
光合电子流对光响应机理模型在超级晚稻光合特性研究中的应用
2019-09-10 07:22贺俐贺晓鹏边建民朱昌兰傅军如吴杨贺浩华
南方农业学报 2019年9期
关键词:光合特性
贺俐 贺晓鹏 边建民 朱昌兰 傅军如 吴杨 贺浩华
摘要:【目的】了解光合电子流对超级晚稻光合特性的影响,进一步揭示植物捕光色素分子内禀参数对超级晚稻光合作用的影响机制,为超级晚稻高光效育种打下基础。【方法】利用LI-6400XT便携式光合测定系统测量不同超级晚稻品种的电子传递速率对光的响应曲线,利用叶氏光合机理模型分析超级晚稻光合特性差异及其产生的原因。【结果】叶氏光合机理模型可较好地拟合超级晚稻叶片的光合电子流对光的响应曲线,由此得到的最大电子传递速率(ETRmax)和饱和光强(PARsat)与实测值高度符合。超级晚稻品种的ETRmax和PARsat均显著高于对照汕优46(P<0.05),超级晚稻本征光能吸收截面(σik)较大,捕光色素分子处于最低激发态的最小平均寿命(τmin)较短。此外,捕光色素分子的有效光能吸收截面(σ´ik)和处于激发态的捕光色素分子(Nk)也影響着超级晚稻对光能的吸收和利用。【结论】超级晚稻通过捕光色素分子内禀参数影响光合电子流,进一步影响其光合特性。
关键词: 光合特性;光合电子传递速率;捕光色素;超级晚稻
中图分类号: S311 文献标志码: A 文章编号:2095-1191(2019)09-1937-08
Abstract:【Objective】The purpose of the study was to understand the effects of photosynthetic electron flow on photosynthetic characteristics of super late rice,and reveal how the intrinsic parameters of plant light-harvesting pigment mo-lecules affected photosynthesis of super late rice, and laid a foundation for breeding high photosynthetic efficiency varie-ties of rice. 【Method】The light-response curves of photosynthetic electrons transfer rate to light of different super late rice varieties were measured by LI-6400XT portable photosynthesis measurement system. The differences in the photosynthe-tic characteristics and their courses were studied based on Ye model of light-response curves of photosynthetic electron flow. 【Result】The Ye model could fit the response curve of photosynthetic electron flow of super late rice to light well, and the obtained fitted maximum electron transport rate(ETRmax) and saturated light intensity(PARsat) were highly consistent to the measured values. The maximum electron transport rate(ETRmax) and maximum electron transport rate(PARsat) of super late rice were significantly higher than those of control Shanyou 46(P<0.05),the super late rice had the greater eigen-absorption cross-section of light-harvesting pigments and the shorter minimum average lifetime(τmin) of photosynthetic pigment molecules in the excited state. Moreover,the effective light absorption cross-section(σ'ik) of light-harvesting pigment molecules and the light-harvesting pigment molecules(Nk) in the excited state were beneficial to absorption and utilization of light energy in super late rice. 【Conclusion】Super late rice affects the photosynthetic electron flow by the characteristics of light-harvesting pigment molecules,further influence its photosynthetic characteristics.
Key words: photosynthetic characteristics; photosynthetic electron transport rate; light-harvesting pigment; super late rice
0 引言
【研究意义】水稻(Oryza sativa L.)是世界上的主要粮食作物之一,大幅度提高水稻单产及超级稻的培育和推广被认为是现阶段实现粮食增产的最有效途径(Kathuria et al.,2007;吴俊等,2016)。光合作用是影响作物生产力高低的关键因素(沈允钢等,1998),当前对水稻光合特性的研究多为环境因素(董桃杏等,2009;赵森等,2013;Wang et al.,2017;周根友等,2018)和不同耕作方式(林诚等,2016;蔡威威等,2018)对水稻光合作用的影响,或不同水稻品种光合特性差异的比较(杨旺兴等,2017;宋微等,2017),这些研究主要集中在光合生理指标如饱和光强(Isat)、净光合速率(Pn)、PS Ⅱ的有效光量子效率(ΦPSⅡ)等的测定,极少涉及光合電子传递速率。植物叶片中光合电子传递速率越快,形成光合能力就越强。光合电子流不仅是植物光合能力形成的根本,还是光合碳同化进程的能量源泉(Archontoulis et al.,2012;叶子飘等,2014;Gao et al.,2016)。因此,从光合电子流的角度研究超级晚稻的光合特性差异及其产生的原因对超级晚稻的高光效育种具有重要意义。【前人研究进展】植物电子传递相关参数如最大电子传递速率(ETRmax)、饱和光强(PARsat)、初始斜率(αe)等需通过光合模型进行拟合计算(闫小红等,2016)。不同模型的拟合效果差异较大,对于参数的拟合计算也非常关键。目前,植物光合电子流对光响应模型主要分为经验模型和半机理模型两大类(叶子飘等,2014)。经验模型未涉及电子传递、光合原初反应等过程,且不能为光抑制现象提出合理解释,无法直接计算光饱和点(叶子飘等,2018a)。半机理模型虽将光能的吸收和激发引入到模型,但未涉及电子的共振传递和转化等过程,同样不涉及捕光色素分子内禀参数在这些过程中的作用,无法计算ETRmax和PARsat等光合参数(Suggett et al.,2004;Brading et al.,2011)。鉴于此,在充分考虑光合作用原初反应过程和捕光色素分子光能吸收特性的基础上,Ye等(2013a)建立了一个光合电子流对光响应的机理模型(下称叶氏光合机理模型),目前该模型已被国内外学者广泛应用。Xu等(2014)利用叶氏光合机理模型对不同施氮量下的水稻光合特性进行研究。Li等(2015)利用光合机理模型对冷处理下的小麦光合特性进行对比分析。胡文海等(2017)采用光合机理模型研究了越冬期广玉兰阳生叶和阴生叶两种不同光环境下叶片PSⅡ功能及其捕光色素分子内禀特性的差异,结果表明阳生叶可通过降低叶绿素含量和捕光色素分子数量以减少对光能的吸收,而阴生叶热耗散能力较强,可有效保护自身免受短时曝露在强光下的伤害。康华靖等(2018)利用叶氏光合机理模型对蓝莓叶片饱和光强进行研究,结果表明,直角双曲线修正模型能较好拟合光合速率和电子传递速率对光的响应曲线,其光饱和点值与实测值接近;而直角双曲线模型、非直角双曲线模型及指数方程模型的拟合结果均远低于实测值。商天其和孙志鸿(2019)利用叶氏光合机理模型对香樟幼龄林不同叶龄叶片的光合特征进行研究,为单萜释放模型区域性BVOCs释放模型提供了参考。【本研究切入点】植物叶片捕光色素分子对光能吸收、传递和电荷分离等过程与光合作用的原初反应密切相关(Govindjee,2002;Nelson and Yocum,2006)。在正常环境下植物的光合作用是由植物本身的内禀特性所决定(叶子飘等,2016)。原初反应过程中形成的光合电子流是植物光合能力形成的重要基础,然而针对植物叶片中哪些内禀特性会对电子传递速率产生影响并进一步影响其光合能力的相关研究鲜有报道。【拟解决的关键问题】从光合电子流对光响应的机理模型出发(Ye et al.,2013a),研究相同环境条件下超级晚稻的光合特性,探索超级晚稻叶片的光合电子流对其光合特性的影响,进一步揭示植物捕光色素分子内禀参数对超级晚稻光合作用的影响机制,为超级晚稻高光效育种打下基础。
1 材料与方法
1. 1 试验地概况及供试材料
试验于2017年6—10月在江西省吉安市青原区雷家村(东经115°02′,北纬27°11′)双季稻水稻生产田内进行。试验地年平均气温18.2 ℃,≥10.0 ℃全年活动积温5600~5700 ℃;全年≥10.0 ℃积温天数260~300 d;年平均降水量1426 mm,年平均日照时数1782 h。土壤理化性质:有机质2.38 g/kg,全氮1.06 g/kg,碱解氮140.08 mg/kg,有效磷14.68 mg/kg,速效钾65.23 mg/kg,pH 5.58。
供试水稻品种分别为淦鑫688(分蘖力强,有效穗较多,穗粒数多,着粒密,结实率较高)、五丰优T025(分蘖力强,有效穗较多,穗粒数多,着粒密,结实率较高,千粒重小)、天丰优101(分蘖力强,有效穗多,穗粒数中,结实率较高)、天优998(分蘖力一般,有效穗偏少,穗粒数较多,结实率高,千粒重大)、五优308(分蘖力中,有效穗多,穗大粒多)和荣优225(分蘖力一般,有效穗中,穗粒数多,结实率较高),对照品种为汕优46,均由江西省作物生理生态与遗传育种重点实验室提供。
1. 2 试验方法
试验采用随机区组设计,以品种为处理,小区面积16 m2,3次重复。大田湿润育秧,6月20日播种,7月18日移栽,行株距26.4 cm×15.0 cm,基本苗2株/穴。施纯N 195 kg/ha,P2O5 105 kg/ha,K2O 180 kg/ha。氮肥施用比例为基肥∶分孽肥∶穗肥=5∶1∶4;磷肥全部作为基肥;钾肥施用比例为基肥∶分孽肥∶穗肥=5∶1∶4。氮、磷、钾肥分别为尿素、钙镁磷肥和氯化钾。移栽前2 d施用基肥,移栽后5~7 d施用分孽肥,叶龄余数2左右施用穗肥,其余田间管理措施同一般高产田。因水稻穗期的光合作用对后期产量影响较大,故在孕穗期(9月2日)晴朗天气下,随机选取3株长势均匀一致的健壮植株,每株选择3片剑叶进行光合参数的活体测定。
1. 3 光合电子流对光响应数据测定
在晴天上午8:00—12:00,利用LI-6400-40B自带光源,设自动测量程序进行数据测量,利用CO2钢瓶控制CO2泵流速为500 mmol/s,浓度为390±2 μmol/mol,相对湿度控制在50%~70%,叶室温度设定为(35±1)℃。测试叶片在不同光强下,每隔180 s记录一个点。光合有效辐射梯度设2000、1800、1600、1400、1200、1000、800、600、400、200、100、50和0 μmol/(m2·s),测定得到电子传递速率(Electron transport rate,ETR)等光合参数。利用Ye等(2013a)的光合电子流对光响应机理模型对光合电子流响应曲线进行拟合,得到ETRmax、PARsat和αe等。
1. 4 ETRmax和PARsat计算
植物叶片的光合电子流对光响应机理模型(Ye et al.,2013a)为:
模型中,ae为初始斜率(μmol/μmol),表示光能转化为光合电子流的最大能力;[βe]为植物叶片PSⅡ动力学下调系数(m2·s/μmol);[γe]为饱和系数(m2·s/μmol);I为光照强度[μmol/(m2·s)]。由下列公式可得到植物叶片的PARsat和相对应的ETRmax:
1. 5 捕光色素分子内禀参数计算
捕光色素分子有效光能吸收截面通过以下公式计算(Ye et al.,2013b):
式中,σ´ik为捕光色素分子的有效光能吸收截面,随着光照强度的增加,σ´ik呈非线性下降。
处于最低激发态的捕光色素分子数与光强的关系为:
式中,Nk为处于最低激发态的捕光色素分子数,gi和gk分别为捕光色素分子处于基态i和激发态k的简并度(无量纲),N0为捕光色素分子数,N0=(叶绿素含量×实测面积×10-3/叶绿素分子量)×6.02×1023,叶绿素分子量900 g/mol。
捕光色素分子的本征光能吸收截面的估算:
1. 6 叶绿素含量测定
取光合参数测定的水稻植株进行叶绿素含量测定,具体操作参照周祖富和黎兆安(2005)的方法。
1. 7 统计分析
采用光合计算软件(http://photosynthetic.sinaapp.com/calc.html)对测量数据进行分析。利用SPSS 22.0对试验数据进行统计分析,并以OriginPro 8.5制图。
2 结果与分析
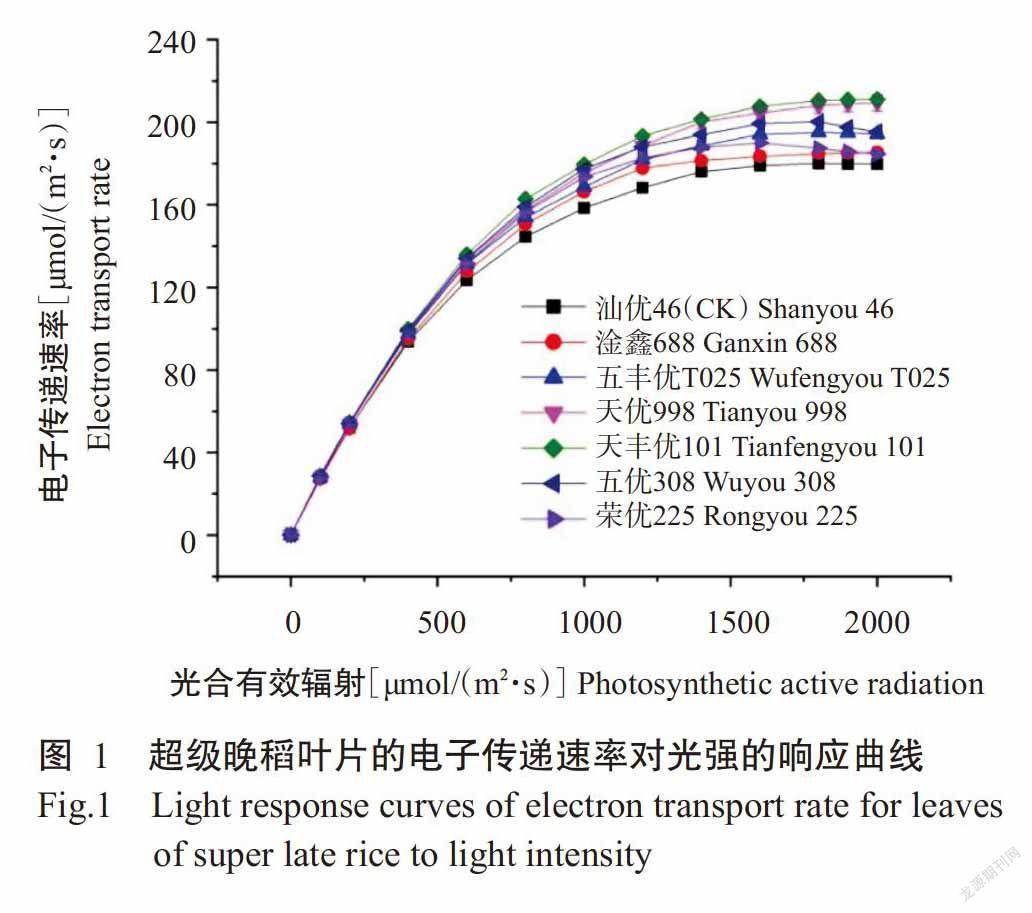
2. 1 电子传递速率对光强的响应曲线
由图1可看出,Ye等(2013a)建立的光合电子流对光响应机理模型很好地拟合了不同光合有效辐射下水稻叶片的ETR-I曲线,所有晚稻品种的叶片在光合有效辐射低于400 μmol/(m2·s)时,电子传递速率随光合有效辐射的增加几乎呈直线增长;随着光合有效辐射的增强,所有水稻品种的电子传递速率缓慢增长,且对照汕优46的电子传递速率始终低于各超级晚稻品种。达饱和光强后,除淦鑫688、天优998和天丰优101的叶片电子传递速率未出现下调趋势外,其他晚稻品种均发生不同程度的PSⅡ的动力学下调(White and Critchley,1999)。与对照相比,超级晚稻保持着较高的电子传递速率,具有更强的光合能力。
由光合电子流对光响应的机理模型拟合得到αe、PARsat和ETRmax等光合参数(表1)。由表1可知,ETRmax和PARsat的拟合值与测量值非常接近,决定系数(R2)均超过0.9990。超级晚稻五丰优T025和天丰优101的初始斜率αe最大,为0.318 μmol/μmol,说明这两个超级晚稻品种将光能转化为电子流的能力最强,其他超级晚稻品种的初始斜率αe与对照汕优46相比无显著差异(P>0.05,下同)。所有超级晚稻品种的ETRmax和PARsat均显著高于对照汕优46(P<0.05,下同),其中天丰优101的ETRmax最大,为210.90±1.78 μmol/(m2·s),天优998的PARsat最大,为1926.67±9.46 μmol/(m2·s)。
2. 2 光合叶绿素含量及捕光色素分子内禀特性的比较
由表2可知,不同晚稻品种间的叶绿素含量存在明显差异,其中,对照汕优46的叶绿素含量最高,为576.00±8.50 mg/m2,五丰优T025的叶绿素含量最低,为408.00±15.10 mg/m2,超级晚稻的叶绿素含量显著低于对照汕优46。除ETRmax和PARsat等光合参数外,通过光合电子流对光响应机理模型还可估算捕光色素分子的一些物理参数,如捕光色素分子数(N0)、处于最低激发态的最小平均寿命(τmin)、本征光能吸收截面(σik)等。由表2可知,超级晚稻叶片中N0的变化情况与叶绿素含量相似,超级晚稻的N0显著低于对照汕优46。对照汕优46的叶绿素含量最高,但其捕光色素分子的σik最小,为(2.13±0.05)×10-21 m2;五丰优T025的叶绿素含量最低,但其捕光色素分子的σik最大,为(3.10±0.04)×10-21 m2;荣优225的σik与对照汕优46相比无显著差异,其他超级晚稻品种的σik显著高于对照汕优46。对照汕优46捕光色素分子的τmin最长,为93.24±0.26 ms,超级晚稻品种的τmin均显著短于对照汕优46。
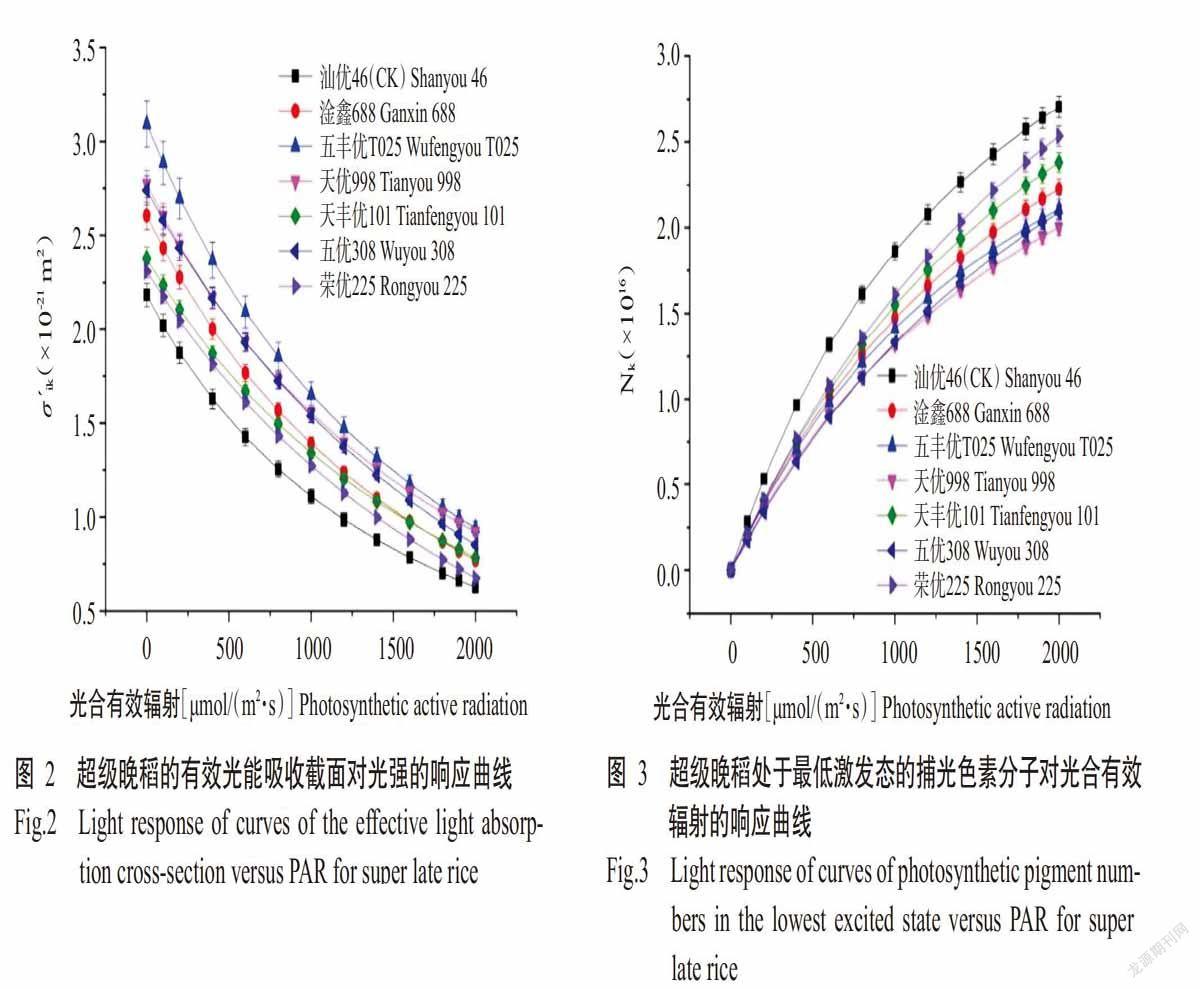
2. 3 不同光照强度下的捕光色素分子有效光能吸收截面
光能吸收截面能反映植物捕光色素分子对光能吸收能力的强弱,捕光色素分子有效光能吸收截面(σ´ik)则表示其在不同光强下捕获光子能力的强弱(Ye et al.,2013b)。由图2可知,各晚稻品种的σ´ik随光合有效辐射的增加呈非线性下降趋势,表明各晚稻品种的捕光色素分子吸收光能的能力随光合有效辐射的增加而下降。但不同光合有效辐射下,各超级晚稻品种的σ´ik始终高于对照汕优46,在光能的吸收能力上显示出一定优势。在高光强条件下,σ´ik最小的是对照汕优46,表示在高强光下超级稻的捕光色素分子吸收光能的能力依然很强,超级晚稻中五丰优T025的σ´ik最大;在低光强条件下,σ´ik最大的依然是五丰优T025,表示其在低光强条件下的光能吸收能力最强。上述结果表明,与对照汕优46相比,超级晚稻在各光强条件下可保持更高的有效光能吸收截面,表现出更強的光吸收能力。从整个光照过程来看,这种表现在低光照下更明显,在高光强条件下,超级晚稻的σ´ik依然大于对照汕优46,但差异不明显。
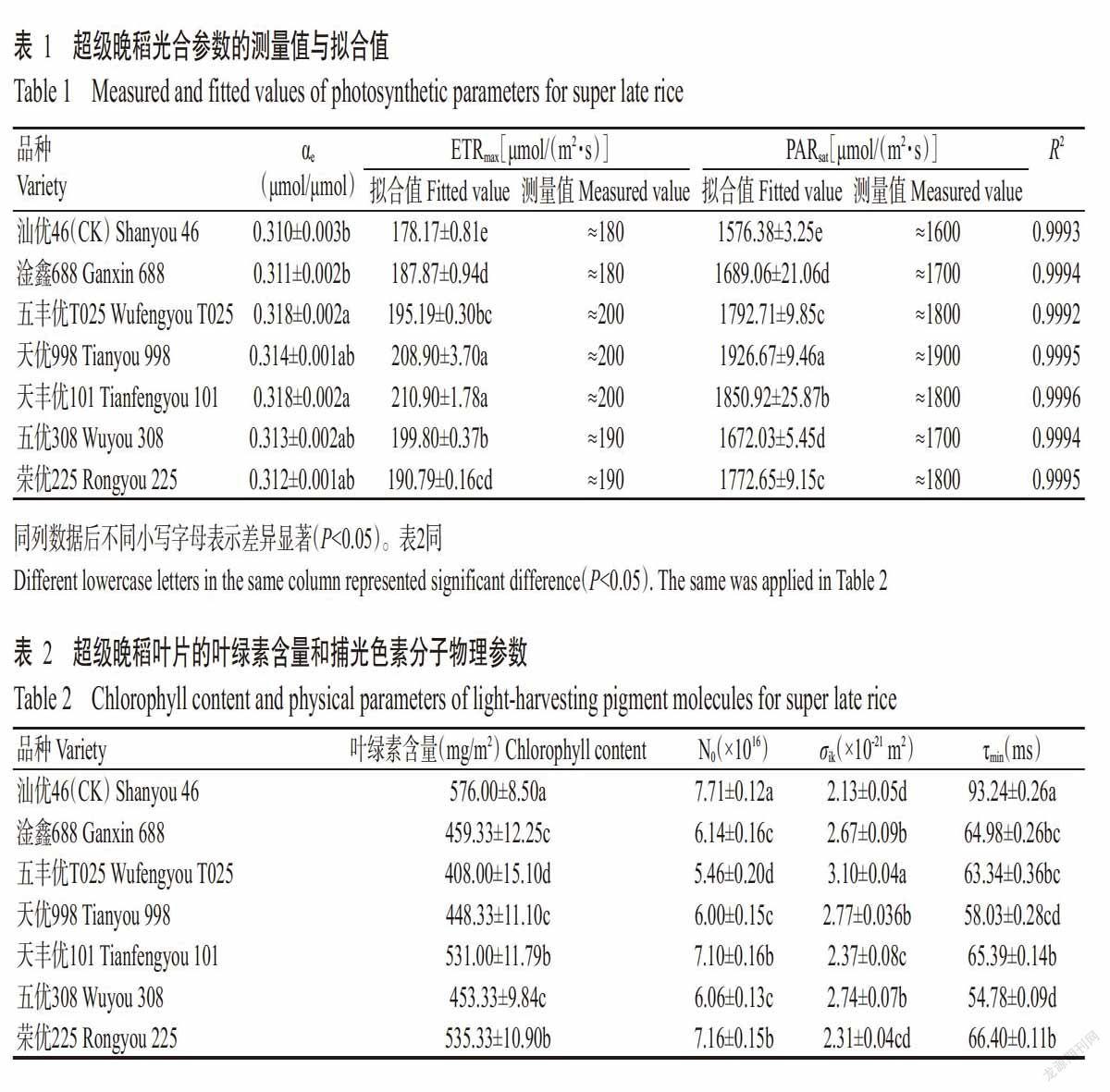
2. 4 处于最低激发态的捕光色素分子数对光合有效辐射的响应曲线
随着光合有效辐射的不断增加,所有晚稻品种的最低激发态的捕光色素分子数(Nk)呈现非线性增加趋势,一旦处于激发态的捕光色素分子不能及时回到基态,这些激发态的捕光色素分子将对植物的光合器官产生伤害,从而影响植物光能吸收和利用(Takahashi and Badger,2011;Niyogi and Truong,2013)。由图3可知,各超级晚稻品种的Nk上升速度均慢于对照汕优46,特别是在高光强处,对照汕优46的Nk最大,超级晚稻天优998的Nk最小,各超级晚稻品种的Nk均低于对照汕优46。以上结果表明,超级晚稻叶片中用于光能吸收的捕光色素分子数相对较多,在一定程度上可减轻强光对水稻叶片光抑制伤害的风险,更有利于超级稻对光能的吸收利用。
3 讨论
本研究结果显示,随着光照强度的增加,超级晚稻的电子传递速率快速增加,在达饱和光强时,电子传递速率达峰值;之后,电子传递速率随光强的增加发生光抑制现象,可能与捕光色素分子有效光能吸收截面(σ´ik)随光强增加而下降有关(叶子飘等,2018b);也有个别品种随着光照强度的增加其电子传递速率基本不变,出现光饱和现象。在不同光照强度下,特别是达饱和光强后,超级稻的电子传递速率均高于对照汕优46,说明超级稻有着更强的光合能力。此外,利用叶氏光合机理模型得到的ETRmax和PARsat拟合值与实测值非常接近,与前人的研究报道相符。叶子飘等(2014)在研究山莴苣、一年蓬和紫菀叶片光合电子流时发现,光合电子流对光响应的曲线呈现典型的C3植物曲线特点,成功拟合得到ETRmax和PARsat等光合参数,并与实测值几乎一致。闫小红等(2016)在利用光合电子流对光响应模型研究大狼把草、山莴苣和酸模叶蓼的光合特性差异时发现,3种植物电子传递速率对光强的响应曲线呈现先快速上升,饱和光强下达峰值,之后缓慢下降,出现光抑制现象,也能拟合得到植物的ETRmax和PARsat。Quiroz等(2017)关于马铃薯作物生长模型的研究也发现,光合电子流对光响应机理模型在马铃薯中使用效果很好,拟合得到的ETRmax和PARsat与实测值接近。康华靖等(2018)在对温州地区主栽蓝莓品种叶片饱和光强的研究结果表明,随光强的升高,叶片的电子传递速率呈线性增大,至饱和光强时达最大值,光强继续增加,电子传递速率则呈明显下降趋势,由模型得到的光饱和点值与实际观测值接近。
植物对光的捕获和利用能力可通过与光合作用相关的一些参数来反映。本研究中,超级晚稻品种的ETRmax和PARsat均显著高于对照仙优46,说明超级晚稻具有更强的光合吸收和利用能力。有研究表明,植物的电子传递速率与RuBP(1,5-二磷酸核酮糖)的再生能力密切相关,较低的电子传递速率会限制植物体内NADPH的生成与光合磷酸化,进而限制植物的最大羧化效率,最终降低其光合能力(Harley et al.,1992)。王强等(2014)对濒危植物长叶榧进行光合生理生态研究,结果发现夏季长叶榧较强的光合作用能力与其具有较高的ETRmax相关。闫小红等(2016)对不同草本植物光合特性差异进行研究,发现大狼把草的PARsat显著高于山莴苣,且具有更高的ETRmax,光适应能力更强。本研究中,超级晚稻的叶绿素含量均低于对照汕优46,但超级晚稻的ETRmax和PARsat均高于对照汕优46,可能与超级晚稻的σik较大、捕光色素分子的τmin较短有关。σik能反应植物捕光色素分子的潜在光能吸收能力,σik越大,捕光色素分子吸收光子的能力越强,越有利于植物光能的吸收(胡文海等,2017)。τmin越小,激子越容易退激发到稳定的基态,越有利于能量传递到光化学反应中心,电子传递速率就越大(闫小红等,2016)。因此,单纯认为叶绿素含量越高,ETRmax和PARsat就越大,越有利于植物光能的吸收并不全面,还必须考虑捕光色素分子的σ ik和τmin等内禀参数的影响。
植物的光合能力与原初反应过程中光合电子流的形成密不可分(叶子飘等,2014)。原初反应过程中捕光色素分子的内禀特性与植物的光合特性也密切相关(Brading et al.,2011)。本研究中,超级晚稻在不同光照强度下的有效光能截面均大于对照汕优46,表明超级晚稻在不同环境下具有更强的光能吸收利用能力。超级晚稻的这种光合特性表现与前人对百香果(Shimada et al.,2017)、草莓(Gao et al.,2017)光合特性的研究结果一致。捕光色素分子吸收光能后,将从基态跃迁到激发态。处于高能态的捕光色素分子将通过共振传递的方式将大部分能量传递到光反应中心,使PSⅡ发生电离,产生光合电子流,剩余的能量则通过荧光退激发和热耗散而消耗掉(Baker,2008;Takahashi and Badger,2011;Rey-nolds et al.,2012),如果过多的捕光色素分子处于激发态且不能及时回到基态,将对植物产生伤害(Takahashi and Badger,2011;Niyogi and Truong,2013)。本研究结果显示,在不同光照强度下,各超级晚稻品种的Nk均低于对照汕优46,表明超级晚稻品种中有较多的捕光色素分子处于基态。这可能与超级晚稻的τmin比对照汕优46小有关,τmin越小,处于基态的捕光色素分子越多,更有利于光能的吸收和利用。本研究仅从光合电子流的角度对超级稻的光合特性进行分析,今后还应从净光合速率对光响应、光能利用效率和光化学量子效率等角度进行全面分析。
4 结论
超级晚稻相比对照汕优46具有较好的光合特性,表现出最大电子传递速率大,饱和光强高的特性。水稻对光的捕获和利用能力是其光合作用的一个重要决定性因素,水稻捕光色素分子的内禀特性与光合特性密切相關。此外,捕光色素分子的有效光能吸收截面和处于激发态的捕光色素分子对光响应曲线的差异也有可能影响其光合特性。
参考文献:
蔡威威,艾天成,李然,金紫缘,徐金刚,曹坤坤. 2018. 控释肥及尿素添加剂对双季稻光合特性及产量的影响[J]. 中国土壤与肥料,(3):54-60. [Cai W W,Ai T C,Li R,Jin Z Y,Xu J G,Cao K K. 2018. Effect of controlled release fertilizer and urea additive on photosynthetic characteristics and yield of double cropping rice[J]. Soil and Fertillizer Sciences in China,(3):54-60.]
董桃杏,蔡昆争,曾任森. 2009. 茉莉酸甲酯(MeJA)对干旱胁迫下水稻幼苗光合作用特性的影响[J]. 生态环境学报,18(5):1872-1876. [Dong T X,Cai K Z,Zeng R S. 2009. Effects of methyl jasmonate(MeJA)on photosynthetic traits of rice seedlings under drought stress[J]. Ecology and Environmental Sciences,18(5):1872-1876.]
胡文海,叶子飘,闫小红,杨旭升. 2017. 越冬期广玉兰阳生叶和阴生叶PSⅡ功能及捕光色素分子内禀特性的比较研究[J]. 植物研究,37(2):281-287. [Hu W H,Ye Z P,Yan X H,Yang X S. 2017. PSⅡ function and intrinsic characteristics of light-harvesting pigment molecules for sun-and shading-leaf in Magnolia grandiflora during overwintering[J]. Bulletin of Botanical Research,37(2):281-287.]
康華靖,余宏傲,叶朝军,曾光辉,谢拾冰,王法格. 2018. 温州地区主栽蓝莓品种叶片饱和光强的研究[J]. 热带亚热带植物学报,26(4):399-406. [Kang H J,Yu H A,Ye C J,Zeng G H,Xie S B,Wang F G. 2018. Studies on light saturation point of main blueberry cultivars in Wenzhou[J]. Journal of Tropical and Subtropical Botany,26(4):399-406.]
林诚,李清华,王飞,何春梅,钟少杰,李昱,林新坚. 2016. 不同施磷水平对冷浸田水稻磷含量、光合特性及产量的影响[J]. 热带亚热带植物学报,24(5):553-558. [Lin C,Li Q H,Wang F,He C M,Zhong S J,Li Y,Lin X J. 2016. Effects of phosphorus fertilizer on phosphorus content,photosynthesis characters and yield of rice in cold waterlogged paddy field[J]. Journal of Tropical and Subtropical Botany,24(5):553-558.]
商天其,孙志鸿. 2019. 香樟幼龄林不同叶龄叶片的光合特征和单萜释放规律[J]. 应用与环境生物学报,25(1):89-99. [Shang T Q,Sun Z H. 2019. Photosynthetic characteristics and monoterpenes emission are related to foliage ontogeny in Cinnamomum camphora[J]. Chinese Journal of Applied and Environmental Biology,25(1):89-99.]
沈允钢,施教耐,许大全. 1998. 光合作用动力学[M]. 北京:科学出版社. [Shen Y G,Shi J N,Xu D Q. 1998. Kine-tics of photosynthesis[M]. Beijing:Science Press.]
宋微,高扬,步金宝. 2017. 不同穗型水稻品种的光合特性和光能利用率[J]. 黑龙江农业科学,(1):24-27. [Song W,Gao Y,Bu J B. 2017. Photosynthetic characteristics and sunlight efficiency of rice varieties with different panicle types[J]. Heilongjiang Agricultural Sciences,(1):24-27.]
王强,金则新,郭水良,管铭,王兴龙. 2014. 濒危植物长叶榧的光合生理生态特性[J]. 生态学报,34(22):6460-6470. [Wang Q,Jin Z X,Guo S L,Guan M,Wang X L. 2014. Photosynthetic traits of the endangered plant species Torreya jackii[J]. Acta Ecologica Sinica,34(22):6460-6470.]
吴俊,邓启云,袁定阳,齐绍武. 2016. 超级杂交稻研究进展[J]. 科学通报,61(35):3787-3796. [Wu J,Deng Q Y,Yuan D Y,Qi S W. 2016. Progress of super hybrid rice research in China[J]. Chinese Science Bulltin,61(35):3787-3796.]
闫小红,周兵,王宁,叶子飘,尹增芳. 2016. 基于光响应机理模型的3种草本植物光合特性差异解析[J]. 南京林业大学学报(自然科学版),40(3):63-69. [Yan X H,Zhou B,Wang N,Ye Z P,Yin Z F. 2016. Differences of photosynthetic characteristics based on a mechanistic model of light-response for three herbaceous plants[J]. Journal of Nanjing Forestry University(Natural Sciences Edition),40(3):63-69.]
杨旺兴,卓伟,马彬林,邹文广,韦新宇,杨滕帮,张受刚,许旭明. 2017. 水稻种质资源光合与水分利用特性比较及聚类分析[J]. 福建农业学报,32(3):248-252. [Yang W X,Zhuo W,Ma B L,Zou W G,Wei X Y,Yang T B,Zhang S G,Xu X M. 2017. Comparison and cluster analysis on photosynthetic characteristics and water usage efficiency of rice germplasms[J]. Fujian Journal of Agricultural Scien-ces,32(3):248-252.]
叶子飘,段世华,安婷,康华靖. 2018a. 最大电子传递速率的确定及其对电子流分配的影响[J]. 植物生态学报,42(4):498-507. [Ye Z P,Duan S H,An T,Kang H J. 2018a. Determination of maximum electron transport rate and its impact on allocation of electron flow[J]. Chinese Journal of Plant Ecology,42(4):498-507.]
葉子飘,胡文海,肖宜安. 2014. 光合电子流对光响应的机理模型及其应用[J]. 植物生态学报,38(11):1241-1249. [Ye Z P,Hu W H,Xiao Y A. 2014. A mechanistic model of light-response of photosynthetic electron flow and its application[J]. Chinese Journal of Plant Ecology,38(11):1241-1249.]
叶子飘,胡文海,闫小红,段世华. 2016. 基于光响应机理模型的不同植物光合特性[J]. 生态学杂志,35(9):2544-2552. [Ye Z P,Hu W H,Yan X H,Duan S H. 2016. Photosynthetic characteristics of different plant species based on a mechanistic model of light-response of photosynthesis[J]. Chinese Journal of Ecology,35(9):2544-2552.]
叶子飘,杨小龙,冯关萍. 2018b. 植物电子传递速率对光响应模型的比较研究[J]. 扬州大学学报(农业与生命科学版),39(1):97-104. [Ye Z P,Yang X L,Feng G P. 2018b. Comparative study on light-response models of electron transport rates of plants[J]. Journal of Yangzhou University(Agricultural and Life Science Edition),39(1):97-104.]
赵森,于江辉,肖国樱. 2013. 高温胁迫对爪哇稻剑叶光合特性和渗透调节物质的影响[J]. 生态环境学报,22(1):110-115. [Zhao S,Yu J H,Xiao G Y. 2013. Effects of high temperature stress on the photosynthesis and osmoregulation substances of flag leaves in Oryza stavia L. ssp. Javanica[J]. Ecology and Environmental Sciences,22(1):110-115.]
周根友,翟彩娇,邓先亮,张蛟,张振良,戴其根,崔士友. 2018. 盐逆境对水稻产量、光合特性及品质的影响[J]. 中国水稻科学,32(2):146-154. [Zhou G Y,Zhai C J,Deng X L,Zhang J,Zhang Z L,Dai Q G,Cui S Y. 2018. Performance of yield,photosynthesis and grain quality of japonica rice cultivars under salinity stress in micro-plots[J]. Chinese Journal of Rice Science,32(2):146-154.]
周祖富,黎兆安. 2005. 植物生理学试验指导[M]. 北京:中国农业出版社:114-116. [Zhou Z F,Li Z A. 2005. A gui-dance on plant physiological experiment[M]. Beijing:China Agricultural Press:114-116.
Archontoulis S V,Yin X,Vos J,Danalatos N G,Struik P C. 2012. Leaf photosynthesis and respiration of three bioe-nergy crops in relation to temperature and leaf nitrogen:How conserved are biochemical model parameters among crop species?[J]. Journal of Experimental Botany,63(2):895-911.
Baker N R. 2008. Chlorophyll fluorescence:A probe of photosynthesis in vivo[J]. Annual Review of Plant Biology,59:89-113.
Brading P,Warner M E,Davey P,Smith D J,Achterberg E P,Suggett D J. 2011. Differential effects of ocean acidification on growth and photosynthesis among phylotypes of Symbiodinium(Dinophyceae)[J]. Limnology and Ocea-nography,56(3):927-938.
Gao S,Huan L,Lu X P,Jin W H,Wang X L,Wu M J,Wang G C. 2016. Photosynthetic responses of the low intertidal macroalga Sargassum fusiforme(Sargassaceae)to saline stress[J]. Photosynthetica,54(3):430-437.
Gao S,Yan Q,Chen L X,Song Y B,Li J M,Fu C X,Dong M. 2017. Effects of ploidy level and haplotype on variation of photosynthetic traits:Novel evidence from two Fragaria species[J]. PLoS One,12(6):e0179899.
Govindjee. 2002. A role for a light-harvesting antenna complex of photosystem II in photoprotection[J]. The Plant Cell,14:1663-1668.
Harley P C,Thomas R B,Reynolds J F,Strain B R. 1992. Modelling photosynthesis of cotton grown in elevated CO2[J]. Plant Cell & Environment,15(3):271-282.
Kathuria H,Giri J,Tyagi H,Tyagi A K. 2007. Advances in transgenic rice biotechnology[J]. Critical Reviews in Plant Sciences,26 (2):65-103.
Krall J P,Edward G E. 1992. Relationship between photosystem II activity and CO2 fixation in leaves[J]. Physiologia Plantarum,86(1):180-187.
Li X N,Hao C L,Zhong J W,Liu F L,Cai J,Wang X,Zhou Q,Dai T B,Cao W X,Jiang D. 2015. Mechano-stimula-ted modifications in the chloroplast antioxidant system and proteome changes are associated with cold response in wheat[J]. BMC Plant Biology,15:219.
Major K M,Dunton K H. 2002. Variations in light-harvesting characteristics of the seagrass,Thalassia testudinum:Evidence for photoacclimation[J]. Journalof Experimental Marine Biology and Ecology,275(2):173-189.
Nelson N,Yocum C F. 2006. Structure and function of photosystems I and II[J]. Annual Review of Plant Biology,57:521-565.
Niyogi K K,Truong T B. 2013. Evolution of flexible non-photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis[J]. Current Opinion in Plant Biology,16(3):307-314.
Panitchayangkoon G,Hayes D,Fransted K A,Caram J R,Harel E,Wen J Z,Blankenship R E,Engel G S. 2010. Long-lived quantum coherence in photosynthetic complexes at physiological temperature[J]. Proceedings of the National Academy of Sciences,107(29):12766-12770.
Quiroz R,Loayza H,Barreda C,Gavilan C,Posadas A,Ramirez D A. 2017. Linking process-based potato models with light reflectance data:Does model complexity enhance yield prediction accuracy?[J]. European Journal of Agro-nomy,82:104-112.
Reynolds Matthew,Foulkes J,Furbank R,Griffiths S,King J,Murchie E,Parry M,Slafer G. 2012. Achieving yield gains in wheat[J]. Plant,Cell & Environment,35(10):1799-1823.
Shimada A,Kubo T,Tominaga S,Yamamoto M. 2017. Effect of temperature on photosynthesis characteristics in the passion fruits ‘Summer Queen’ and ‘Ruby Star’[J]. The Horticulture Journal,86(2):194-199.
Suggett D J,MacIntyre H L,Geider R J. 2004. Evaluation of biophysical and optical determinations of light absorption by photosystem II in phytoplankton[J]. Limnology and Oceanography Methods,2:316-332.
Takahashi S,Badger M R. 2011. Photoprotection in plants:A new light on photosystem II damage[J]. Trends in Plant Science,16(1):53-60.
Wang X X,Wang W C,Huang J L,Peng S B,Xiong D L. 2017. Diffusional conductance to CO2 is the key limitation to photosynthesis in salt-stressed leaves of rice(Oryza sativa)[J]. Physiologia Plant,163(1):45-58.
White A J,Critchley C. 1999. Rapid light curves:A new fluorescence method to asses the state of the photosynthetic apparatus[J]. Photosynthesis Research,59(1):63-72.
Xu J Z,Yu Y M,Peng S Z,Yang S H,Liao L X. 2014. A modified nonrectangular hyperbola equation for photosynthetic light-response curves of leaves with different nitrogen status[J]. Photosynthetica,52(1):117-123.
Ye Z P,Robakowski P,Suggett D J. 2013a. A mechanistic model for the light response of photosynthetic electron transport rate based on light harvesting properties of photosynthetic pigment molecules[J]. Planta,237:837-847.
Ye Z P,Suggett D J,Robakowski P,Kang H J. 2013b. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species[J]. New Phytologist,199(1):110-120.
(責任编辑 王 晖)
猜你喜欢
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
湖北农业科学(2015年16期)2015-10-28
长江蔬菜·学术版(2015年1期)2015-07-10
长江蔬菜·学术版(2015年6期)2015-06-24
天津农业科学(2015年6期)2015-06-16
