
穗发芽程度对粳稻稻米品质和蛋白质组分的影响
2019-09-10 07:22:44裘实卫平洋魏海燕葛佳琳韩超,郭保徐栋胡蕾吴培张军贺文畅张洪程
江苏农业学报 2019年3期
裘实 卫平洋 魏海燕 葛佳琳 韩超, 郭保 徐栋 胡蕾吴培 张军 贺文畅 张洪程

摘要:本研究以南粳9108、金粳818和常软06-2为试验材料,人工模拟水稻生育后期穗发芽条件,设置了破胸露白(处理一)和芽长3-5 cm(处理二)2个发芽水平,以3个粳稻品种未作处理的样品作为对照(CK),研究穗发芽对稻米品质相关指标的影响。结果表明:(1)稻谷发芽显著影响稻米品质,特别是软米品种南粳9108,容易穗发芽且发芽后稻米品质劣变严重,其中,稻米加工品质随穗发芽程度加深显著下降。(2)外观品质随着穗发芽程度加深而显著变劣,同时,粒长、粒宽和长宽比均显著下降。(3)蒸煮食味品质和营养品质随穗发芽程度加深而降低,稻米直链淀粉含量和蛋白质含量呈下降趋势而胶稠度呈现破胸前先上升而后下降的趋势。食味值随稻谷发芽程度加深而降低,这与加工品质和外观品质的变化趋势一致。各处理的RVA谱特征值中,峰值黏度、热浆黏度、崩解值和最终黏度均呈下降趋势。(4)各处理蛋白质组分中,随着穗发芽程度的增加,谷蛋白含量降低,醇溶蛋白含量增加,籽粒谷醇比呈现下降趋势。这说明穗发芽对籽粒淀粉结构和蛋白质结构的劣化破坏,大幅度降低了优质食味稻米的稻米品质,对稻米的优质生产造成了严重的危害。
关键词:穗发芽;优质稻米;稻米品质;RVA;蛋白组分分析
中图分类号: S511.2+2
文献标识码: A
文章编号: 1000-4440( 2019) 03-0523-08
近年来中国南方地区水稻生产水平不断提升,优质粳稻种植面积逐年扩大,多地创造了优质粳稻高产纪录[1-4]。稻麦轮作是中国长江中下游地区重要的耕作制度,近些年,生育期较长的高产优质粳稻在中国南方地区种植面积不断扩大,造成水稻成熟期推迟,加之特殊的气象条件的发生[5],给该地区实现水稻优质高产带来严峻挑战。2016年9-10月长江下游地区水稻在抽穗后较长时间面临了低温寡照多雨的不良天气影响,水稻生产上普遍存在田间发芽和收获后稻谷因高含水量而发芽的现象,严重影响优质粳稻的稻米品质和收获后的安全存储。
穗发芽是指谷物灌浆中后期在遇到连日阴雨且温度适宜条件下而形成的穗部发芽现象,也称胎萌[6-7]。穗发芽和种子休眠密切相关,两者存在显著正相关,而穗发芽和种子休眠是由多基因控制的复杂数量性状[8-9]。赵庆勇等[1O]研究发现,粳稻品种的穗发芽率存在明显的品种(系)间差异,大部分品种(系)的穗发芽率较低,但也存在穗发芽率较高的品种(系)。在穗发芽生理方面,胡伟民等[ll]、张静[12]发现,杂交水稻制种繁种生产中不同杂交种子穗发芽率和穗发芽指数具有显著差异,室内发芽力、α一淀粉酶活性和β一淀粉酶活性与穗发芽的发生率具有相关性。刘百龙等[13]认为,较高的α一淀粉酶活性、种子含水量、可溶性糖和可溶性蛋白质含量是水稻穗发芽的诱因。为了探究穗发芽对稻米品质的影响,马良勇等[14]、黄艳玲等[15]研究发现,发芽严重影响了稻米的加工、外观、营养和蒸煮品质,发芽还导致RVA谱的特征值最高黏度和崩解值减小,降低稻米的食味品质。然而前人关于穗发芽的研究大多是对穗发芽的原因及生理生化变化,同时穗发芽对品质影响的研究多以杂交稻和早籼稻为主,针对穗发芽对粳稻稻米品质相关性状影响的研究尚少。本研究针对水稻生产一线的实际问题,分析穗发芽对长江中下游流域粳稻稻米品质性状影响并探究其内在影响因素,为该地区水稻高产优质栽培提供参考。
1 材料与方法
1.1 供试品种
以中熟中粳品种南粳9108、金粳818、常软06-2为试验材料,参试材料在安徽凤台、江苏扬州和淮安等地穗发芽较严重。其中,优质软米品种[16]南粳9108由江苏省农业科学院提供,金粳818由天津市农业科学院提供,常软06-2由常熟农业科学研究所提供,参试品种均属优质食味粳稻品种。
1.2 试验设计
于2016年10月5日前后,水稻收获前选取以上3个在田间生产中出现较为严重穗发芽的优质稻米品种,筛选未发芽的稻穗,田间集中取样,样品清理去杂后,以不同品种的离体整穗为材料,用蒸馏水清洗后分别置于光照培养箱中,用装设好的喷洒设备对稻穗样品进行间断性地喷水处理,模拟穗发芽发生的环境条件[15,17],直至样品达到破胸露白(处理一)和芽长3-5 cm(处理二)的不同穗发芽程度,将处理后的样品常温晾干备测,室内储存60 d,待其各方面理化性状稳定后再进行稻米品质及蛋白质组分的测定分析,试验重复2次。
1.3 测定项目与方法
1.3.1 加工品质与外观品质测定 水稻脱粒并晒干,用NP-4350型风选机等风量风选,参照《GB/T17891-1999优质稻谷》测定糙米率、精米率、整精米率、垩白粒率、垩白大小、垩白度等。
1.3.2 稻米淀粉黏滞特性 采用澳大利亚NewportScientific仪器公司生产的Super3型黏度测定仪测定,并使用配套软件TCW分析。按照AACC规程(1995-61-02)和RACI标准方法,当米粉的含水量为12.00%时,样品量为3.00g,蒸馏水为25.00g,在搅拌测定过程中,罐内温度50℃下保持1 min,以每1 min 11.84℃的速度上升到95℃并保持2.5 min,再以每1 min 11.84 0C的速度下降到50℃并保持1.4 min。搅拌器在起始10 s内转动速度为960r/min,之后保持在160 r/min。RVA谱特征值用峰值黏度、热浆黏度、最终黏度、崩解值(峰值黏度一热浆黏度)、消减值(最终黏度一峰值黏度)等特征值来表示。
1.3.3 直链淀粉含量及胶稠度测定直链淀粉含量测定:利用碘比色法进行直链淀粉含量测定,称量待测过60目筛的米粉样品0. 100 0 g,置于100ml容量瓶中。加入1.0 ml 95%乙醇,加入9.0 ml1.00 mol/L的氢氧化钠溶液,将容量瓶置沸水中煮10 min后取出,冷却至室温后加蒸馏水定容。吸取5.0 ml样品溶液.加入已盛有半瓶蒸馏水的100ml容量瓶中,再在这容量瓶中加入1.00 ml l.00mol/L的乙酸溶液,使样品酸化,加入1.50 m 1碘液,充分摇匀,用蒸馏水定容,静置20 min。用5.00 m 1的0.09 mol/L的氢氧化钠溶液代替樣品,配制空白溶液。用空白溶液于分光光度计波长620 nm处调节零点并测出有色样品的吸光度值。根据已知直链淀粉含量的标准样品而作出的标准曲线计算样品的直链淀粉含量。胶稠度采用米胶延长法测定。
1.3.4 总蛋白质与蛋白质组分测定用精米样品和分离出的蛋白质组分样品分别制备过0.25 mm孔径筛的样品0.5-0.8g,置于FOSS公司的凯氏定氮仪配套的消化管中,加入质量比为7:1混合的硫酸钾和硫酸铜作催化剂,同时加入浓硫酸,用漏斗封住管口,并置于通风厨中消化1.5 h后,置于全自动凯氏定氮仪中测定总蛋白质含量及蛋白质组分含量。
1.3.5 蛋白质组分的分离 准确称取米粉样品10.0g,加入40 ml正己烷,振荡过夜,去除上清液及残留的液体,干燥24 h,加入40 ml蒸馏水,振荡4h,3 000 9离心10 min,收集上清液,并再次加入40 ml蒸馏水,振荡4h,3 000 g离心10 min,合并2次收集的上清液即为白蛋白提取液。沉淀中加入40 ml 5% NaCl,用与白蛋白提取同样的方式提取球蛋白。再向沉淀中加入0.1 mol/L NaOH,振荡th,提取谷蛋白。沉淀再用70%乙醇提取醇溶蛋白。白蛋白、球蛋白和醇溶蛋白提取液用盐酸分别调pH至4.1、4.3和4.8,3 0009离心15 min,沉淀蛋白质。用蒸馏水清洗沉淀。各蛋白质组分冷冻干燥。1.3.6食味指标采用米饭食味计(STAIA,日本佐竹公司产品)自动测定米饭的气味、光泽、色泽、完整性、味道、口感的评分和综合评分值。
1.4 统计分析
采用Excel进行数据处理分析,DPS软件进行方差分析。
2 结果与分析
2.1 穗发芽的气象因子
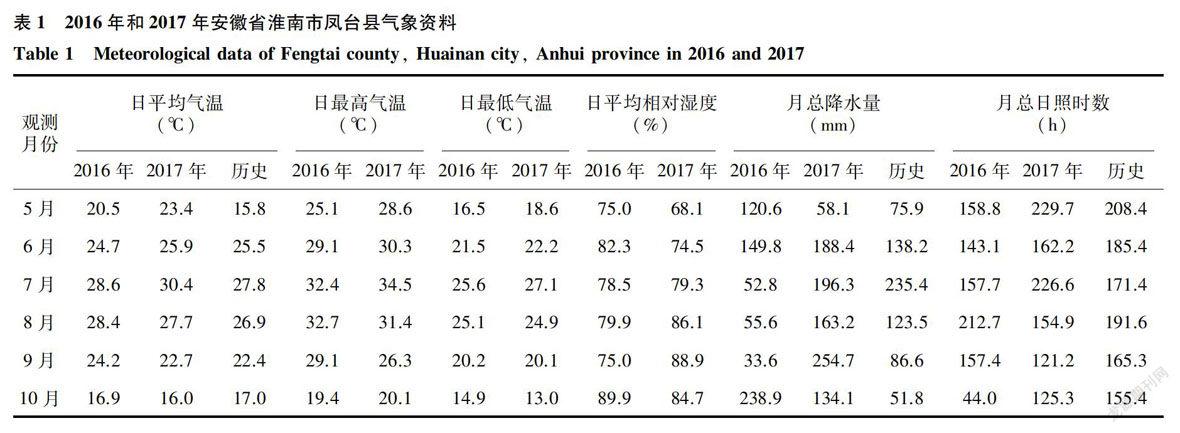
2016年和2017年是苏皖地区穗发芽较为严重的2年,以安徽省淮南市凤台县校外试验点的气象状况为例,由表1可看出,2016年8月和9月的日均温达到28.4℃和24.2℃,高于历史均值(26.9℃和22.4℃),较高的温度加快了籽粒灌浆的速率,促进了早熟。进入10月份以后,连续低温阴雨天气,降水量达到238.9 mm,远高于历史均值的51.8 mm,且水稻生长环境中的平均湿度达到89.9%,因此,不断缩小的温度日较差和连续阴雨寡照造成的田间低温高湿环境,为穗发芽的田间发生创造了较为适宜的气候条件。
2.2 穗发芽对稻米品质影响
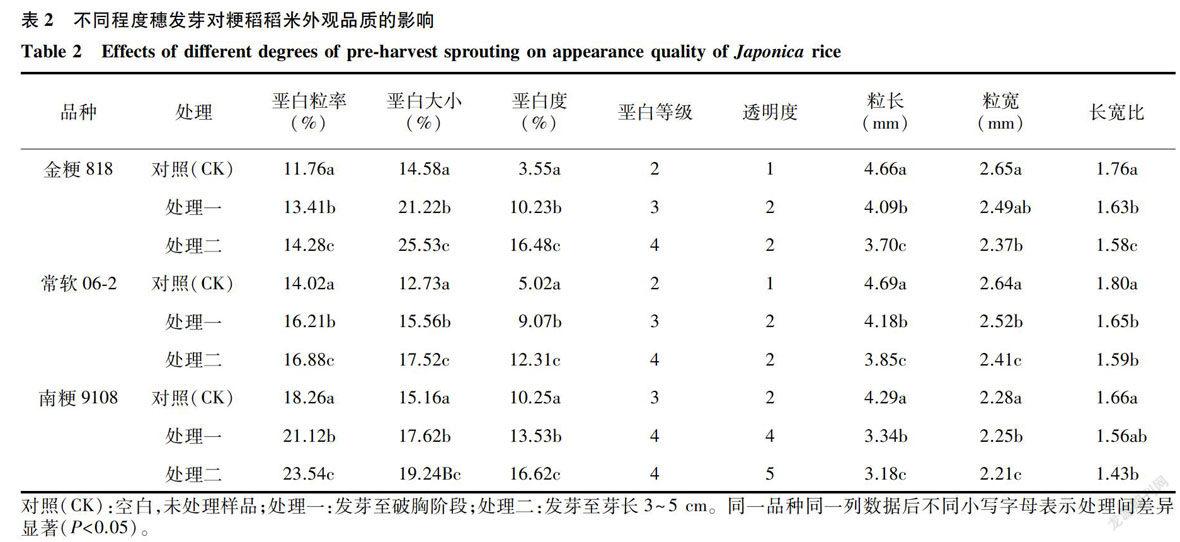
2.2.1 稻米外观品质 外观品质是稻米的重要商品价值指标,外观品质中,又以垩白大小、垩白粒率和粒型为重要评价指标。由表2可知,不同穗发芽程度下,籽粒的外观品质发生了显著变化,随着发芽程度加深,不同粳稻品种的稻米垩白粒率、垩白大小和垩白度均显著增加,垩白等级显著增加,其中处理间垩白粒率的差异性达到6% -23%,以金粳818的垩白度上升幅度最大。同时,不同品种处理间透明度下降明显,这与我们实际观察到的籽粒浑浊现象一致,这与前人的研究结果[18]相似。随着发芽程度的加深,籽粒的粒长、粒宽均显著或极显著地下降,与稻米加工品质变化一致。综上,穗发芽对稻米的外观品质和商品经济价值造成了显著影响,降低了稻米外观品质。
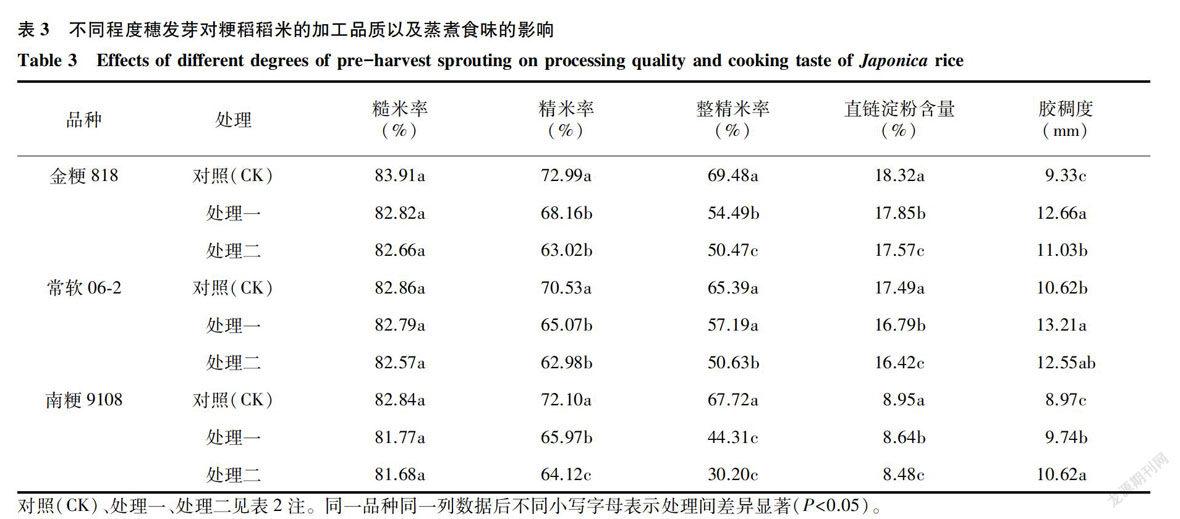
2.2.2 稻米加工品质和蒸煮食味品质 稻米的加工品质根据其品种类型的不同,在不同稻区有不同的表现,南方粳稻品种的稻米品质在江苏中部里下河稻区(扬州)较优[19]。由表3可知,随着穗发芽程度的加深,不同品种稻米糙米率、精米率、整精米率均显著或极显著下降,其中整精米率的下降尤以优质软米品种南粳9108[20]为甚,可见,穗发芽现象对南方稻区的优质软米品种影响严重。
另外,随着发芽程度的加深,稻米籽粒的直链淀粉含量显著或极显著下降,可见,穗发芽对籽粒的食味品质具有显著的负效应,淀粉在穗发芽过程中被机体本身消耗。但是在此并不能简单将直链淀粉含量的下降和蒸煮食味品质的提高相等价,而应该将这种变化归咎于淀粉粒结构的劣性变化,这反而导致了食味品质的下降。品种金粳818和常软06-2的胶稠度随着发芽深度的增加,先增大后减小,在破胸后籽粒胶稠度下降明显,但南粳9108属于典型的半糯性软米,随着穗发芽程度的增加,其黏性显著下降,胶稠度呈上升趋势,这与南粳9108整精米率极显著下降的趋势一致。可见,穗发芽的发生显著降低了稻米的加工和蒸煮食味品质。
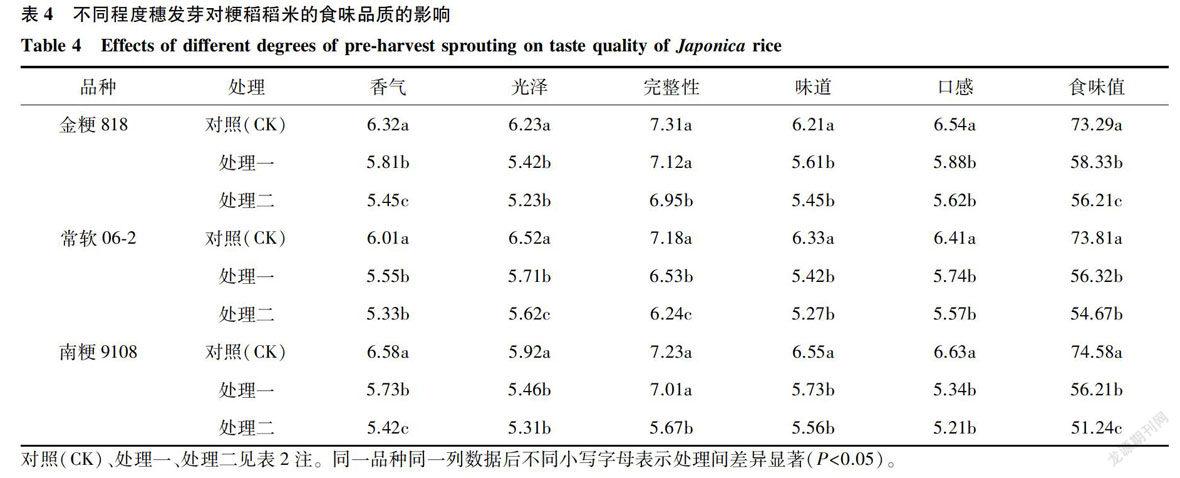
2.2.3 稻米食味品质 由表4可知,不同程度穗发芽对金粳818,常软06-2和南粳9108食味品质的影响达到显著和极显著水平。稻米的食味值、口感、味道、完整性、光泽和香气均随着发芽程度的加深而下降。从变异系数来看,因为食味值反映稻米食味变化的综合情况,其中南粳9108的食味对穗发芽的发生最为敏感,食味值从74.58下降到51.24,口感从6.63下降到5.21。结合表4与表2、表3分析,穗发芽后,各品种间籽粒的光泽、完整性与稻米的外观品质变化同步下降,而口感的变化趋势也和稻米的直链淀粉含量和蛋白质含量的变化趋势一致,这表明食味值与籽粒外观品质与蒸煮食味品质显著相关,而穗发芽对籽粒外观和理化性质的影响降低了米饭的综合食味。
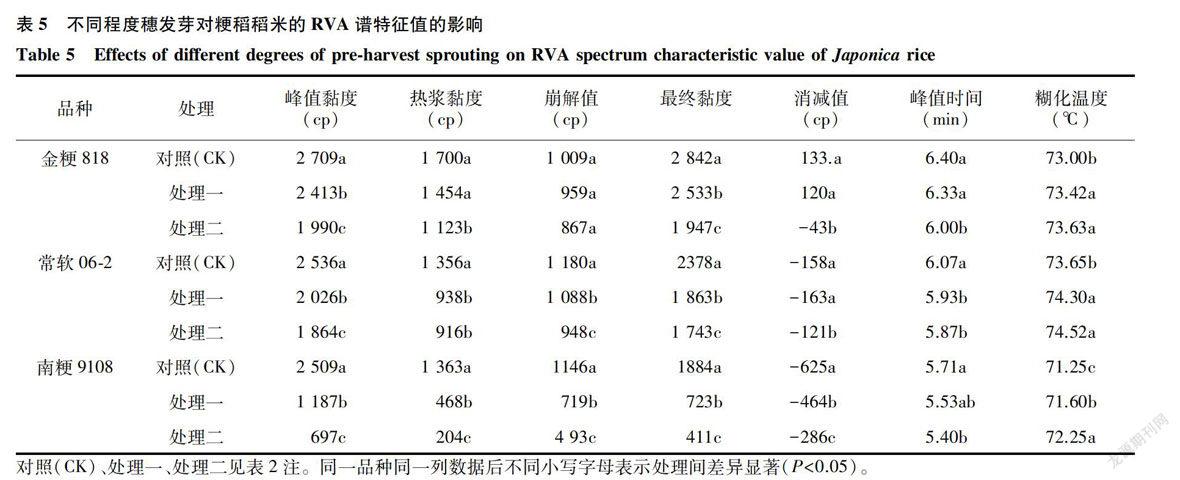
2.2.4 稻米淀粉黏滞特性由表5可知,不同程度穗发芽对3个品种的RVA谱特征值的影响较大。不同处理间的稻米峰值黏度、热浆黏度、崩解值、最终黏度均存在显著或极显著的差异。穗发芽使稻米的峰值黏度、热浆黏度和最终黏度均呈现下降趋势,随着穗发芽程度的加深,金粳818籽粒的崩解值和消减值的变化趋势一致,基本保持不变,而常软06-2和南粳9108两个软米品种的变化趋势与其一致。另外,南粳9108的RVA谱特征值中峰值黏度、热浆黏度和最终黏度显著变化,数据变异性显著高于金粳818和常软06-2。同时,随着穗发芽程度的增加,稻米的峰值时间减少,而糊化温度则呈增加趋势,这说明,穗发芽加快了稻米的糊化过程,但是糊化温度升高。可见,穗发芽问题对稻米黏滞性产生显著或极显著影响,其中软米品种南粳9108对穗发芽问题较为敏感。
2.2.5 稻米总蛋白质及各蛋白质组分含量 由表6可知,不同程度穗发芽对籽粒蛋白质组分有显著影响。但是,清蛋白和球蛋白在籽粒中含量较低,且穗发芽过程中清蛋白含量处理二显著高于处理一和对照,而球蛋白含量随穗发芽程度的加深而显著下降。籽粒蛋白质组分中,谷蛋白和醇溶蛋白含量较高,3个优质稻米品种谷醇比分别达到5. 437、5.496和6.646,但是随着穗发芽程度的加深,籽粒的谷蛋白含量均呈下降趋势,而醇溶蛋白质含量均呈上升趋势,总蛋白质含量呈现显著下降趨势,其中,南粳9108的总蛋白质含量变异系数达到0.043,谷蛋白含量的变异系数达到0.064,可见,穗发芽显著影响籽粒的蛋白质组分,特别是谷蛋白含量,进而影响谷醇比,降低米饭的营养和食味品质。
3 讨论
3.1 不同程度穗发芽对粳稻稻米品质的影响
外观品质是稻米的重要商品价值指标,外观品质中,又以垩白大小、垩白粒率和粒型为重要评价指标。穗发芽条件下,优质稻米的垩白粒率、垩白大小和垩白度均显著增加,垩白等级显著或极显著增加,透明度下降,这与我们实际观察到的穗发芽稻米籽粒浑浊化变化一致。籽粒的长度均值,宽度均值和长宽比均值均呈下降趋势,且稻米的整精米率显著下降,这可能与籽粒的粒型结构和稻米的垩白率存在相关关系[18]。籽粒的加工品质和经济产量的较大下降,可能与穗发芽过程中籽粒淀粉体晶体结构破坏和可溶性糖的转化[21]而导致籽粒表层结构松散有关。
黄燕玲等[15]认为,穗发芽严重影响稻米的加工品质,引起稻米的精米率和整精米率下降,对糙米率的影响不显著,这与我们的研究结果一致,说明稻米穗发芽对籽粒的糊粉层及内部结构存在影响,而不影响稻米的糠层结构。刘百龙等[13]认为,较容易穗发芽的水稻品种的籽粒中可溶性贮藏蛋白质的含量较高,穗发芽后籽粒中蛋白质结构在蛋白酶的作用下快速分解,造成总蛋白质含量减少,这与我们研究的结果一致。钱春荣等[22]认为,直链淀粉含量与蛋白质含量间存在明显的动态关系,稻米食味值随蛋白质含量的下降而增加,当蛋白质含量下降到某一临界值后,随蛋白质含量下降而下降,蛋白质与直链淀粉含量适中的后代株系食味值较高。而边嘉宾等、张欣等23-24认为,直链淀粉含量与食味值呈极显著的负相关关系,蛋白质含量与直链淀粉含量呈不显著的负相关关系,与食味值呈显著负相关关系。这些显著的变化也可以通过本研究中食味值测定结果来验证,本研究中稻米食味值、口感、味道、完整性、光泽和香气均随着发芽程度的加深而下降,其中非软米品种金粳818在不同穗发芽程度处理下食味下降更甚。因此,蛋白质含量过高或过低都会导致稻米蒸煮食味品质变劣,协调蛋白质含量与直链淀粉含量的平衡關系是提高稻米蒸煮食味品质的关键。软米品种的食味优势在于较低的直链淀粉含量和适量的蛋白质含量,而随着穗发芽程度的加深,直链淀粉含量和蛋白质含量均呈下降趋势,可见营养品质显著下降时食味品质存在变化,且这种变化趋势并不能简单的与食味的提高或降低划等号,而是淀粉和蛋白质结构的劣性变化所致[15,18]。
3.2 不同程度穗发芽对粳稻RVA谱特征值的影响
RVA谱特征值,特别是最高黏度、崩解值和消碱值能较好地反应水稻品种间蒸煮食味品质的差异性,已成为评价籽粒淀粉黏滞性和稻米食味的重要指标。陈书强等[25]认为,蒸煮食味品质与淀粉RVA谱特征值和碾米品质相关性最高,淀粉RVA谱特征因子主要通过峰值黏度、冷胶黏度和热浆黏度对食味值和胶稠度的作用来影响蒸煮食味品质优劣。舒庆尧等[26]认为,RVA谱能较好地区分直链淀粉含量相似的稻米的品质优劣。米饭的硬度与消减值呈极显著正相关,与崩解值呈极显著负相关;而米饭的黏性与消减值呈显著负相关。张杰等27]认为,直链淀粉含量与峰值黏度、热浆黏度、崩解值在至少一种环境中呈显著或极显著负相关,而与冷胶黏度、消减值、回复值、峰值时间在至少一种环境中呈显著或极显著正相关。隋炯明等[28-30]研究结果表明,食味与稻米RVA谱特征值中的最高黏度、最低黏度、最终黏度和崩解值呈正相关,与消减值呈负相关,而食味好的品种的最高黏度高、崩解值大,消减值小,且蛋白质含量与热浆黏度、冷胶黏度、崩解值呈显著或极显著负相关。本研究发现,随发芽程度加深,稻米的RVA谱特征值呈现下降趋势,即最高黏度、热浆黏度、最终黏度均显著或极显著降低,进而降低了稻米品质。
3.3 不同程度穗发芽对粳稻蛋白质组分的影响
一般而言,籼稻的蛋白质含量显著低于粳稻,而软米品种作为粳稻中的特殊类型,因其较低的蛋白质和直链淀粉含量以及较好的口感正成为南方水稻育种的新方向[16,0]。粳稻稻米中的蛋白质含量及其组成受品种、产地、生长发育环境特别是灌浆期的气候环境以及加工精度等不同的影响[31]。前人研究结果表明,决定优质食味水稻的蛋白质组分主要是谷蛋白和醇溶蛋白,因此引入谷醇比的概念,提倡选育高谷蛋白低醇溶蛋白的优质水稻品种32-33]。
前人研究发现,发芽萌动过程中蛋白质含量及组分存在显著变化,董召荣等[17]和张艳等[34-35]研究结果表明,随穗发芽程度加深,蛋白质含量逐渐降低,主要蛋白质组分呈下降趋势,而可溶性蛋白质组分含量呈上升趋势,氨基酸含量显著上升,这与本研究结果一致,这可能与穗发芽过程中蛋白酶活性增强,储藏蛋白质被机体内部消耗用以营养生长有关。本研究发现水稻穗发芽过程中,总蛋白质含量呈下降趋势,醇溶蛋白含量呈上升趋势而谷蛋白含量呈下降趋势,而球蛋白与清蛋白含量变化不显著,这说明穗发芽过程中优质食味稻米原有的营养结构被打破,降低稻米食味且又几乎不能被肠胃所吸收的醇溶蛋白含量增加,虽然稻米中增加的游离氨基酸可以提高食味,但稻米食味品质仍显著下降。这说明穗发芽对优质稻米品种营养品质的影响是显著的。
穗发芽现象对南方稻区优质水稻稻米品质和籽粒中蛋白质组分均存在显著影响,随着穗发芽程度的加深,优质水稻稻米品质呈显著下降趋势,其加工品质、外观品质、蒸煮食味品质及营养品质均显著劣化。因此,及时收获以回避高湿天气,选育抗穗发芽优质水稻品种将促进南方地区优质水稻产业发展,有效降低南方稻区水稻生产损失。
参考文献:
[1]郭保卫,朱大伟,朱聪聪,等.有序摆抛栽对粳型超级稻稻米品质的影响[J].作物学报,2015,41(3):487-498.
[2]胡雅杰,钱海军,曹伟伟,等.机插方式和密度对不同穗型水稻品种产量及其构成的影响[J].中国水稻科学,2016,30(5):493-506.
[3]胡雅杰,刑志鹏,龚金龙,等.钵苗机插水稻群体动态特征及高产形成机制的探讨[J].中国农业科学,2014,47(5):865-879.
[4]张洪程,朱聪聪,霍中洋,等,钵苗机插水稻产量形成优势及主要生理生态特点[J].农业T程学报,2013,29( 21):50-59.
[5]李恩菊,赵景波,厄尔尼诺/拉尼娜事件对山东省气候的影响[J].陕西师范大学学报(自然科学版),2010,38(3):80-84.
[6] BEWLEY J D, BRADFORD K J. Seeds: physiology ofdevelopment, germination and dormancy[M]. New York:Springer,2013.
[7]JUNC-SUN BAEK, NAM-JIN CHUNC. Influence of rainfall duringthe ripening stage on pre-harvest sprouting, seed quality, and lon-gevity of rice(Oryza SativaL)[J].Korean J Crop Sci,2014,59( 4):406-412.
[8]MARZOUGUI S, SUGIMOTO K, YAMANOUCHI U.Mapping andcharacterization of seed dormancy QTLs using chromosome segmentsubstitution lines in rice.[J].Theor Appl Genet, 2012. 124(5):893-902.
[9]高永峰,刘继恺,范晶,等.水稻穗发芽调控基因Os- VP1的RNA干涉载体构建及遗传转化研究[J].中国农业科学,2010,43(7):1321-1327.
[10]赵庆勇,陈涛,赵春芳,等.粳稻穗发芽的品种(系)间差异及其影响因素分析[J].江苏农业学报,2017,33(3):481-486.
[11]胡伟民,马华升,樊龙江,等.杂交水稻制种不育系穗上发芽特性[J].作物學报,2003,29(3):441-446.
[12]张静.水稻种子穗发芽及相关可溶性糖的QTL定位[D].北京:中国农业科学院,2013.
[13]刘百龙,王威豪,韦善富,等.水稻种子穗上发芽的生理学初步研究[J].种子,2013,32(1):89-91.
[14]马良勇,杨长登,李西明,等.早稻穗发芽对水稻产量和米质的影响[J].中国稻米,2004,10(1):15-16.
[15]黄艳玲,申广勒,石扬娟,等.穗发芽深度对早稻米品质和RVA谱特征值的影响研究[J].中国农学通报,2008,24(7):119-122.
[16]朱大伟,张洪程,郭保卫,等,中国软米的发展及展望[J].扬州大学学报(农业与生命科学版),2015,36(1):47-52.
[17]董召荣,柯建国,马传喜,等.小麦籽粒发芽深度对其加T品质的影响[J].南京农业大学学报,2000,23(2):9-12.
[18]朱雪梅,邵继荣,杨文钰,等.温度对杂交中籼稻灌浆期淀粉体发育及胚乳透明度的影响[J].作物学报,2005,3l(6): 790-795.
[19]吉志军,尤娟,王龙俊,等.不同基因型水稻稻米加T品质和外观品质的生态型差异[J].南京农业大学学报,2005,28(4):16-20.
[20]赵庆勇,朱镇,张亚东,等.超级稻新品种南粳9108产量稳定性及高产特征分析[J].中国稻米,2016,22(6):61-65.
[21]夏仲炎,王学栋.籼稻品种商品品质性状研究[J].安徽农业科学,1986(3):31-36.
[22]钱春荣,冯延江,杨静,等,水稻籽粒蛋白质含量选择对杂种早代蒸煮食味品质的影响[J].中国水稻科学,2007,21(3):323-326.
[23]边嘉宾,施利利,张欣,等.稻米主要品质性状的相关及主成分分析[J].中国农学通报,2012,28( 24):8-12.
[24]张欣,施利利,丁得亮,等.米饭理化指标与食味品质的相关性研究[J].中国农学通报,2010,26( 12):45-47.
[25]陈书强.粳稻米蒸煮食味品质与其他品质性状的典型相关分析[J].西北农业学报,2015,24(1):60-67.
[26]舒庆尧,吴殿星,夏英武,等.稻米淀粉RVA谱特征与食用品质的关系[J].中国农业科学,1998,31(3):1-4.
[27]张杰,郑蕾娜,蔡跃,等.稻米淀粉RVA谱特征值与直链淀粉、蛋白质含量的相关性及QTL定位分析[J].中国水稻科学,2017,31(1):31-39.
[28]隋炯明,李欣,严松,等.稻米淀粉RVA谱特征与品质性状相关性研究[J].中国农业科学,2005.38(4):657-663.
[29]李刚,邓其明,李双成,等.稻米淀粉RVA谱特征与品质性状的相关性[J].中国水稻科学,2009,23(1):99-102.
[30]刘建伟,三轮精博,后藤清和,等.杂交稻米的理化特性与食味评价的研究[J].中国粮油学报,2003,18(1):5-9.
[31]HAMAKER B R,CRIFFIN V K.Changing the viscoelastic proper-ties of cooked rice throu曲protein disruption[J]. CerealChemistry, 1990, 67(3):261-264.
[32]吴洪恺,刘世家,江玲,等.稻米蛋白质组分及总蛋白质含量与淀粉RVA谱特征值的关系[J].中国水稻科学,2009,23( 4):421-426.
[33]张欣,施利利.稻米蛋白质相关性状与RVA特征谱及食味品质的关系[J].食品科技,2014,39(10):188-191.
[34]张艳,王德森.小麦籽粒发芽对品质的影响[J].麦类作物学报,2006,26(2):58-62.
[35]李毅念,卢大新,萌动小麦的干物质损耗与品质分析[J].农业工程学报,2005,21(8):190-192.
