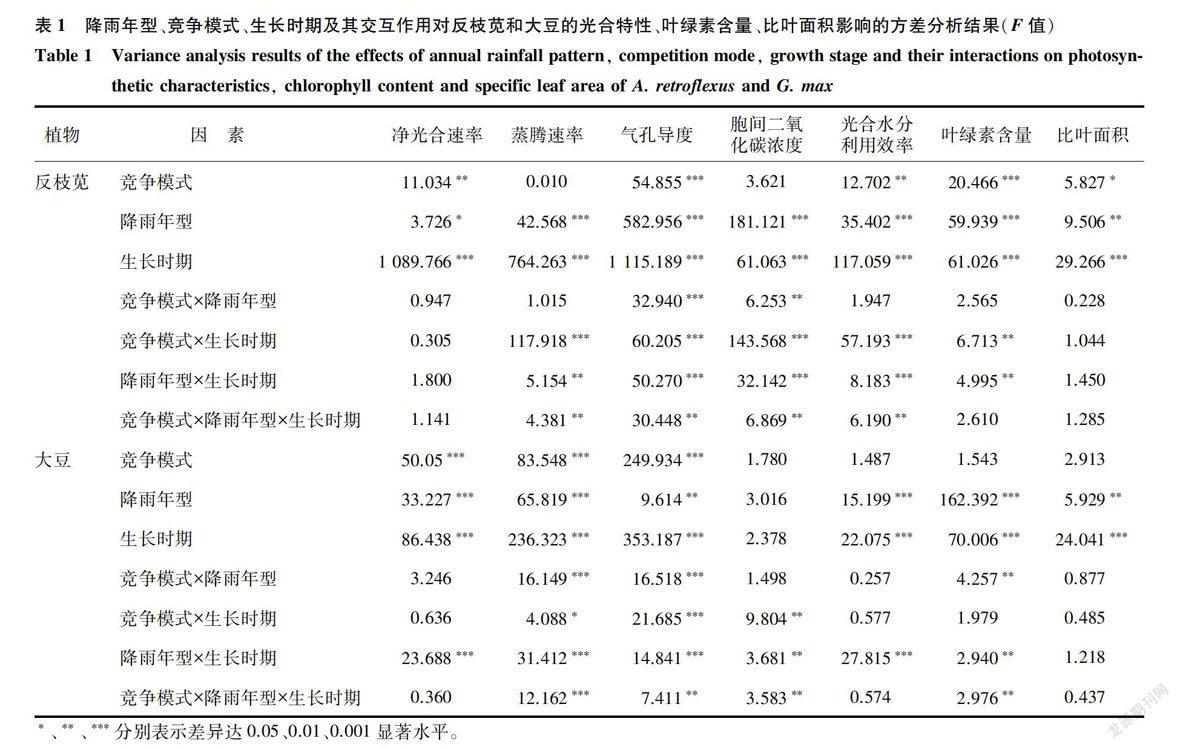
图2、表1表明,降雨年型和竞争模式对反枝苋和大豆的净光合速率(Pn)均有显著影响(P<0.05)。大豆的净光合速率均以高雨量年型为最大,低雨量年型最小(除7月开花结荚期外);降雨年型对苗期(6月)反枝苋净光合速率的影响不大,但在7月混种条件下和8月单种条件下影响显著。种间竞争显著降低大豆的Pn,混种大豆Pn小于单种大豆;种间竞争促进反枝苋的Pn,混种反枝苋Pn大于单神反枝苋。两种植物Pn的最大值均出现在生长前期,且全生育期内反枝苋的Pn均远大于大豆的Pn(除8月份单种高雨量年型外)(P<0.001)。
2.2 反枝苋和大豆蒸腾速率对不同降雨年型的响应
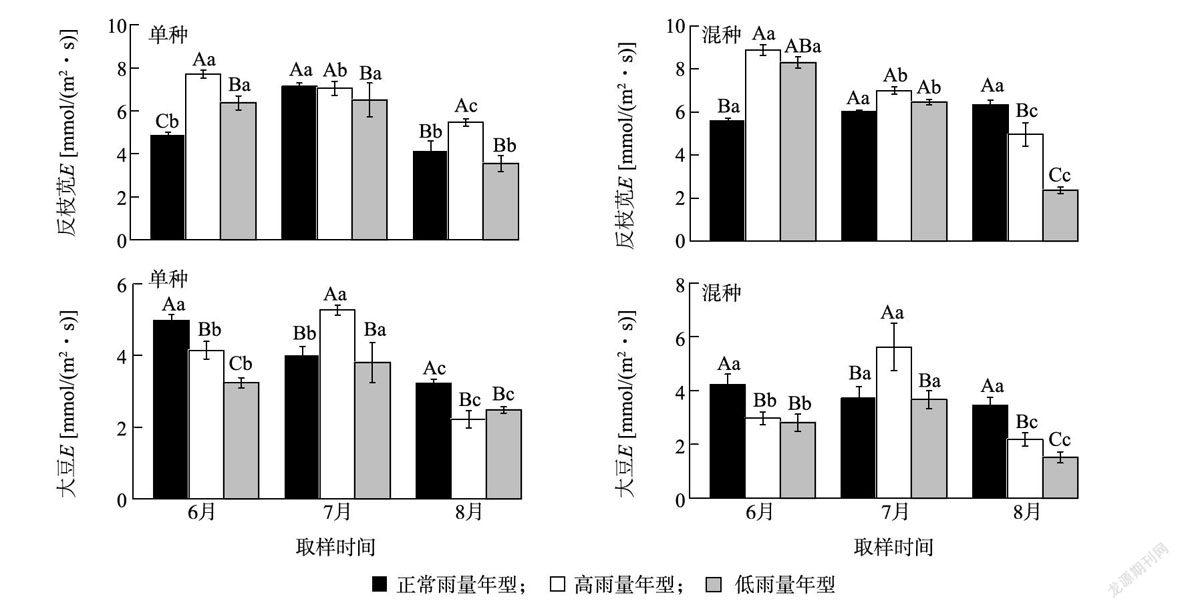
降雨年型对反枝苋和大豆蒸腾速率均有显著影响( P
2.3 反枝苋和大豆气孔导度对不同降雨年型的响应
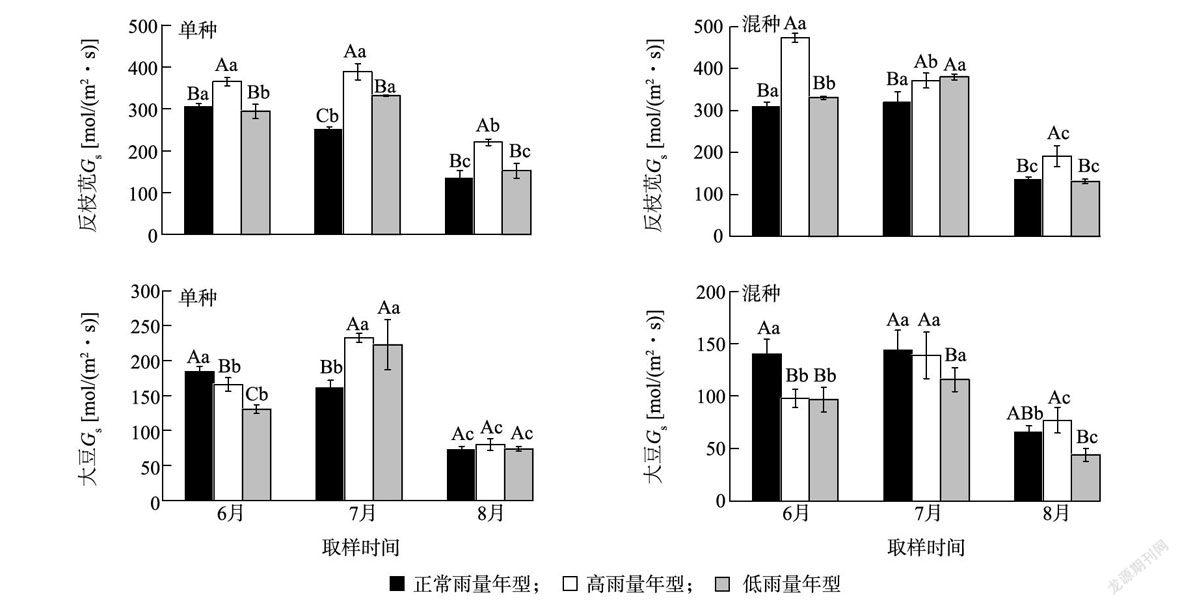
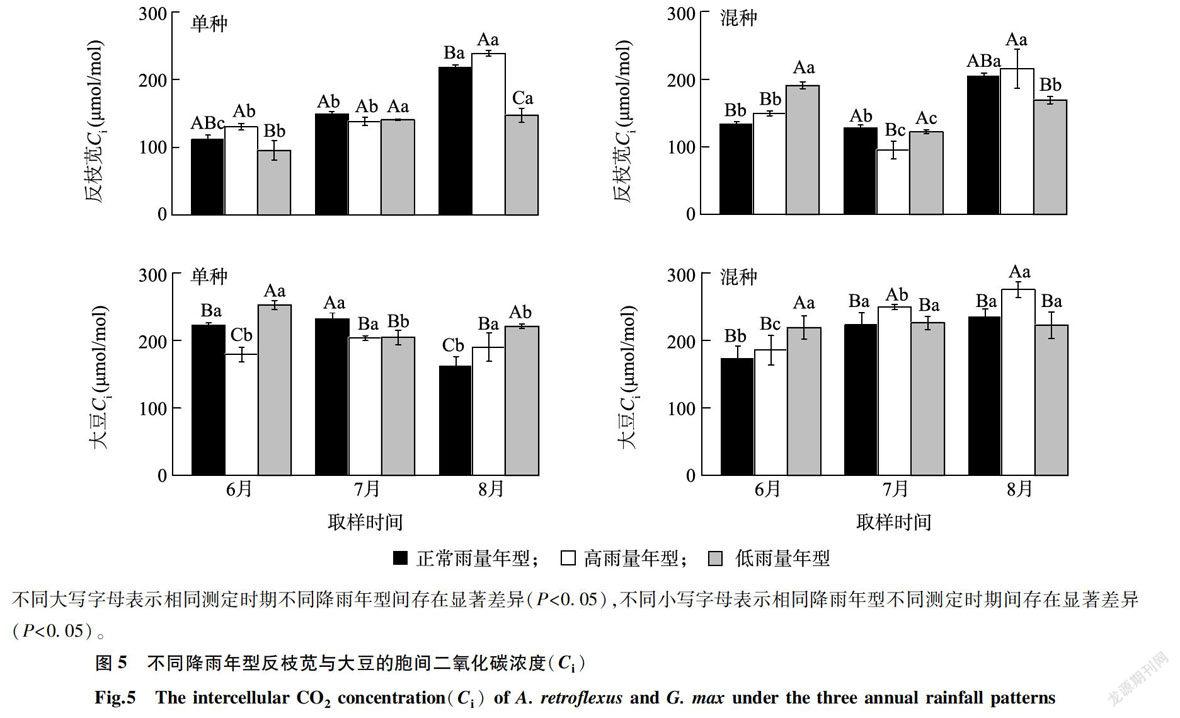
图4、表l表明,降雨年型和竞争模式对反枝苋和大豆气孔导度(Gs)的影响显著(P 2.4 反枝苋和大豆胞间二氧化碳浓度对不同降雨年型的响应 降雨年型对反枝苋胞间二氧化碳浓度(Ci)的影响显著(P 图6、表1表明,降雨年型对反枝苋和大豆光合水分利用效率(WUE)的影响显著(P 降雨年型对反枝苋和大豆叶片叶绿素含量的影响显著( P
2.7 反枝苋和大豆比叶面积对不同降雨年型的响应
图8、表1表明,降雨年型对反枝苋和大豆的比叶面积有显著影响(P<0.01)。两物种的比叶面积均在高雨量年型最大,在低雨量年型最小(除8月混种反枝苋及大豆外)(P<0.05)。竞争模式对大豆比叶面积的影响不显著(P>0.05),对反枝苋比叶面积的影响显著(P<0.05),混种反枝苋比叶面积显著高于单种反枝苋(P<0.05)。在相同降雨年型和生长时期大豆比叶面积显著高于反枝苋(除7月份混种高雨量年型外)( P 3 讨论 3.1 不同降水年型反枝苋与大豆气体交换参数的比较 在植物气体交换参数中最关键的参数是净光合速率(Pn),具有较强的光合能力也是植物在竞争中获得优势的一个关键特性[21,22]。很多外来入侵植物成功入侵的主要原因之一就是其具有比本地植物更高的光合能力[23]。Feng等研究结果表明,外来人侵植物红花酢浆草(Oxalis corymbosa)和草胡椒(Peperomia pellucida)比同属本地种具有更高的Pn、Gs、E及WUE[24]。陈新微等研究结果表明,入侵植物银胶菊(Parthenium hysterophorus)、三叶鬼针草(Bidens pilosa)比本地伴生植物小蓟(Cirsiumsetosum)具有更高的Pn、Gs、E及WUE[22]。与他们的研究结果相似,本试验结果显示在每一生长时期,反枝苋的Pn、Gs、E及WUE均大于大豆(除8月单种反枝苋小于大豆外),尤其是在生长前期,反枝苋Pn值为大豆的约2-5倍,且反枝苋的气体交换参数受降雨年型的影响不显著,在低雨量年型,仍具有较高的光合能力,但大豆则不能在干旱的条件下保持较高的光合能力;并且反枝苋WUE在低雨量年型最大,大豆WUE在低雨量年型最小。由此可见,反枝苋作为在全世界具有广大分布区的C4型杂草[25],对降雨波动的适应能力强,而C3型大豆對降雨波动的适应能力较弱,这可能是反枝苋入侵中国东北大豆田的重要机制之一。 本試验结果还显示,在整个生长过程中大豆的Ci均大于反枝苋,这与王健林等[26]的研究结果相似,即C3植物可以通过高浓度的Ci快速补充C02,而C4植物反枝苋的独特光合过程可以使其在低CO2浓度下进行光合作用。光合作用是植物生长和干物质积累的关键机制,干旱胁迫下植物调节自身的Gs和E以减少水分流失,使细胞中的渗透压发生变化及Ci降低,减少进入叶绿体的C02,降低光合速率[27]。两种植物的E与Gs值在生长前期与生长后期的最小值均出现在低雨量年型下,说明两种植物均可通过调节气孔闭合程度减少蒸腾来适应降雨欠缺的条件。 竞争对大豆的Pn、Gs、E和反枝苋的Pn、Gs和WUE的影响显著,且竞争对两物种Pn的影响不随降雨年型和生长时期的改变而改变(交互作用不显著)。当两者混种时,混种反枝苋的Pn和Gs显著大于单种反枝苋,而混种大豆的Pn和Gs显著小于单种大豆,说明种间竞争有利于反枝苋生长和保持较高的光合速率,却降低大豆的光合作用,减弱其生长,反枝苋在种间竞争中占有优势,故而容易入侵成功。 3.2 不同降雨年型反枝苋与大豆叶片光合色素含量的比较 叶绿素参与植物的光吸收、传导和转化,是植物光合作用过程中关键的构成部分。叶绿素含量的高低会影响植物光合作用的强弱[28-29]。以往关于降雨和水分波动对大豆叶片叶绿素含量影响的研究结果不一。有研究者发现干旱对大豆叶片叶绿素含量无显著影响[30]。也有研究者发现干旱降低大豆叶片叶绿素含量,且影响效果随着大豆品种不同而不同[31]。还有研究者发现降雨量对大豆叶片叶绿素含量的影响在苗期和开花结荚期不明显,但鼓粒期影响显著[29]。本试验发现,大豆叶片叶绿素含量在高雨量年型最大,大豆Pn值也在高雨量年型最大,尤以苗期显著。张茜等研究发现,降雨季节波动对反枝苋叶片叶绿素含量的影响较大[29]。本试验结果显示反枝苋叶片叶绿素含量在高雨量年型最大,低雨量年型最小,受降雨年型的影响显著。 3.3不 同降雨年型反枝苋与大豆比叶面积的比较 入侵植物的生物学特性在很大程度上决定了其入侵性,其中一个重要特征是比叶面积较大[24]。相对于本地植物,一些入侵植物往往具有更大的比叶面积[32-35]。具有较大比叶面积的植物可更好地捕获光能,在光照弱的情况下有更好的适应能力[26]。潘玉梅等研究发现,低光照下鬼针草属的外来入侵植物的比叶面积大于本地植物,但高光照下差别不大[35]。本研究结果显示,在相同降雨年型和竞争模式下,大豆的比叶面积大于反枝苋,与上述结果不一致。这可能是由于本研究中两物种对光能的竞争策略不同,大豆倾向于形成较大的叶面积,而反枝苋倾向于通过较高的株高获得优势[36-37]。这表明相对于比叶面积的大小,植株高度对反枝苋的入侵性更重要。 总之,不管处于何种降雨年型和竞争模式下,C4型入侵植物反枝苋均比C3型本地作物大豆具有更高的Pn、Gs、E及WUE;在3种降雨年型下,反枝苋在苗期均具有较高的Pn,不受降雨年型的影响,但大豆受降雨年型的影响显著;在低雨量年型下,反枝苋的WUE最大,大豆的WUE最小。两物种的这些差别很可能是反枝苋能够成功入侵东北大豆田的重要原因。 参考文献 [l] PACHAURI R K, REISINCER A . Climate change 2007:Synthesis report [ R ] .Spain : Valencia , 2007. [2] PARUELO J M. LAUENROTH W K. Relative abundance of plantfunctional types in grasslands and shrublands of North America[J] . Ecologcal Applications , 1996.6 : 1212-1224. [3] PARUELO J M, JOBBACY E G, SALA O E, et al. Functionaland structural convergence of temperate grassland and shrubland e-cosystems [J] . Ecological Applications , 1998 .8 : 194-206. [4] WINSLOW J C, HUNT J E R, PIPER S C. The influence of sea-sonal water availability on global C3 versus C4 grassland biomassand its implications for climate change research [J] . EcologicalModeling,2003 , 163 : 153-173. [5] NIU S L, YUAN Z Y, ZHANG Y F. et al. Photosynthetic respon-ses of C3 and C4 species to seasonal water variability and competi-tion[J].Joumal of Experimental Botany, 2005, 56( 421):2867-2876, [6] MONSON R K, LITTLEJOHN R 0, WILLIAMS III G J.Photo-synthetic adaptation to temperature in four species from theColorado shortgrass steppe:a physiological model for coexistence[J]. Oecologia, 1983,58:43-51. [7]PATTERSON D T.Weeds in a changing climate[J].Weed Sci-ence,1995 ,43:685-701. [8]BUNCE J A.Long-term growth of alfalfa and orchard grass plots atelevated carbon dioxide[J].Journal of Biogeochemistry, 1995,22:341-348. [9]POTVIN C, VASSEUR L. Long-term C02 enrichment of a pasturecommunity[J].Ecology,1997,78:666-677. [10] ZISKA L H.The impact of elevated C02 0n yield loss from a C3and C4weed in field-grown soybean[J].Global Change Biology,2000,6:899-905. [11]魯萍,金成功,张茜,等,反枝苋和大豆对降雨季节波动的生理生态响应[J].作物杂志,2017(2):114-120. [12]张茜.降雨波动对不同栽培模式下反枝苋光合特性的影响[D].哈尔滨:东北农业大学,2016. [13]李晓晶,张宏军,倪汉文.反枝苋的生物学特征及防治[J].农药科学与管理,2004,25(3):13-16. [14]陈光斗,赵万春,安成立,不同降水年份渭北不同降水年份渭北旱塬小麦高产栽培优化模式研究[J].干旱地区农业研究,1995,13(3):8-14. [15]张云兰,王龙吕,周聪明,等.高温伏旱区旱地农作系统水分供需平衡特征与生态适应性研究[J].水土保持研究,2010,17(6):95-105. [16]冯良山,孙占祥,肖继兵,等.不同微集水方式在不同降水年型对玉米产量的影响[J].东北农业大学学报,2011,42(1): 50-54. [17]丛雪,吴岩,鲁萍,等.氮素波动对反枝苋和大豆最大净光合速率和光合氮利用效率的影响[J].作物杂志,2013(1): 73-77. [18]徐广惠,王宏燕,刘佳,抗草甘膦转基因大豆(RRS)对根际土壤细菌数量和多样性的影响[J].生态学报,2009,29(8): 4535-4541. [19]李合生,孙群,赵世杰,等.植物生理生化实验原理和技术[M]北京:高等教育出版社,2000. [20]王满莲,冯玉龙.紫茎泽兰和飞机草的形态、生物量分配和光合特性对氮营养的响应[J].植物生态学报,2005,29(5): 697 -705. [21]王康满,侯元同,山东归化植物一新记录属——银胶菊属[J].曲阜师范大学学报(自然科学版),2004,30(1):83-84. [22]陈新微,李慧燕,刘红梅,等,入侵种银胶菊和三叶鬼针草与本地种气体交换特性的比较[J].生态学报,2016,36( 18):5732-5740. [23] POOTER L Light-dependent changes in biomass allocation and their importance for growth of rain forest tree species[J].Functional Ecology, 2001, 15(1):113-123. [24]FENG Y L,FU G L,ZHENC Y L.Specific leaf area relates to thedifferences in leaf construction cost, photosynthesis, nitrogen allo-cation. and use efficiencies between invasive and noninvasive aliencongeners[J].Planta, 2008, 228: 383-390. [25]鲁萍,梁慧,王宏燕,等,外来入侵杂草反枝苋的研究进展[J].生态学杂志,2010,29(8):1662-1670. [26]王建林,杨新民.气孔导度和叶片内部导度制约C3和C4作物光合作用的比较分析[J].东北农业大学学报,2011,42(1): 129-132. [27]RIZWAN Z,HAORAN D, MUHAMMAD A, et al.Potassium fer-tilizer improves drought stress alleviation potential in cotton bv en-hancing photosynthesis and carbohydrate metabolism[J].Environ-mental and Experimental Botany, 2017, 137: 73-83. [28]张云海,何念鹏,张光明,等.氮沉降强度和频率对羊草叶绿素含量的影响[J].生态学报,2013 ,33( 21):6786-6794. [29]张茜,金成功,鲁萍,等,降雨季节波动对反枝苋与大豆光合色素的影响[J].作物杂志,2016(1):154-161. [30]乔振江,蔡昆争,骆世明.低磷和干旱胁迫对大豆植株干物质积累及磷效率的影响[J].生态学报,2011,31( 19):5578-5587. [31]董守坤,赵坤,刘丽君,等,干旱胁迫对春大豆叶绿素含量和根系活力的影响[J].大豆科学,2011. 30(6):949-953. [32] CROTKOPP E,REJMA'NEK M. ROST T L Toward a causal ex-planation of plant invasiveness: seedling growth and life-historvstrategies of 29 pine( Pinus) species[J].American Naturalist,2002 ,159:396-419. [33] DENG X, YE W H, FENG H L,et al.Cas exchange characteris-tics of the invasive species Mikania micrantha and its indigenouscongener M.cordata( Asteraceae) in South China[J].BotanicalBuLletin of Academia Sinica, 2004 .45: 213-220. [34] BURNS J H.Relatedness and environment affect traits associatedwith invasive and noninvasive introduced Commelinaceae[J].Eco-logical Applications, 2006, 16: 1367-1376. [35]潘玉梅,唐賽春,韦春强,等.不同光照和水分条件下鬼针草属入侵种与本地种生长、光合特征及表型可塑性的比较[J].生物多样性,2017,25( 12):1257-1266. [36] MA X Y. WU H W, JIANG W L,et al-Interference between re-droot pigweed (Amaranthus retroflexus L.)and cotton( Gossypiumhirsutum L.): Growth analysis[J].PLoS ONE, 2015, 10(6):e0130475. [37] LU P,LI J X. JIN CG,et al.Different growth responses of an in-vasive weed and a native crop to nitrogen pulse and competition[J]. PLoS ONE ,2016,11(6):e0156285. (责任编辑:张震林)
猜你喜欢
农民致富之友(2018年9期)2018-06-27
农产品市场周刊(2017年6期)2017-03-10
农产品市场周刊(2017年4期)2017-03-03
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
儿童故事画报·智力大王(2015年10期)2016-01-27
幼儿智力世界(2015年5期)2015-08-20
