
吉林省水稻品种的遗传多样性及株型演化分析
2019-09-10 07:22:44张希瑞高文硕王敬国邹德堂
江苏农业学报 2019年3期
张希瑞 高文硕 王敬国 邹德堂

摘要:本研究以51份吉林省不同年代的水稻品种为材料,连续2年对涉及穗、剑叶、倒二叶和倒三叶的21种株型性状进行了调查,并利用152对多态性的SSR引物,对供试品种的遗传多样性水平和不同年代品种间的遗传关系进行了分析。结果表明,多数株型性状在不同年代的品种间均存在极显著的差异(P<0.01)。随育成年代的推移育成品种的剑叶宽度、倒二叶宽度和倒三叶宽度逐渐增大,株高、穗抽出度、穗茎基角、剑叶基角和张角、倒二叶基角和张角以及倒三叶基角和张角逐渐缩小;弯曲穗型是吉林水稻品种的主要穗型。152对SSR引物共检测到648个等位变异,观测等位基因数、基因多样性指数、多态性信息量和Shannon信息指数的均值分别为4.263 0、0.365 4、0.556 3和1.042 2。地方品种与不同年代育成品种之间遗传差异最大。基于遗传相似系数可以将供试品种划分为2个亚群,聚类结果与它们的育成年代基本相同。
关键词:水稻;遗传多样性;株型
中图分类号:S511
文献标识码:A
文章编号: 1000-4440( 2019) 03-0497-09
水稻作为世界五大粮食作物之一,是全世界一半以上人口赖以生存的基本粮食作物。中国栽培水稻主要分为两大类:籼稻和粳稻[1],南方稻区主要以籼稻品种为主,北方稻区主要以粳稻品种为主。吉林省是北方粳稻种植的重要省份,为中国的粮食安全做出了重大贡献。吉林省水稻资源数量较多,来源广泛,但对其遗传多样性及株型性状演化方面的研究却较少,这不利于种质资源的合理利用。研究吉林省水稻品种的遗传多样性,掌握其分布特点及规律,对于充分发掘、利用现有的水稻种质资源,合理选配亲本,拓宽育成品种遗传基础,培育理想株型新品种等具有十分重要的意义。
水稻株型性状与产量关系密切。Fujino等[2]利用26种水稻农业经济性状对北海道100年育种计划水稻株型的演化进行了分析,提出了基于表型变异的适合当地水稻育种计划的遗传多样性模型。徐海等[3]以东北代表性水稻品种辽粳5号与日本的秋田小町为父母本构建RIL群体,研究了其株型性状与产量的关系,得出了株高、倒三节间长度、倒四节间长度和剑叶基角与产量显著正相关的结论。易小林等[4]以桂南杂交水稻为材料,分析株型性状与产量的关系,得到的结果为剑叶长和剑叶宽与产量呈负相关。Lei等[5]通过穗部相关性状对锦糯稻的遗传多样性进行了研究,得出穗部质量性状变异显著高于数量性状变异的结论。
利用分子标记研究水稻遗传多样性的报道也很多,如杨静等[6]利用52对SSR引物对54份黑龙江省水稻品种的遗传多样性进行了分析,结果表明83. 4%的供试品种遗传相似系数在0.74 -0.84。郝伟等[7]应用SSR标记分析了东北三省35个水稻品系的遗传多样性和亲缘关系,得出的结论是吉林和黑龙江水稻品种的SSR遗传多样性水平基本相同,但都低于辽宁水稻品种。Inta等[8]利用SSR和InDel标记发现水稻品种在原生境和栽培地的遗传多样性存在差异,并认为杂交和基因渗入导致了传统地方品种的遗传侵蚀。Sahu等[9]利用42种具有多态性的InDel标记,分析了188份印尼水稻地方品种的遗传分化和遗传多样性,发现籼稻和粳稻之间存在明显的遗传分化。王敬国等[1O]利用154个SSR标记对来自东北亚7个不同地区的9个品种的278份水稻的遗传多样性进行了系统的分析,结果表明吉林省地方品种的遗传多样性高于育成品种。
以往的多数研究都是利用性状[11-13]或分子标记[14-16]单独对作物进行遗传多样性分析,很少将性状和分子标记结合起来,共同研究某一地区水稻品种的遗传多样性及性状演化。而且,至今有关吉林省不同年代栽培水稻资源遗传多样性的研究报道还很少。本研究利用152对多态性SSR标记,对来自中国吉林省的51份水稻品种进行了遗传多样性分析,并比较了不同历史阶段品种的部分株型性状特征,旨在揭示吉林省不同年代品种间的遗传关系及株型演化规律,以期为亲本选配和种质资源的科学利用提供依据。
1 材料与方法
1.1 性状调查
51份吉林水稻品种由东北农业大学水稻研究所提供,连续2年(2016和2017年)种植于东北农业大学阿城实验实习基地,田间管理同生产田。根据不同的育成年份将参试种质分为4组:组群I包括25份地方品种(红毛、红苗稻子、元子二号、黄尖头光陆稻、当地北海道、二节稻、太南稻、大红毛改良北海道、向阳红芒、芦苇稻、红毛稻、金早、红毛稻子、金钩、白大肚兴亚、老光头、小白毛2、红尖、元子黏稻、小白皮田泰、陆羽132-1、小白毛、长春无芒、吉林日落和小白粳子桦甸白);组群Ⅱ为6份1958年到1970年育成的品种(松辽1号、松辽2号、长白5号、吉粳44、吉粳46和长白6号);组群Ⅲ是1971年至1999年育成的10份品種(双丰八号、九稻六号、吉粳61号、九稻八号、长白7号、吉粳63号、吉玉粳、通育211、长白9号和超产1号);组群Ⅳ包括10份2000年之后育成品种(通黏2号、九稻33、九稻46、吉粳88、吉粳502、通育403、长白16号、长白17号、吉粳106和吉特639)。
将所有参试品种已经萌发的种子播种在苗床上。选用秧龄为35 d左右的健壮幼苗插秧,行距30 cm,穴距10 cm,每穴1株,每个品种种植60棵,6行,每行10棵,行长1m。全部品种随机分布在一个池子里。为了避免边际效应,排除每个品种的2个边行以及中间4行每行首尾2棵在内的36棵单株。在每个品种的中央区域随机选择5棵单株挂牌并进行套袋自交。这5棵植株用于性状调查,其中1株的种子和叶片用于下一年的种植和DNA的提取。
齐穗期后,对田间每个挂牌单株的穗抽出度、穗长、剑叶长度、剑叶宽度、倒二叶长度、倒二叶宽度、倒三叶长度和倒三叶宽度进行测定。另外,采用徐正进等的方法对穗颈基角、穗颈张角、剑叶基角、剑叶张角、倒二叶基角、倒二叶张角、倒三叶基角、倒三叶张角进行调查[17]。
穗抽出度是指穗颈节与剑叶叶枕之间的距离。
当穗颈节从剑叶叶鞘中完全抽出时,穗抽出度为正值。当穗颈节不能完全抽出时(包在剑叶叶鞘内),穗抽出度为负值。基角指叶片(穗颈节)基部挺直部分与茎秆的夹角。张角指叶枕(穗颈节)至叶尖(穗尖)的连线与茎秆的夹角。穗长不包括芒的长度。待参试品种完熟后,收取挂牌单株进行考种。分别测定各单株的株高和穗下第一至第四节间长度。
株高是指植株主茎的实际长度(不含芒)。利用主茎分别对每个单株4个节间的长度值进行测量。穗下第一节间长度是指穗颈节至穗下第一节间基部的实际长度,穗下第二节间长度是指穗下第一节间基部至穗下第二节间基部的实际长度,以此类推。各长度单位均为厘米( cm),角度单位均为度(o)。每个样本取5个单株进行测量,利用DPS 9.50进行数据分析[18]。
1.2 总DNA的提取
2017年,以每个品种挂牌单株中1株的幼嫩叶片为材料,采用CTAB方法进行总DNA的提取和纯化[19]。使用琼脂糖凝胶电泳的方法对提取的DNA质量和浓度进行检测。
1.3 引物和多态性检测
参照http//www. gramene.org上的水稻引物信息,下载随机分布于水稻12条染色体上的1 000对SSR引物序列,委托公司进行引物合成。利用小白粳子桦甸白、长白5号、吉粳61和吉粳88 4个品种进行引物筛选。在1000对引物中筛选出152对表现高度多态性且条带清晰的引物,用于遗传多样性分析。
1.4 PCR扩增和产物检测
PCR反应选择10. 00¨l的反应体系。包括1.00μl的模板DNA,1.00μl lOxPCR buffer,0.75μl的MgCl2( 25 mmol/μl),0.10μlTaq酶(10U/μll) ,0. 15μl dNTP( 10 mmol/μl),1.00μl SSR引物(2 μmol/yl),6.00μl ddH20。扩增反应程序为94℃预变性5 min;94℃变性30 s,褪火30 s,72℃延伸30 s,35个循环,4 cC保存。然后,PCR产物在6%的聚丙烯酰胺凝胶上分离,并利用快速银染法[20-21]检测。每对SSR引物检测到1个位点,每条多态性条带视为1个等位基因;有带时赋值为“1”,无带时赋值为“0”。
1.5 数据分析
株型数据利用DPS 9.50进行分析[18]。应用POPGene l.32软件[22]计算51个品种间的遗传一致度、遗传距离[23]、遗传分化系数(Fst)、基因流(Nm)[24]。应用PowerMaker 3.25软[24]计算观测等位基因数目(Na)、遗传多样性指数(He)和多态性信息含量( PIC)[25]。按非加权配对算术平均法( UPGMA),利用NTSYS-pc 2.1软件[26]提供的SHAN模块进行聚类分析,并绘制树状图。利用Ge-nAIEx6.2软件[27]进行分子方差分析。
2 结果与分析
2.1 性状变异
21种株型性状的2年数据平均值统计如表1所示。2年间变异系数最大的5个性状分别是穗茎基角、倒二叶基角、剑叶基角、穗下第四节间长度和倒二叶张角(0.575 7、0.521 1、0.454 2、0.453 6和0.449 9),变异系数最小的5个性状分别为倒二叶宽、剑叶宽、穗下第二节间长度、株高和穗下第一节间长度( 0.151 9、0.147 6、0.143 0、0.128 7和0.123 3).
2.2 株型性状的差异分析
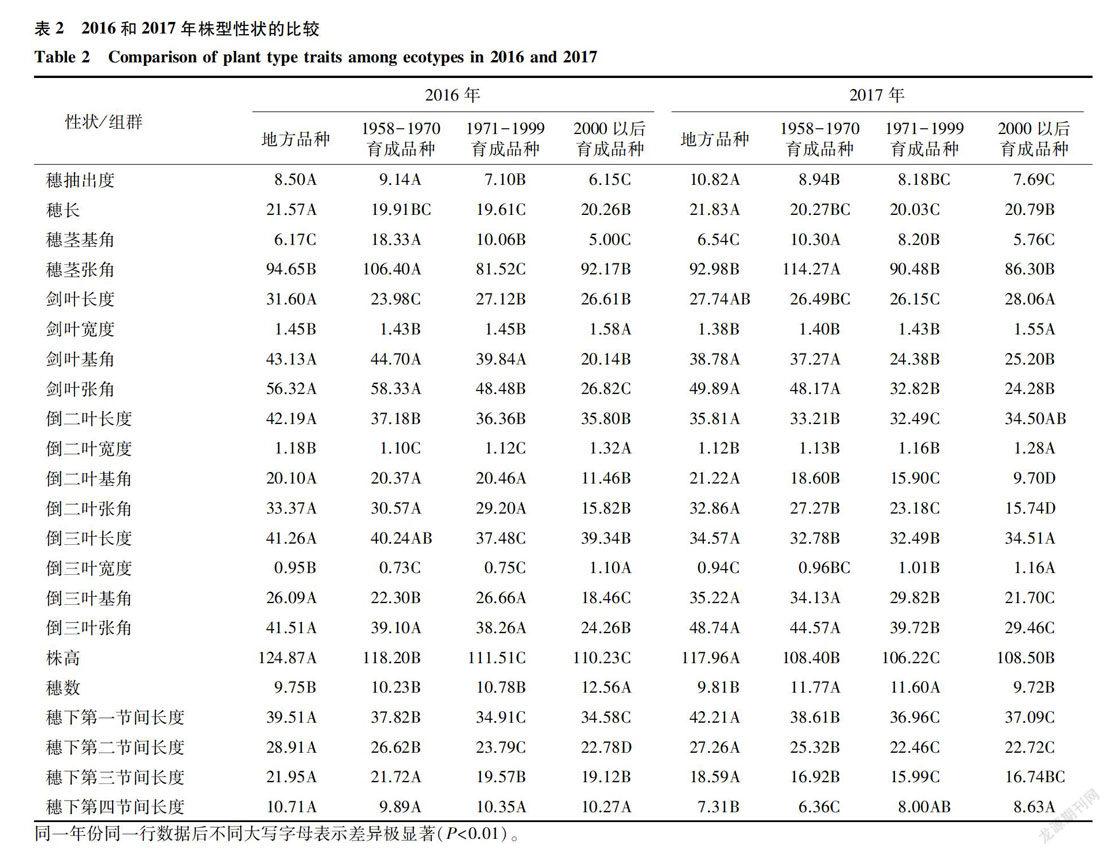
表2为2016年和2017年吉林省水稻种质株型性状组间的多重比较。不同年代品种的穗部呈现如
当穗颈节从剑叶叶鞘中完全抽出时,穗抽出度为正值。当穗颈节不能完全抽出时(包在剑叶叶鞘内),穗抽出度为负值。基角指叶片(穗颈节)基部挺直部分与茎秆的夹角。张角指叶枕(穗颈节)至叶尖(穗尖)的连线与茎秆的夹角。穗长不包括芒的长度。待参试品种完熟后,收取挂牌单株进行考种。分别测定各单株的株高和穗下第一至第四节间长度。
株高是指植株主茎的实际长度(不含芒)。利用主茎分别对每个单株4个节间的长度值进行测量。穗下第一节间长度是指穗颈节至穗下第一节间基部的实际长度,穗下第二节间长度是指穗下第一节间基部至穗下第二节间基部的实际长度,以此类推。各长度单位均为厘米( cm),角度单位均为度(o)。每个样本取5个单株进行测量,利用DPS 9.50进行数据分析[18]。
1.2 总DNA的提取
2017年,以每个品种挂牌单株中1株的幼嫩叶片为材料,采用CTAB方法进行总DNA的提取和纯化[19]。使用琼脂糖凝胶电泳的方法对提取的DNA质量和浓度进行检测。
1.3 引物和多态性检测
参照http//www. gramene.org上的水稻引物信息,下载随机分布于水稻12条染色体上的1 000对SSR引物序列,委托公司进行引物合成。利用小白粳子桦甸白、长白5号、吉粳61和吉粳88 4个品种进行引物筛选。在1000对引物中筛选出152对表现高度多态性且条带清晰的引物,用于遗传多样性分析。
1.4 PCR扩增和产物检测
PCR反应选择10. 00¨l的反应体系。包括1.00μl的模板DNA,1.00μl lOxPCR buffer,0.75μl的MgCl2( 25 mmol/μl),0.10μlTaq酶(10U/μll) ,0. 15μl dNTP( 10 mmol/μl),1.00μl SSR引物(2 μmol/yl),6.00μl ddH20。擴增反应程序为94℃预变性5 min;94℃变性30 s,褪火30 s,72℃延伸30 s,35个循环,4 cC保存。然后,PCR产物在6%的聚丙烯酰胺凝胶上分离,并利用快速银染法[20-21]检测。每对SSR引物检测到1个位点,每条多态性条带视为1个等位基因;有带时赋值为“1”,无带时赋值为“0”。
1.5 数据分析
株型数据利用DPS 9.50进行分析[18]。应用POPGene l.32软件[22]计算51个品种间的遗传一致度、遗传距离[23]、遗传分化系数(Fst)、基因流(Nm)[24]。应用PowerMaker 3.25软[24]计算观测等位基因数目(Na)、遗传多样性指数(He)和多态性信息含量( PIC)[25]。按非加权配对算术平均法( UPGMA),利用NTSYS-pc 2.1软件[26]提供的SHAN模块进行聚类分析,并绘制树状图。利用Ge-nAIEx6.2软件[27]进行分子方差分析。
2 结果与分析
2.1 性状变异
21种株型性状的2年数据平均值统计如表1所示。2年间变异系数最大的5个性状分别是穗茎基角、倒二叶基角、剑叶基角、穗下第四节间长度和倒二叶张角(0.575 7、0.521 1、0.454 2、0.453 6和0.449 9),变异系数最小的5个性状分别为倒二叶宽、剑叶宽、穗下第二节间长度、株高和穗下第一节间长度( 0.151 9、0.147 6、0.143 0、0.128 7和0.123 3).
2.2 株型性状的差异分析
表2为2016年和2017年吉林省水稻种质株型性状组间的多重比较。不同年代品种的穗部呈现如下的特征:地方品种和1958-1970年间育成品种的穗抽出度都大于另外2个年代的品种;地方品种的穗抽出度极显著大于1971-1999年间和2000年以后育成品种,2000年以后育成品种的穗抽出度在2年里均最小。地方品种的穗长极显著大于其他年代育成品种,2000年以后育成品种的穗长较长。1971-1999年间育成品种的穗长最短,而且与其他年代的品种之间都呈现极显著的差异。1958-1970年间育成品种的穗茎基角和穗茎张角都极显著大于其他年代的育成品种;1971-1999年间和2000以后育成品种的穗茎基角极显著的小于1958-1970年间育成品种,而且二者之间存在极显著的差异。这些差异表明,第一,育成品种的穗长逐渐增大,而穗抽出度逐渐缩小;第二,弯曲穗型是吉林省水稻品种的主要穗型。
不同年代品种的剑叶呈现如下的差异:2年里,2000年以后育成品种的剑叶宽度都大于其他年代育成品种,而且差异均达到了极显著水平;按照剑叶基角和张角排序,2年里均呈现1958-1970年育成品种>1971-1999年育成品种>2000以后育成品种的趋势,2016年三者之间的剑叶张角均达到极显著差异,而1971-1999年育成品种和2000以后育成品种的剑叶基角和剑叶张角在2017年的差异却没有达到极显著水平。
不同年代品种的倒二叶性状呈现如下的特征:地方品种的倒二叶长度在2年里均是最大的,而且在2年里与其他年代品种间均达到了极显著差异;2000年以后育成品种的倒二叶宽度在2年里均最宽,而且极显著大于其他年代品种;按照倒二叶基角和张角排序,2年里2000年以后育成品种均极显著大于其他3组,但是地方品种、1958-1970年育成品种和1971-1999年育成品种的倒二叶基角、张角在2017年的差异不显著,而在2016年差异均极显著。
不同年代品种的倒三叶性状呈现如下的差异:2年里,地方品种的倒三叶长度都大于其他年代品种,且与1971-1999年育成品种差异极显著;2000年以后育成品种的倒三叶宽度极显著大于其他3组。以上关于剑叶、倒二叶、倒三叶性状的差异分析结果表明:育成品种的剑叶宽度和倒二叶宽度逐渐增加,而剑叶基角和剑叶张角逐渐缩小。
2年中,地方品种的株高均极显著高于其他年代育成品种;1971-1999年育成品种的株高均极显著低于其他组;这说明地方品种株高最高。分解到节间长度上来看,2年里地方品种的穗下第一节间长度、穗下第二节间长度和穗下第三节间长度都极显著大于其他3组,这应该是造成地方品种株高最高的主要原因;1971-1999年間育成品种的穗下第一节间长度和穗下第二节间长度都低于其他年代的品种,这应该在一定程度上决定了这一时期的品种株高最低。各个组群的穗数在2年里变化规律不明显,1971-1999年育成品种的穗数在2年里都是较多的,地方品种的穗数在2年里都几乎是最少的。这说明不同时期品种的分蘖能力并没有显著的差别。
上述株型性状的差异分析可以归纳出吉林省不同年代品种的株型特点为:地方品种株高最高,穗下第一、第二和第三节间长度最长,倒二叶、倒三叶长度最大;1958-1970年育成品种的穗茎基角和张角最大,第四节间长度最短,倒二叶和倒三叶宽度较小;1971-1999年育成品种的株高最矮、穗下第一节间和第二节间长度最小;2000年以后育成品种的穗抽出度、穗茎基角、倒二叶基角和张角都最小,剑叶、倒二叶和倒三叶宽度均最大。
此外,上述株型性状的差异分析还可以归纳出吉林省育成品种株型演化的特征为:育成品种的剑叶宽度、倒二叶宽度和倒三叶宽度逐渐增大,株高逐渐降低并趋于稳定,穗抽出度逐渐缩短,穗茎基角、剑叶的基角和张角、倒二叶的基角和张角以及倒三叶的基角和张角逐渐缩小;弯曲穗型是吉林品种的主要穗型。
2.3 SSR引物多态性分析
152对多态性SSR引物共检测到648个等位变异。观测等位基因数( Na)的范围为从2到9,均值为4. 263。基因多样性指数(He)的范围为0.071 1-0.666 7,均值为0.365 4。多态性信息量(PIC)范围是0.134 1- 0.826 l,均值为0.556 3。Shannon信息指数(I)范围为0.198 5 - 1.966 0,平均值是1.042 2。Na、He和PIC排在前十的位点分别为RM1347、RM1340、RM336、RM1350、RM1379、RM1360、RM1374、RM1337、RM1365和RM1369,分别位于第2、第6、第7、第3、第2、第1、第10、第12、第7和第6染色体上。
2.4 不同组群之间的遗传关系
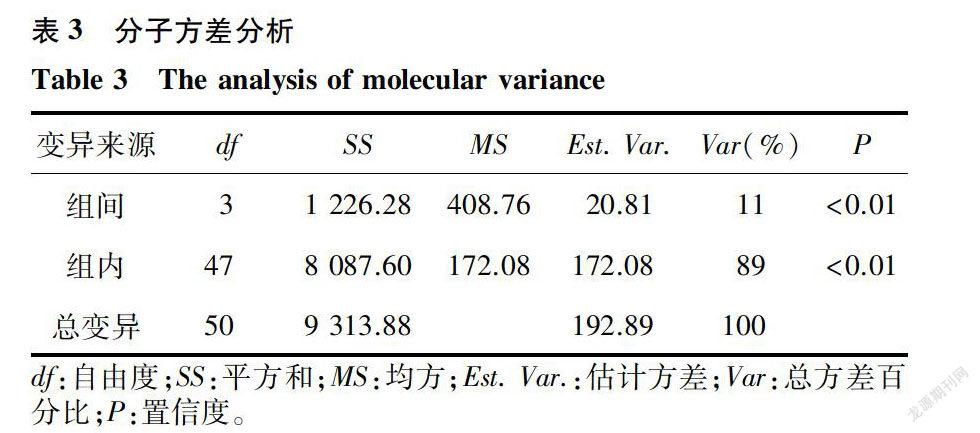
分子方差分析结果表明,组群内和组群间都存在显著的變异(P<0.01)。其中组群间变异为11%,组群内的变异为89%(表3)。
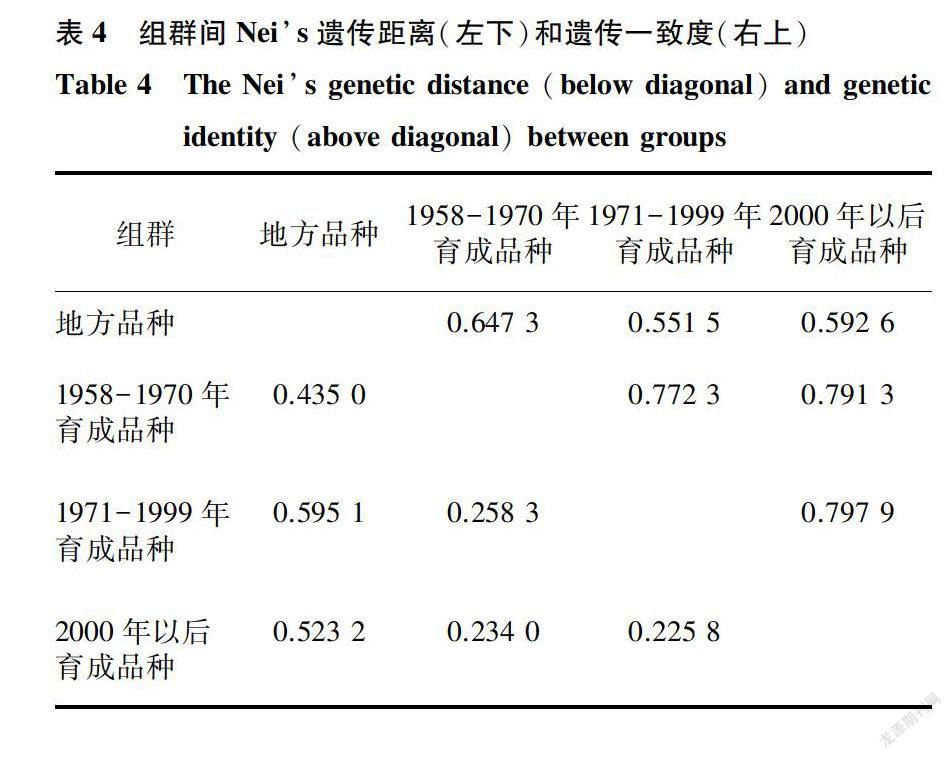
不同组群间的遗传距离和遗传一致度分析结果表明,地方品种与1958-1970年育成品种、2000年以后育成品种和1971-1999年育成品种之间,具有最大的遗传距离(0.435 0、0.523 2和0.595 1)和最小的遗传一致度(0.647 3、0.551 5和0.592 6)。1971-1999年育成品种和2000年以后育成品种之间的遗传距离最小( 0.225 8),遗传一致度最大(0.797 9)。这说明吉林省地方品种与不同年代育成品种之间遗传差异最大,1971-1999年育成品种与2000年以后育成品种间遗传差异最小。此外,1958-1970年育成品种、1971-1999年育成品种、2000年以后育成品种两两之间的遗传距离(0.258 3、0.234 0和0.225 8)都小于它们分别与地方品种之间的遗传距离(0.435 0、0.595 1和0.523 2),同时三者两两之间的遗传一致度(0.772 3、0.791 3和0.792 9)都大于它们分别与地方品种之间的遗传一致度(0.647 3、0.551 5和0.592 6)(表4)。这说明吉林省育成品种间的遗传差异都小于它们分别与地方品种间的遗传差异。
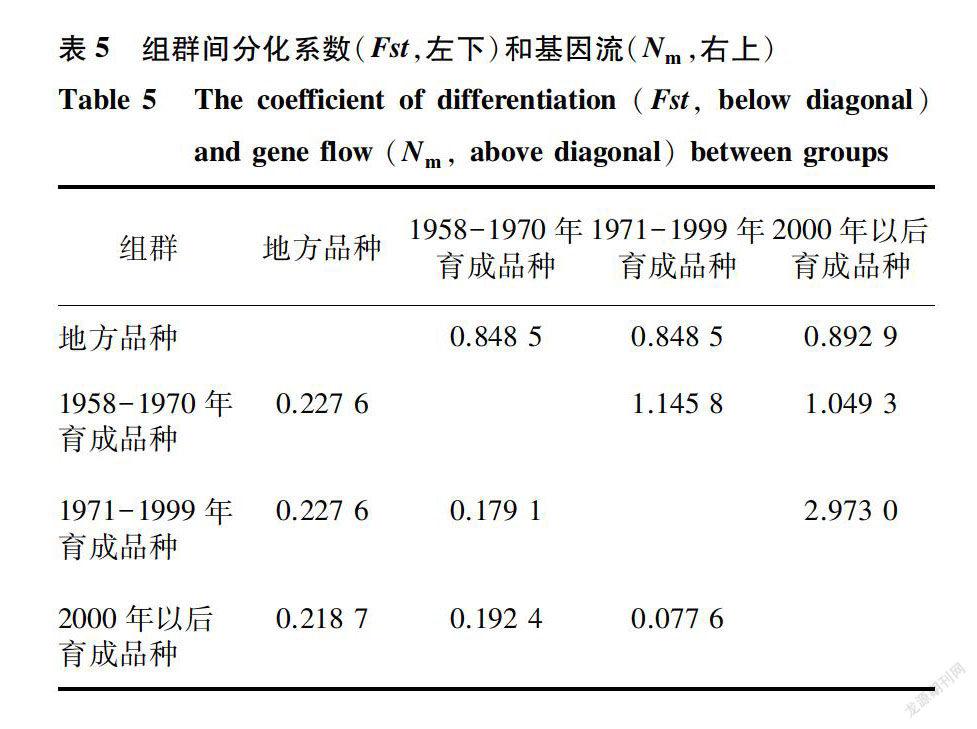
组群间遗传分化系数(Fst)和基因流(Nm)的分析结果(表5)表明,组群间Fst最小为0.077 6,最大为0.227 6,平均值为0.187 2,这表示各组群之间的变异为18. 72%,组群内的变异为81. 28%,这与分子方差分析的结果大致相同。不同组群间Nm的最小值为0.848 5,最大值为2.973 0,平均值为1.293 0。地方品种与1958-1970年育成品种、1971-1999年育成品种和2000年以后育成品种具有最大的遗传分化系数(0.227 6、0.227 6和0.218 7)和最小的基因流(0.848 5、0.848 5和0.892 9)。1971-1999年育成品种和2000年以后育成品种之间的遗传分化系数最小(0.077 6)、基因流最大(2.973 0)。1958-1970年育成品种、1971- 1999年育成品种、2000年以后育成品种两两之间的遗传分化系数( 0.179 l、0.192 4和0.077 6)都小于它们分别与地方品种之间的遗传分化系数(0.227 6、0.227 6和0.218 7),同时三者两两之间的基因流(1.145 8、1.049 3和2.973 0)都大于它们分别与地方品种之间的基因流(0.848 5、0.848 5和0.892 9)。这与遗传距离和遗传一致度的分析结果相同。
2.5 分子聚类分析
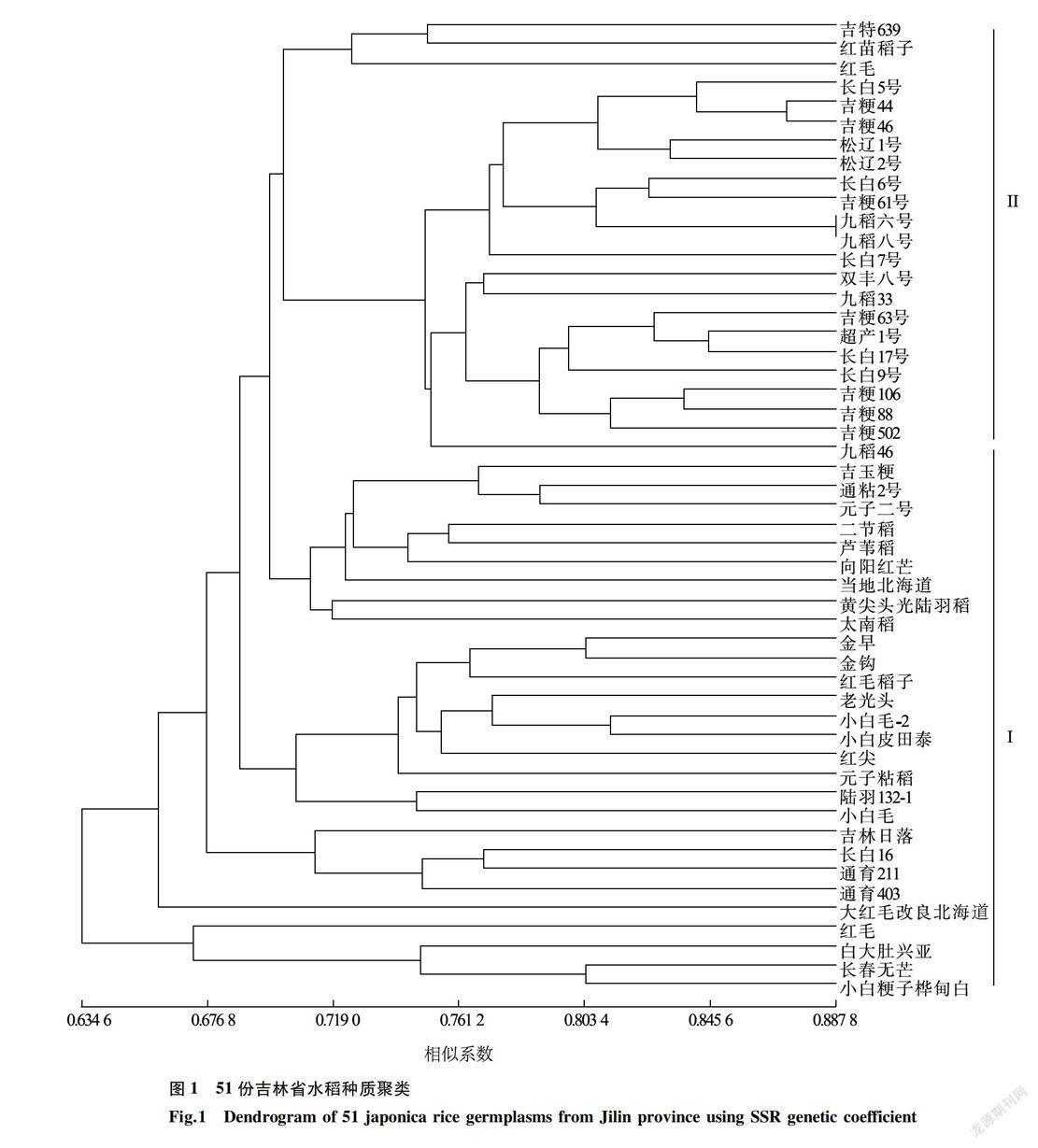
当遗传相似系数取值为0.697 9时,供试的51份水稻种质可以被划分为2个亚群(图1)。第一亚群以地方品种为主,包括23个地方品种和5个育成品种。这5个育成品种分别是1971年至1999年育成的吉玉粳和通育211,以及2000年以后育成的通粘2号、长白16和通育403,都直接或间接的含有日本种质血缘。第二亚群以育成品种为主,包括21个育成品种和2个地方品种。
在遗传相似系数取值为0.751 3时,可将第二亚群再分为3个小亚群。第一小亚群包括2个地方品种和吉特639(2000年以后育成)。第二小亚群包括10个品种,以1958年至1970年育成品种为主,包括这一年代育成的所有6个品种以及4个1971年至1999年育成品种。需要指出的是,在第二小亚群中的吉粳61、九稻六号、九稻八号和长白7号这4个品种是1971年至1999年育成品种中育成时期最早的,都不晚于1986年。第三小亚群也有10个品种,包括6个2000年以后育成的品种以及4个1971年至1999年的育成品种。以上分析结果表明,供试品种的聚类结果与它们的育成年代基本相同。
3 讨论
3.1 水稻株型
水稻株型是指与水稻品种产量能力相关的特征或水稻植株在空间上的排列方式,即长势,具体性状包括株高、节间长度、穗型、叶长、叶宽、叶基角、叶张角、分蘖数和生物量等。有关水稻株型性状的研究报道有很多,如韦还和等[28]以超级稻甬优12为材料研究高产、更高产、超高产3个群体的株型特征,得出超高产群体在穗长、穗粒数、一次和二次枝梗数4个性状上均高于其他2个群体的结论。孙成明等通过对水稻株型概念及其研究进展的分析,阐明了株型对水稻栽培的重要性和必要性[29]。株型性状易受环境条件的影响,采用多年的重复试验可以减少环境误差。本研究对吉林省水稻品种2年的株型性状调查结果进行多重比较,分析21种株型性状在2年中的共同的趋势。结果表明不同组群间一些株型性状存在显著差异。通过株型性状的差异分析,不仅总结了吉林省不同年代水稻品种的株型特征,而且对吉林省育成品种的株型演化规律进行了总结。
3.2 SSR标记分析
利用SSR标记研究不同水稻遗传多样性的报道已有很多,如刘丹[30]利用54个多态性SSR引物对东北近40年育成的粳稻品种的遗传多样进行了分析,得出了与本研究结果相一致的结论。李红宇等[31]利用53个SSR标记结合24种表型性状对东北三省107份推广水稻品种的遗传多样性进行了分析,结果为遗传多样性呈现黑龙江>吉林>辽宁的趋势。孙健等[32]利用42对SSR标记对黑龙江主栽水稻品种的遗传多样性进行了分析,共检测到139个等位基因,分析结果表明黑龙江水稻遗传基础狭窄。陈英华等[33]利用SSR标记分析了东北三省有代表性的区域试验水稻品种的遗传多样性,指出东北稻区整体的遗传多样性狭窄,明显低于其他稻区的水平。Chen等[34]利用30对多态性SSR标记对53份亚洲稻和非洲稻种质的遗传多样性进行了分析,系统发生树揭示53种基因型分属于3个亚群,并筛选出10个可区分种质差异的特异性标记。Muhamad等利用SSR和SNP 2种分子标记对印度尼西亚过去60年里水稻育成品种间的亲缘关系及株型演化进行了分析,结果表明1943 -1966年育成品种的遗传多样性明显大于其他年代育成品种,并提出表型和分子的变异可能是由于当地育种工作的杂交和选择造成的论断[35]。本研究对来自吉林省的地方品种和不同育种年代的水稻种质的SSR遗传多样性进行分析,得出总体表现遗传多样性水平低、遗传基础狭窄的结果。
3.3 吉林水稻种质间的亲缘关系
本研究利用随机分布在水稻12条染色体的152對SSR引物对51份吉林省水稻种质进行分析,其遗传相似系数高达0. 63以上,且多数育成品种聚为一类,在分子水平上进一步证明了吉林水稻种质的亲缘关系较近,遗传基础狭窄,这与郝伟等的研究结果一致[7]。追溯吉林省水稻系谱,绝大多数育成品种含有骨干亲本松辽2号、松辽4号和元子2号等的血缘,且吉林地方品种匮乏,直接或间接利用的亲本多引自黑龙江和日本,这可能是造成其遗传基础狭窄的主要因素。为拓宽吉林水稻种质遗传背景,育种家们需要广泛引进中国优异水稻种质资源,通过人工杂交,聚合有利基因,培育优质品种。
4 结论
吉林省不同年代品种的株型特点:地方品种的株高、穗下第一节间长度、穗下第二节间长度和穗下第三节间长度都最长,倒三叶宽度最小,与其他年代品种间差异极显著。1958-1970年育成品种株高最矮,穗下第三节间长度和穗下第四节间长度都最短、倒三叶长度最小,与其他年代品种间差异极显著;倒二叶长度也最小。1971- 1999年育成品种穗数最多。2000以后育成品种倒二叶基角最小,且与其他年代品种间差异极显著,剑叶基角和剑叶张角最小,倒二叶最宽、倒三叶基角和张角最小。
吉林省育成品种株型演化的特征:育成品种的株高、穗长、剑叶宽度和倒二叶宽度逐渐增大,穗抽出度、剑叶的基角和张角以及倒二叶的基角和张角逐渐缩小;弯曲穗型是吉林品种的主要穗型。
152对SSR引物在51份参试材料中共扩增出648个等位变异,观测等位基因数、基因多样性指数、多态性信息量和Shannon信息指数的均值分别为4.263 0、0.365 4、0.556 3和1.042 2。吉林省地方品种与不同年代育成品种之间遗传差异最大,不同年代育成品种间的遗传差异都小于它们分别与地方品种间的遗传差异。
供试品种被聚类为2个亚群,聚类结果与品种的育成年代基本相同。
参考文献:
[1] DALLY A M,SECOND G.Chloroplast DNA diversity in wild andcultivated species of rice( Genus Oryza, section Oryza). Cladistic-mutation and genetic-distance analysis[J].Tagtheoretical&Ap-plied Geneticstheoretische Und Angewandte Genetik. 1990, 80(2):209-222.
[2] FUJINO K,NISHIMURA T,KIUCHI H,et al-Phenotypicchanges during lOO-year rice breeding programs in Hokkaido[J].Breeding Science, 2017 ,67(5):528-534.
[3]徐海,宫彦龙,夏原野,等.中日水稻品种杂交后代的株型性状与产量和品质的关系[J].中国水稻科学,2016,30(3): 283-290.
[4]易小林,陈会鲜,王彩先,等.桂东南地区杂交水稻理想株型初探[J]。南方农业学报,2016,47(2):196-200.
[5] LEI Q,ZHOU J,ZHANG W, et al-Morphological diversity ofpanicle traits in Kam fragrant glutinous rice( OrTza sativa)[J].Genetic Resources&Crop Evolution, 2018 ,65(3):775-786.
[6]杨静,刘海英,金正勋,等,黑龙江省水稻品种SSR标记遗传多样性分析[J].东北农业大学学报,2008,39(6):1-10.
[7]郝伟,张旭,徐正进,等.东北三省水稻遗传多样性和亲缘关系的SSR分析[J].河南农业科学,2008(4),18-24.
[8] INTA A,BALSLEV H. GUSTAFSSON MHG, et al-Cenetic di-versity patterns of rice( OrTza sativa L.) landraces after migrationby Tai Lue and Akha between China and Thailand[ J]. CeneticResources& Crop Evolution, 2016 ,63(5):845-858.
[9] SAHU P K,MONDAL S, SHARMA D,et al.InDel marker basedgenetic differentiation and genetic diversitv in traditional rice( Oryza sativa L.) landraces of Chhattisgarh, India[J].PLoSONE, 2017 ,12( 11): e0188864.
[10]王敬国.东北亚地区粳稻遗传多样性及主要株型性状与SSR标记的关联分析[D].哈尔滨:东北林业大学,2014.
[11]郭元元,周生茂,陈振东,等.广西黄瓜地方品种鉴定评价及遗传多样性分析[J].南方农业学报,2018,49(7):1273-1281.
[12]王维婷,郭溆,刘超,等,黄秋葵种质资源结实特点遗传多样性评价[J].山东农业科学,2017,49(2):26-29,35.
[13]芮文婧,张倩男,王晓敏,等.47份大果番茄种质资源表型性状的遗传多样性[J].江苏农业科学,2017,45 (12):92-95.
[14]曾绍贵,朱邦彤,罗木旺,等.100份朝天椒的农艺性状和SRAP标记遗传多样性分析[J]。江苏农业学报,2018,34(4):871-879.
[15]余青青,王普昶,赵丽丽,等.7个燕麦品种的ISSR遗传多样性分析[J].江苏农业科学,2018 ,46(19):34-37.
[16]王少铭,侯颖辉,罗莉斯,等,基于SRAP和ISSR标记的薄荷种质资源遗传多样性及亲缘关系分析[J].南方农业学报,2018,49(11):2148-2154.
[17]徐正进,董克,水稻叶片基角,开张角和披垂度的同时测定方法[J].沈阳农业大学学报,1991(2):185-187.
[18]唐启义.DPS数据处理系统一实验设计、统计分析及数据挖掘[M].北京:科学出版社,2007.
[19] EDWARDS K, JOHNSTONE C,THOMPSON C.A simple andrapid method for the preparation of plant genomic DNA for PCR a—nalysis[J].Nucleic Acids Research, 1991, 19(6):1349.[20] TRICIANO R, CAETANO-ANOLLES G.Laboratory exercises onDNA amplification fingerprinting for evaluating the molecular di- versity of horticultural species[J].Hort Technology, 1998,8(6):413-423.
[21] CAETANO G,SCHLARBAUM S,Trigiano R.DNA amplificationfingerprinting and marker screening for pseudo-testcross mapping offlowering dogwood( Comus floridaL)[J].Euphytica, 1999. 106(3):209-222.
[22] YEH F,YANG R, BOYLE T.Popgene Version l.32 MicrosoftWindows-based freeware for populations genetic analysis[J].Uni-versitv of Alberta. 1999, 38(8):212-213.
[23] NEI M. Cenetic distance between populations[J].American Natu-ralist ,1972 ,106( 949): 283-292.
[24] LIU K, MUSE S V.PowerMarker: an integrated analysis environ-ment for genetic marker analysis[J].Bioinformatics. 2005. 21(9):2128-2129.
[25] NEI M, TAJIMA F,TATENO Y.Accuracy of estimated phyloge-netic trees from molecular data[J]. Joumal of MolecularEvolution,1982,18(6):387-404.
[26] ROHLF F J.NTSYS-pc-numerical taxonomy and multivariatesvstem analysis [J].The American Statistician, 1987, 41(4):330.
[27] PEAKALL R, SMOUSE P E.GenAIEx 6.5: genetic analysis inExcel. Population genetic software for teaching and research-an up-date[J].Bioinformatics, 2012, 28( 28): 2537-2539.
[28]韋还和,李超,张洪程,等.水稻甬优12不同产量群体的株型特征[J].作物学报,2014 ,40( 12):2160-2168.
[29]孙成明,苏祖芳,水稻株型的研究进展(综述)[J].上海农业学报,2004 ,20(1):41-44.
[30]刘丹.东北粳稻遗传多样性及穗部性状基因定位研究[D]沈阳:沈阳农业大学,2014.
[31]李红宇,侯昱铭,陈英华,等,用SSR标记评估东北三省水稻推广品种的遗传多样性[J].中国水稻科学,2009, 23(4): 383-390.
[32]孙健,王敬国,刘化龙,等,黑龙江省主栽水稻品种的遗传多样性分析[J].作物杂志,2011(1):63-67.
[33]陈英华,李红宇,侯昱铭,等.东北地区水稻种质资源遗传多样性分析[J].华北农学报,2009,24(3):165-173.
[34] CHEN C,HE W, NASSIROU T Y, et al.Molecular characteriza-tion and genetic diversitv of different genotypes of Oryza sativa andOryza glaberrima[J].Electronic Journal of Biotechnology, 2017,30(8):48-57.
[35] MUHAMAD K, EBANA K. FUKUOKA S,et al.Genetic relation-ships among improved varieties of rice( Oryza sativa L.) in Indo-nesia over the last 60 vears as revealed by morphological traits andDNA markers[J].Cenetic Resources&Crop Evolution, 2016, 64(4):1-15.
(责任编辑:陈海霞)
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27 19:44:27
作物学报(2022年7期)2022-05-11 12:58:02
军事文摘(2021年22期)2021-11-26 00:43:51
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中国农业信息(2017年12期)2017-09-03 04:33:01
上海农业学报(2017年4期)2017-04-10 12:40:28
长江蔬菜·学术版(2016年12期)2017-01-12 20:57:59
养生保健指南(2016年12期)2017-01-06 22:20:23
中国科技博览(2016年25期)2016-12-20 20:04:30
