
艾烟对SAMP8小鼠嗅球形态和嗅球内谷氨酸、γ-氨基丁酸和乙酰胆碱酯酶的影响
2019-09-10 07:22左滢竹林瑶赵百孝
世界中医药 2019年5期
左滢竹 林瑶 赵百孝
摘要 目的:觀察艾烟干预对快速老化SAMP8小鼠嗅球形态和嗅球内谷氨酸(Glu)、γ-氨基丁酸(GaBa)和乙酰胆碱酯酶(AChE)表达的影响,探讨艾烟的可能效应途径。方法:选取6月龄雄性快速老化模型SAMP8小鼠48只并随机分为艾烟组、模型组、嗅觉障碍组和嗅觉障碍艾烟组,以12只SAMR1作为空白对照组对照。3-甲基吲哚(3-MI)腹腔注射制作嗅觉障碍模型。艾烟组和嗅觉障碍艾烟组每日予以艾烟吸嗅30 min,其余组正常空气吸嗅。干预6周后通过苏木精-伊红(HE)染色观察各组嗅球形态,并利用酶联免疫吸附试验法比较嗅球内Glu、GaBa及AChE表达情况。结果:各组嗅球结构未见明显差异,艾烟组颗粒细胞密度略高于模型组、嗅觉障碍组和嗅障艾烟组。艾烟组Glu、AChE较模型组显著降低,差异有统计学意义(P<0.05),且AChE含量低于嗅觉障碍组,差异有统计学意义(P<0.05)。嗅障艾烟组Glu含量介于模型组和艾烟组之间,3组差异有统计学意义(P<0.05)。结论:一定浓度的艾烟干预可以良性调节嗅球内神经递质含量,并通过嗅觉通路的神经投射干预脑内神经递质的表达,改善氧化应激。
关键词 艾烟;嗅球;神经递质;嗅觉通路;谷氨酸;γ-氨基丁酸;乙酰胆碱酯酶
Abstract Objective:To observe the effects of moxa smoke on the olfactory bulb morphology and the expression of glutamate acid (Glu),gamma-aminobutyric acid (GaBa) and acetylcholinesterase (AChE) in the olfactory bulb of aging SAMP8 mice,in order to explore the possible pathway of moxa smoke effect.Methods:A total of 48 six-month-old male SAMP8 mice were randomly divided into 3 groups:a moxa smoke group,a model group,a dysosmia group and a dysosmia model treated with moxa smoke group.12 SAMR1 mice were used as control group.3-methylindole (3-MI) was injected intraperitoneally to produce an olfactory dysfunction model.The moxa smoke group and dysosmia model treated with moxa smoke group inhaled moxa smoke for 30 min every day.Other groups inhaled normal air.After 6 weeks′ intervention,the morphology of olfactory bulb was observed by HE staining,and the contents of Glu,GaBa and AChE in olfactory bulb were tested by ELISA.Results:There was no significant difference in olfactory bulb structure among the 5 groups.The density of granular cells in the inner plexus layer of the moxa smoke group was slightly higher than that of the model group,the dysosmia group and the dysosmia treated with moxa smoke group.Contents of Glu and AChE in moxa smoke group were significantly lower than those in model group (P<0.01,P<0.05),and its AChE content was lower than that in the dysosmia group (P<0.05).The content of Glu in dysosmia treated with moxa smoke group was between the model group and the moxa smoke group,and there was significant difference among the 3 groups (P<0.05).Conclusion:With certain concentration,moxa smoke intervention can benignly regulate the content of neurotransmitters in the olfactory bulb.Through the neural projection of the olfactory pathway,the expression of neurotransmitters in the brain can be interfered and the oxidative stress could be improved.
Key Words Moxa smoke; Olfactory bulb; Neurotransmitter; Olfactory pathway; Glutamate acid; Gamma-aminobutyric; Acetylcholinesterase
中图分类号:R242文献标识码:Adoi:10.3969/j.issn.1673-7202.2019.05.017
艾叶作为传统灸材,其燃烧产生的烟雾不但在艾灸过程中发挥重要作用,更可单独应用治疗多种疾病,自古以来便备受推崇。早在《五十二病方》中就记载了以干艾、柳蕈熏燃外治的病案;《肘后备急方》有“以干艾斛许,揉团纳瓦甑中,并独留一目,以痛处著甑目而烧艾熏之”的烟熏操作法;《本草纲目》中亦有“熟艾烧烟薰之……治风虫牙痛”的阐述。现代药理学研究发现,艾叶燃烧生成的艾烟含有多种具有抗氧化、抗炎、镇静作用的芳香物质[1],其中许多成分亦常见见于的芳香开窍药物或精油[2]。部分已被证实是通过嗅觉通路发挥保护脑神经功能[3],改善学习记忆的作用[4]。本前期研究发现,单纯艾烟干预可以治疗性提高ApoE-/-及SAMP8小鼠脑内单胺类神经递质含量,减轻氧化应激,延缓脑组织衰老[5-6]。但艾烟究竟通过何种途径发挥作用至今尚不明确,可能是通过血脑屏障进入中枢神经系统,也可能经嗅神经传导通路起效。
本实验选用快速老化模型小鼠,通过3-甲基吲哚腹腔注射毁损嗅黏膜造模[7],旨在保持嗅觉中枢完整性的情况下,观察单纯艾烟干预对小鼠嗅球形态以及嗅球内神经递质含量的影响,初步探讨艾烟起效途径与嗅觉通路相关性,为进一步探索艾烟在艾灸整体效应机制中的意义奠定基础。
1 材料与方法
1.1 材料
1.1.1 动物 选取SPF级6个月龄雄性SAM小鼠60只,其中SAMP8系48只,同品系正常老化小鼠SAMR1系12只,平均体质量(30±2.3)g,购自中科泽晟科技有限公司[SCXK(京)2014-0011]。采用标准动物饲料喂养,自由饮水;饲养环境温度(22±2)℃,湿度50%~60%。采用人工控制室内照明,12/12 h光暗环境交替循环。
1.1.2 药物
特制细艾条(Φ0.5 cm×20 cm,河南南阳汉医艾绒有限公司);玉米油(上海源叶生物科技有限公司)。
1.1.3 试剂及仪器 3-甲基吲哚(3-MI,99%,北京银河天虹化工有限公司);自制艾烟干预箱(75 cm×45 cm×60 cm);P5L2C光散射式数字粉尘测试仪(北京宾达绿创科技有限公司);磷酸缓冲盐溶液(PBS)(Coolaber,北京华菁美达科技有限公司);4%多聚甲醛(Coolaber,北京华菁美达科技有限公司);石蜡包埋机(EG1150,德国Leica公司);石蜡切片机(RM2235,德国Leica公司);谷氨酸(Glu)、γ-氨基丁酸(GaBa)和乙酰胆碱酯酶(AChE)试剂盒均购于上海酶联生物研究所;Multiskan MK3标准规格酶标仪(Thermo Scientific)。
1.2 方法
1.2.1 分组及模型制备 选取48只SAMP8小鼠并用随机数字表法分为艾烟组,嗅觉障碍+艾烟组(嗅障艾烟组),嗅觉障碍组,模型组,每组12只。12只SAMR1雄性小鼠作為空白对照组对照。适应性饲养2周后进行造模。将3-MI与玉米油混合溶解,制成30 mg/mL的溶液,按照300 mg/kg剂量对嗅障艾烟组及嗅觉障碍组小鼠进行腹腔注射。其余组依据体质量注射同等剂量玉米油。
给药24 h后行埋藏食物小球实验(Buried Food Pellet Test,BFPT)。小鼠从测试的前24 h开始限食(2.4 g/d),自由饮水,测试时将嗅觉行为测试盒内铺入洁净的玉米垫料,厚度约4 cm。将1 g重的食物小球埋藏在垫料下约2 cm深处,每次测试时将小鼠从相同位置放入测试盒中。从小鼠放入测试盒中开始计时,小鼠前爪或牙齿抓住食物小球时停止计时,将这段时间定义为小鼠找到食物小球的时间。小鼠找到的食料允许其在测试中吃完。每日同一时间重复实验,连续3 d。以BFPT实验300 s(3次测试的平均值)内未找到饲料作为嗅觉障碍造模成功。
1.2.2 干预方法 根据前期实验以及临床诊室测量数据,本实验中干预用艾烟干预条件定为5.0~15.0 mg/m3,以模拟临床诊室中艾烟浓度。实验开始前将光散射式数字粉尘测试仪置于箱体下部,细艾条点燃插入底座并固定于艾烟吸嗅箱隔板中部,箱壁及隔板均分布通气孔,可保证烟气的充分弥散。艾条于箱内点燃1 min后,将小鼠逐一放入箱内,每只小鼠由隔板隔开,避免互相干扰。艾烟干预过程中,每5 min记录一次浓度值,若数值高于15 mg/m3,可移动上方插板调整通气口。实验过程中艾烟浓度基本维持在10.5~12.8 mg/m3。艾烟组和嗅障艾烟组小鼠每日接受艾烟干预30 min,共6周。嗅觉障碍组、模型组及空白对照组不特殊处理,给予正常空气吸嗅。
实验末次干预结束后,禁食不禁水12 h。每组随机选取3~4只小鼠[艾烟组和模型组取3只(后期各脱落1只),其余组取4只]腹腔注射10%水合氯醛(0.3 mL/100 g)进行麻醉,开胸暴露心脏,将灌流针头经左心室插入主动脉,PBS快速冲洗并灌注4%多聚甲醛。断头开颅,参照小鼠解剖图谱,取出嗅球,置于4%多聚甲醛中固定。其余小鼠处死后,于冰台上快速取出嗅球组织,置于标记的2 mL冻存管中,液氮冻存数分钟后移入-80 ℃冰箱待测。
1.2.3 检测指标与方法 采用苏木精-伊红(HE)染色法观察各组其余8只小鼠嗅球组织形态结构,小鼠嗅球固定7 d后常规石蜡包埋、切片,经烤片、梯度乙醇和二甲苯脱蜡至水、染色、脱水封片后,于光学显微镜下观察嗅球形态,采集图像并分析。采用酶联免疫吸附试验法检测嗅球内Glu、GaBa、AChE含量,制备10%浓度的小鼠嗅球组织匀浆,检测时严格按照试剂盒说明进行操作。
1.3 统计学方法
采用SPSS 20.0统计软件分析数据。计量资料符合正态性分布且通过方差齐性检验,以均值±标准差(±s)表示,采用单因素方差分析,并用LSD法进行组间比较,以P<0.05为差异有统计学意义。
2 结果
2.1 各组小鼠嗅球HE染色结果比较
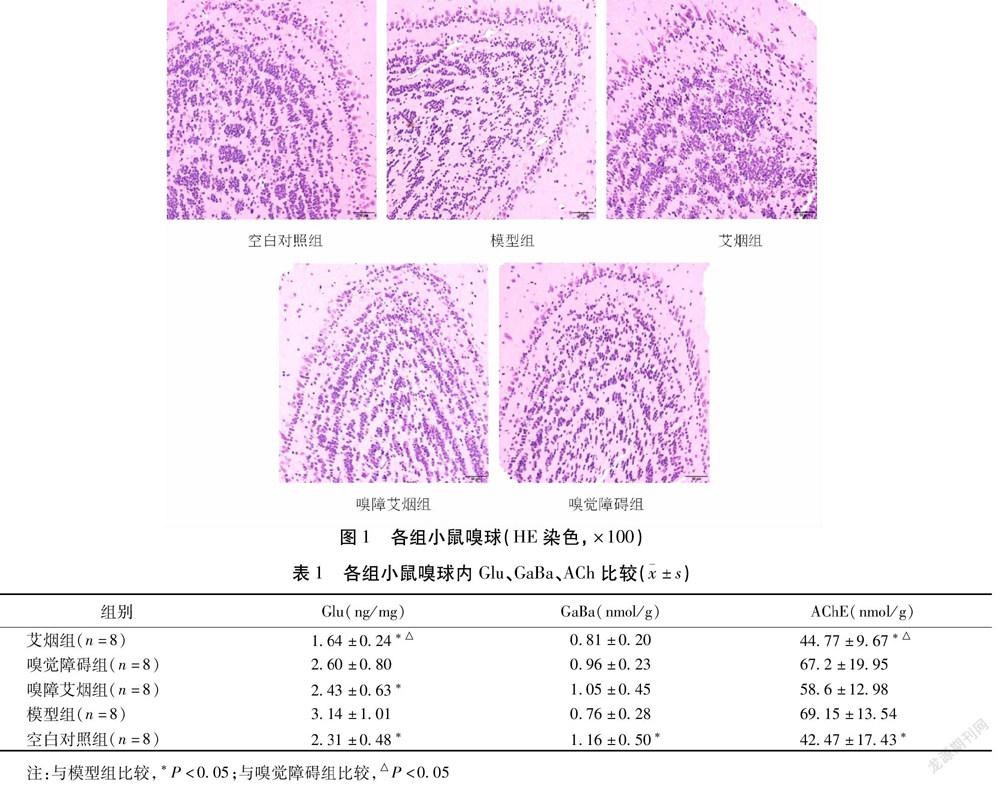
光镜下可观察到空白对照组小鼠嗅球结构清晰,呈同心圆状规则排列,自内向外依次可观察到颗粒细胞层、僧帽细胞层、外丛层、突触小球层;空白对照组小鼠嗅球僧帽细胞胞质红染,呈圆形或卵圆形,细胞核大,树突明显;颗粒细胞核仁清晰,胞核深染,呈串珠样簇集。其余组小鼠嗅球细胞整体结构未见明显差异,但僧帽细胞层神经元数量比较空白对照组略有减少;模型组、嗅觉障碍组及嗅障艾烟组颗粒细胞层细胞数量和密度稍有降低。艾烟组部分僧帽细胞楔形,突起明显;且颗粒细胞层细胞数量及密度明显高于模型组、嗅觉障碍组及嗅障艾烟组。见图1。
2.2 各组小鼠嗅球内Glu、GaBa、AChE含量比较
空白对照组、艾烟组及嗅障艾烟组嗅球内Glu含量均低于模型组,差异有统计学意义(P<0.05),且艾烟组较嗅障艾烟组显著降低,差异有统计学意义(P<0.05)。嗅觉障碍组与模型组比较,差异无统计学意义(P>0.05)。
嗅球内GaBa含量空白对照组显著高于模型组,差异有统计学意义(P<0.05),其余各组比较,差异无统计学意义(P>0.05)。
空白对照组与艾烟组嗅球AChE含量均显著低于模型组,差异有统计学意义(P<0.05)。艾烟组AChE含量显著低于嗅觉障碍组,差异有统计学意义(P<0.05)。但与嗅障艾烟比较,差异无统计学意义(P>0.05)。嗅觉障碍组、嗅障艾烟组与模型组比较,差异无统计学意义(P>0.05)。见表1。
3 讨论
嗅觉系统是感觉神经系统的重要组成部分,其发育和调控与整个机体的生长和发育有着紧密的联系,对情绪、睡眠、记忆等大脑高级活动有重要影响。嗅觉通路由嗅上皮(OE)、嗅球(OB)和嗅觉皮层(OC)3级结构组成,通过逐级信号传递实现对气味的识别和认知。嗅觉刺激可以不经丘脑中继直接到达中枢,具有起效迅速且高度敏感的特点。同时由于嗅觉神经元对受体的特异性表达,气味的识别和传导具有高度特异性[8]。此外,嗅觉通路与神经退行性病变关系密切,且给药生物利用率高,吸收快,不良反应小。因此近年来成为中枢系统疾病研究热点之一。
嗅球是嗅觉通路的第一中转站,在嗅觉的产生及嗅觉与高级活动的联系中发挥至关重要的作用。嗅球的病理改变可引起学习记忆行为的改变和相关中枢神经生化活动的改变。近年来发现,帕金森病、阿尔茨海默病等神经退行性疾病多伴有嗅觉功能减退以及嗅觉系统病变[9]。阿尔茨海默患者在疾病早期即可出现嗅球病变,其改变甚至早于皮层[10]。此外,动物实验发现单纯损毁嗅球即可一定程度降低正常大鼠学习记忆力[11]。吴子建[12]发现艾烟和艾油的芳香气味分子可以诱发嗅球动作电位。不同浓度艾烟还可以引起嗅球超微结构及c-Fos蛋白表达的改变[13]。因此推测嗅球结构与功能可能与艾烟改善SAMP8小鼠学习记忆的效应具有一定关联性。本实验结果显示,各组小鼠嗅球整体结构并无明显差异,3-MI诱导的嗅觉障碍模型嗅觉障碍仅局限于外周感受器,嗅球并未出现明显病变,与相关报道一致[14]。嗅球在机体成年期依旧能持续不断地生成以球周细胞和颗粒细胞为主的中间神经元。颗粒细胞是嗅球内最多的一种中间神经元,参与处理各种气味信息。嗅觉剥夺的成鼠嗅球可观察到颗粒细胞明显减少[15],而丰富的嗅觉经验则有助于新生细胞的增加[16]。艾烟组颗粒细胞密度较模型组有所增加,而嗅障艾烟组未表现出相同变化,可能是由于艾烟的气味刺激促进了新生神经细胞的成活和分化。
Glu和GaBa是嗅觉系统中最主要的2种神经递质。Glu作为兴奋性神经递质,它不但参与学习记忆形成和信息贮存的过程,还与神经系统的突触形成、重塑及退化密切相关。但脑内Glu的过度蓄积可引起神经细胞损伤[17]。GaBa是抑制性神经递质,对神经干细胞的增殖、分化和神經元的定向迁移具有调节作用[18-19]。此外,乙酰胆碱也参与对嗅觉系统可塑性和感知力的调节[20]。AChE是乙酰胆碱的水解酶,可通过调节乙酰胆碱含量控制突触间信息传递,对反映胆碱能神经元活性有重要意义。实验结果显示,模型组Glu和AChE显著高于空白对照组,提示SAMP8嗅球内也存在兴奋性神经递质的过度蓄积和胆碱酯酶活性的升高。GaBa含量空白对照组显著高于模型组,其余组各组无明显差异,可能兴奋性递质在嗅觉活动中的作用较抑制性递质更为重要,因此变化更加敏感。也可能艾烟干预对嗅球内GaBa递质的合成和释放影响较小,或受限于艾烟浓度、时间因素未对GaBa代谢产生作用。艾烟组Glu显著低于模型组和嗅障艾烟组,AChE含量显著低于模型组和嗅觉障碍组,说明单纯艾烟干预可以下调嗅球内过高的谷氨酸和胆碱酯酶的异常代谢,改善氧化应激,且这种效应依赖于嗅觉通路的完整性。此外,嗅障艾烟组Glu较模型组也有明显降低,考虑可能是艾烟通过黏膜、皮肤等途径发挥的作用,或是艾烟的气味刺激加快了嗅觉障碍的恢复从而产生一定效应。
综上所述,艾烟治疗作用的发挥与嗅觉通路具有一定相关性。基于对嗅觉系统结构和工作方式的理解,艾烟的芳香气味刺激可能是通过调节嗅球内神经递质,提高嗅觉系统神经兴奋性,并借助神经联系影响大脑各区域的神经递质表达,改善脑内氧化应激状态。
参考文献
[1]吴子建,王斌,段文秀,等.顶空进样-气相色谱-质谱联用法检测3年陈艾条燃烧产物中挥发性成分[J].安徽中医药大学学报,2017,36(2):64-67.
[2]吴蒙,徐晓军.迷迭香化学成分及药理作用最新研究进展[J].生物质化学工程,2016,50(3):51-57.
[3]张新渐,韦德勇,王洪云,等.丁香酚生物活性研究进展[J].中国中医药现代远程教育,2017,15(24):157-160.
[4]贺利敏,李光武,王敏,等.辛夷挥发油经嗅觉通路改善自闭症模型昆明鼠学习记忆能力及其神经递质含量[J].神经解剖学杂志,2016,32(4):507-512.
[5]王昊,杨佳,赵百孝,等.艾燃烧生成物对ApoE基因敲除小鼠脑内神经递质5-HT、GABA的影响[J].世界中医药,2016,11(8):1410-1413.
[6]孟笑男,许焕芳,崔莹雪,等.艾燃烧生成物对快速老化模型小鼠P8大脑单胺类神经递质含量的影响[J].中华中医药杂志,2014,29(9):2761-2763.
[7]李丽娟,李金红,毛兰群,等.化学性嗅觉障碍模型中嗅觉诱发电位的研究[J].中国耳鼻咽喉头颈外科,2014,21(4):209-212.
[8]Touhara K.Odor discrimination by G protein-coupled olfactory receptors[J].Microsc Res Tech,2002,58(3):135-141.
[9]周小燕,王晓明.神经退行性疾病的嗅觉障碍[J].中华临床医师杂志(连续型电子期刊),2016,10(16):2488-2491.
[10]Kovács T,Cairns NJ,Lantos PL.Olfactory centres in Alzheimer′s disease:olfactory bulb is involved in early Braak′s stages[J].Neuroreport,2001,12(2):285-288.
[11]劉智斌,牛文民,杨晓航,等.损毁嗅球对痴呆大鼠学习记忆功能影响作用的实验研究[J].陕西医学杂志,2009,38(1):17-19.
[12]吴子建.艾烟熏灸对膝骨性关节炎的影响以及嗅觉通路参与艾烟作用的机理研究[D].合肥:安徽中医药大学,2017.
[13]吴子建.不同环境浓度艾烟吸入对大鼠嗅球超微结构和c-Fos蛋白表达的影响[C].北京:2017世界针灸学术大会暨2017中国针灸学会年会论文集,2017.
[14]Peele DB,Allison SD,Bolon B,et al.Functional deficits produced by 3-methylindole-induced olfactory mucosal damage revealed by a simple olfactory learning task[J].Toxicol Appl Pharmacol,1991,107(2):191-202.
[15]Mandairon N,Sacquet J,Jourdan F,et al.Long-term fate and distribution of newborn cells in the adult mouse olfactory bulb:Influences of olfactory deprivation[J].Neuroscience,2006,141(1):443-451.
[16]Rochefort C,Gheusi G,Vincent JD,et al.Enriched odor exposure increases the number of newborn neurons in the adult olfactory bulb and improves odor memory[J].J Neurosci,2002,22(7):2679-2689.
[17]胡捷先.脑缺血后谷氨酸通路及其调控的研究进展[J].复旦学报:医学版,2016,43(6):724-731.
[18]Carleton A,Petreanu LT,Lansford R,et al.Becoming a new neuron in the adult olfactory bulb[J].Nature Neuroscience,2003,6(5):507-518.
[19]Bolteus AJ,Bordey A.GABA release and uptake regulate neuronal precursor migration in the postnatal subventricular zone[J].J Neurosci,2004,24(35):7623-7631.
[20]宫田田.乙酰胆碱对小鼠嗅球神经环路的调控作用[D].桂林:桂林医学院,2016.
猜你喜欢
右江医学(2022年3期)2022-04-17
河南农业·综合版(2022年1期)2022-03-01
大众健康(2021年2期)2021-03-09
中国药学药品知识仓库(2021年18期)2021-02-28
分析化学(2019年3期)2019-03-30
农业与技术(2019年3期)2019-03-13
江苏农业科学(2017年12期)2017-09-16
中国中药杂志(2017年4期)2017-03-28
中国实用医药(2016年22期)2016-08-19
中国民族民间医药·下半月(2016年3期)2016-04-19
