
His-pull down联合质谱鉴定谷氨酸棒杆菌中乙酰羟酸合酶IlvN的相互作用蛋白
2019-09-10 06:40侯正杰张权威莫晓琳夏利谭淼孙全伟马倩陈宁
食品与发酵工业 2019年16期
侯正杰,张权威,莫晓琳,夏利,谭淼,孙全伟,马倩,2,3*,陈宁,2,3*
1(天津科技大学 生物工程学院,天津,300457) 2(代谢控制发酵技术国家地方联合工程实验室,天津,300457)3(天津市微生物代谢与发酵过程控制技术工程中心,天津,300457)
支链氨基酸包括L-缬氨酸(L-valine)、L-亮氨酸(L-leucine)和L-异亮氨酸(L-isoleucine),其疏水脂质链都具有分支的甲基基团,所以被称为分支链氨基酸[1]。支链氨基酸作为人体必需氨基酸,不仅是蛋白质的合成原料,而且还具有特殊的生理、生物学功能,也可以作为生物体能源[2]。目前,支链氨基酸被广泛应用于氨基酸输液、营养强化剂等医药行业[3]和洗涤剂[4]、除草剂领域[5-7],需求量逐年增高。目前生产支链氨基酸的方法主要为微生物发酵法[8]。支链氨基酸生产菌种主要为谷氨酸棒杆菌(Corynebacteriumglutamicum)(包括其亚种黄色短杆菌Brevibacteriumflavum和乳糖发酵短杆菌Brevibacteriumlactofermentum等)[9-10]和大肠杆菌(Escherichiacoli)[11]。目前,大多数生产企业所采用的菌株主要是通过常规诱变育种获得[12],遗传背景不清晰,导致产量很难再进一步提高[13]。
谷氨酸棒杆菌中分支链氨基酸的合成主要涉及中心碳代谢与分支合成途径。中心碳代谢中积累的丙酮酸是分支合成途径的关键底物。分支合成途径从丙酮酸出发[14],经过一系列相同或相关酶的催化,最终形成不同的分支链氨基酸产品。其中,缬氨酸与异亮氨酸合成的最后4步反应为共同的酶催化,分别以丙酮酸、α-酮丁酸为出发底物[15],依次经乙酰羟酸合酶(acetohydroxy acidsynthase,由ilvBN编码)、乙酰羟酸同分异构体还原酶(acetohydroxy acid isomeroreductase,由ilvC编码)、二羟酸脱水酶(dihydroxy acid dehydratase,由ilvD编码)、转氨酶(transaminase,由ilvE编码)催化而成。其中,乙酰羟酸合酶(IlvBN)作为3种分支链氨基酸分支合成途径共同的第一步反应催化酶,是分支合成途径的限速酶,其与丙酮酸、α-酮丁酸的亲和力强弱,直接决定了缬氨酸或异亮氨酸的合成方向与合成效率。
谷氨酸棒杆菌乙酰羟酸合酶由催化亚基和调节亚基组成,分别由ilvB和ilvN编码[16]。其中,大亚基(IlvB)负责催化功能,小亚基(IlvN)负责调控该酶受产物分支链氨基酸的反馈抑制[17]。研究表明,通过调节乙酰羟酸合酶的活性,可以获得不同分支链氨基酸生产菌株[18-19]。IlvBN受终产物反馈抑制的解除对于分支链氨基酸合成效率的提升具有关键的作用,是代谢工程中重要的改造位点。从STRING(functional protein association networks,http://string-db.org/)数据库中可提取到CorynebacteriumglutamicumATCC 13032中与IlvBN存在密切相关性的多酶群模型,由此可见,IlvBN与其他蛋白质之间存在密切的相互作用。特别是与IlvN存在相互作用的蛋白,对于反馈抑制的调节,会产生不同的影响。
本研究以本实验室诱变筛选得到的缬氨酸高产菌CorynebacteriumglutamicumXV为研究对象,利用his-pull down技术,筛查与IlvN具有相互作用的蛋白,这些蛋白的后续调控将成为分支链氨基酸生产菌株代谢工程育种的潜在改造策略。本研究以关键酶乙酰羟酸合酶调节亚基IlvN的相互作用蛋白研究为切入点来探究关键酶之间的相互作用模式,为谷氨酸棒杆菌的代谢工程改造以及强化氨基酸产品工业化生产提供重要信息。
1 材料与方法
1.1 材料与试剂
E.coliDH5α、E.coliRosetta、质粒pET-28a:本实验室保藏;限制性内切酶EcoR I、Hind Ⅲ、T4 DNA连接酶:日本Takara公司;高效Ni-NTA:GenScript公司;过硫酸铵、丙烯酰胺、N,N′-亚甲基双丙烯酰胺:SbaseBio公司;TEMED:Sigma公司;蛋白脱盐柱:美国GE公司;BCA蛋白浓度试剂盒:PPLYGEN公司;胰蛋白酶:Promega公司。
1.2 仪器与设备
JY96-ⅡN超声波细胞破碎仪,宁波新芝;AKTA avant25蛋白纯化系统,GE Healthcare公司;DYCZ-25E型P4垂直电泳仪,北京六一仪器厂;SpectraMax M4多功能酶标仪,Molecular Devices公司;EPORATOR电转仪,eppendorf公司;CRZ1 GⅢ高速冷冻离心机,日本日立公司;EASY-nLCTM 1200纳升级UHPLC,Thermo公司;Q ExactiveTMHF-X质谱仪,Thermo公司。
1.3 方法
1.3.1ilvN基因的克隆以及原核表达载体构建
以C.glutamicumXV基因组为模板,上游引物:5′CCGGAATTCATGGCTAATTCTGACGTCACCC3′(含EcoR I酶切位点);下游引物:5′CCCAAGCTTTTAGATCTTGGCCGGAGCC 3′(含Hind Ⅲ酶切位点),PCR扩增出对应序列DNA长度(519 bp)。目的产物用Takara限制性内切酶EcoR I和Hind Ⅲ酶切后,用T4 DNA连接酶将其与经相同内切酶切割的表达载体pET-28a连接,连接产物转化至感受态大肠杆菌E.coliDH5α感受态细胞中,在含卡那霉素(100 μg/mL)琼脂平板上培养挑选单克隆,经菌落PCR验证后挑取阳性菌株,保菌。以碱裂解法小提重组质粒,用Takara限制性内切酶EcoR I和Hind Ⅲ进行双酶切鉴定和测序。
1.3.2 构建蛋白表达菌株
将携带ilvN基因的重组质粒通过电转化法转化到E.coliRosetta菌株中,使用电压为1 850 V。在含卡那霉素(100 μg/mL)和氯霉素(50 μg/mL)的琼脂平板上挑选单克隆,经菌落PCR验证后挑取阳性菌株,保菌。
1.3.3 IlvN蛋白诱导表达
从保菌管中取适量菌液接种于装有5 mL含卡那霉素(100 μg/mL)和氯霉素(50 μg/mL)LB培养基的摇管中进行活化,37 ℃,200 r/min培养12 h;从培养好的摇管中以体积分数为1%的接种量接种于装有100 mL LB培养基的种子瓶中,双抗,37 ℃,200 r/min培养12 h;从培养好的种子瓶以1%接种量接种于装有400 mL LB培养基的发酵瓶中,双抗,37 ℃,200 r/min培养至OD600为0.6左右时加入0.1 mmol/L的异丙基-β-D-硫代半乳糖苷(isopropyl-β-D-thiogalact opyranosied,IPTG)诱导剂,18 ℃,180 r/min继续诱导培养10 h。之后8 000 r/min,4 ℃离心15 min收集菌体,收集到的菌体用PBS缓冲液清洗3遍。清洗好的菌体用适量平衡缓冲液重悬起来,加入10 mmol/L PMSF,冰水浴超声破碎20 min,破碎完成后,13 000 r/min、4 ℃离心15 min取上清,沉淀用离心前原体积的平衡缓冲液重悬起来,取上清样和沉淀样处理后进行SDS-PAGE检测蛋白表达情况。
1.3.4 IlvN蛋白纯化
吸取2 mL的Ni+树脂(体积分数为50%的树脂匀浆)于柱内,用平衡缓冲液冲洗平衡好。将收集到的破碎液上清与平衡好的Ni+树脂混合,使用小型磁力搅拌器,4℃吸附1 h。吸附后上柱,用含不同咪唑浓度的梯度缓冲液洗脱后分别取样进行SDS-PAGE检测,确定最佳洗涤缓冲液和洗脱缓冲液中咪唑浓度。用10倍柱体积的洗涤缓冲液洗脱杂蛋白,10倍柱体积的洗脱缓冲液洗脱目的蛋白,分别取样后进行SDS-PAGE检测。收集洗脱蛋白,用GE蛋白脱盐柱脱盐,酶标仪BCA法测定蛋白浓度。
1.3.5 提取C.glutamicumXV胞内蛋白
取1.2 L BHI培养基,接种量φ(菌液)=1%,32 ℃,200 r/min培养12 h,离心收集菌体。菌体用PBS缓冲液洗涤后,液氮快速研磨3次,之后用冰冷的平衡缓冲液重悬起来,加入10 mmol/L PMSF,冰水浴超声破碎1 h,13 000 r/min,4 ℃离心15 min取上清,透过0.45 μm的滤膜去除杂质。酶标仪BCA法测定胞内蛋白浓度。
1.3.6 His-pull down实验
纯化脱盐后的IlvN蛋白与2 mL Ni+树脂混合,使用小型磁力搅拌器,4 ℃吸附1 h;之后加入提取的C.glutamicumXV胞内蛋白于混合体系中,使用垂直混悬仪4 ℃混合孵育过夜。孵育完成后进行纯化。未和胞内蛋白混育的一组作空白对照,其他条件均相同。取样后进行SDS-PAGE检测,对比差异性条带。
1.3.7 蛋白酶解
用刀片切取特异性条带上的蛋白胶粒,加入蛋白溶解液补足体积至100 μL,加入2 μL 1 g/L胰酶和500 μL 100 mmol/L TEAB缓冲液,混匀后于37℃酶解过夜。加入等体积的体积分数为1%甲酸,混匀后于室温、12 000 r/min离心5 min,取上清缓慢通过C18除盐柱,之后使用1 mL清洗液(体积分数为0.1% 甲酸、4%乙腈)连续清洗3次,再加入0.4 mL洗脱液(体积分数为0.1%甲酸、45%乙腈)连续洗脱2次,洗脱样合并后冻干。
1.3.8 液质检测与蛋白鉴定
蛋白酶解后直接上样,使用EASY-nLCTM1200纳升级UHPLC系统进行液质检测,所得到的数据用Proteome Discoverer 2.2软件进行数据库检索,进行蛋白定性。
色谱条件:预柱为Acclaim PepMap100 C18Nano-Trap(2 cm×100 μm, 5 μm),分析柱为Reprosil-Pur 120 C18-AQ(15 cm×150 μm,1.9 μm),流动相A(体积分数为100%水、0.1%甲酸)和B(体积分数为80%乙腈、0.1%甲酸),液相色谱洗脱条件如表1所示。
质谱条件:EASY-SprayTM离子源,设定离子喷雾电压为2.3 kV,离子传输管温度为320 ℃,质谱采用数据依赖型采集模式,质谱全扫描范围为m/z350~1 500,一级质谱分辨率设为60 000(200m/z),C-trap最大容量为3×106,C-trap最大注入时间为20 ms;选取全扫描中离子强度TOP 40的母离子使用高能碰撞裂解(higher energy collison induced dissociation,HCD)方法碎裂,进行二级质谱检测,二级质谱分辨率设为15 000(200m/z),C-trap最大容量为1×105,C-trap最大注入时间为45 ms,肽段碎裂碰撞能量设为27%,阈强度设为8.3×103,动态排阻范围设为60 s。Proteome Discoverer 2.2分析参数如表2所示。
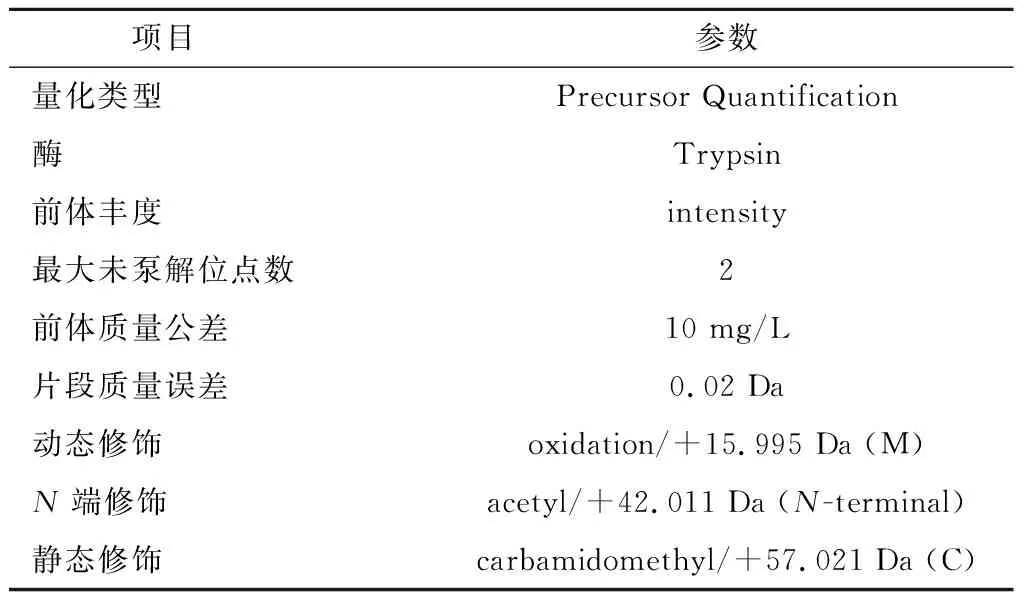
表2 Proteome Discoverer 2.2分析参数Table 2 Analysis parameters of Proteome Discoverer 2.2
2 结果与分析
2.1 重组质粒的酶切鉴定结果
重组质粒经EcoR I和Hind Ⅲ双酶切后,产生了2条片段,分别为目的片段和线性化载体。结合测序结果,进一步证实ilvN克隆片段的插入方向和编码区序列正确,与Genbank中载入的序列完全一致。
2.2 IlvN蛋白的表达、纯化
以携带不含ilvN基因的pET-28a空质粒的Rosetta菌株作为空白对照,为减少包涵体的形成,对IlvN蛋白进行低温诱导表达(图1),泳道4为细胞破碎液上清样,可以看出IlvN蛋白仍有一部分包涵体形成,但也有明显的可溶性表达。后续通过调整诱导表达条件,进一步提高了IlvN的可溶性表达。之后对可溶性IlvN蛋白用镍离子树脂进行纯化,咪唑梯度洗脱,洗脱浓度分别为10、50、75、100、150、200、300、500 mmol/L(图2)。由电泳结果可以看出用含100 mmol/L咪唑的缓冲液可以洗脱掉杂蛋白,300 mmol/L咪唑的缓冲液洗脱目的蛋白时效果较好,可以得到比较纯的蛋白。以最优条件纯化后进行SDS-PAGE电泳(图3),由BandScan软件分析纯化脱盐后的蛋白纯度达到了98%,可以进行后续实验。利用多功能酶标仪,BCA法测得脱盐后蛋白质量浓度为0.56 g/L。
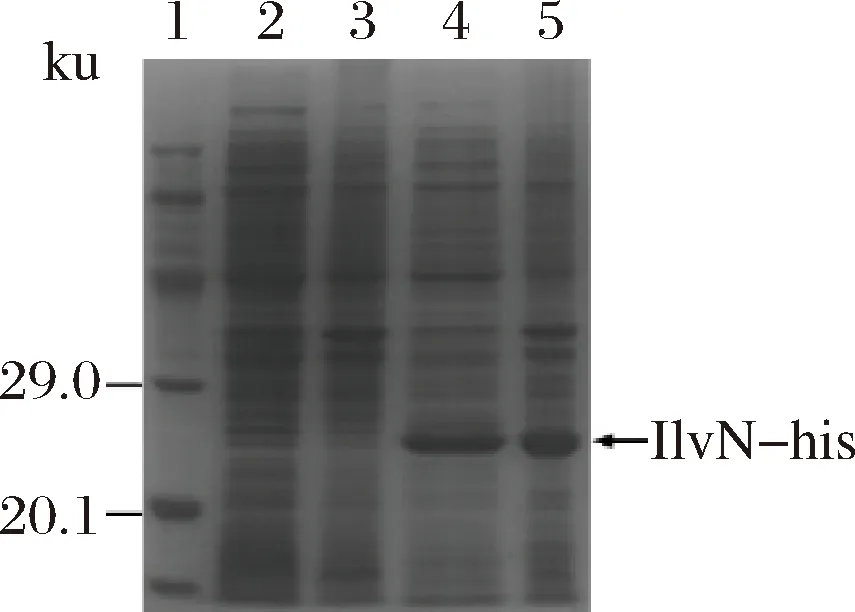
1-蛋白分子质量标准;2-空白对照上清;3-空白对照沉淀;4- IlvN-his上清;5-IlvN-his沉淀图1 Ilvn蛋白表达SDS-PAGE结果Fig.1 SDS-PAGE result of Ilvn protein expression
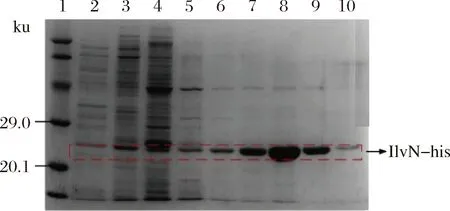
1-蛋白分子质量标准; 2-破碎液自然流穿样;3-10 mmol/L咪唑洗涤;4~10-50、75、100、150、200、300、500 mmol/L咪唑梯度洗脱图2 Ilvn蛋白咪唑梯度洗脱SDS-PAGE结果Fig.2 SDS-PAGE result of Ilvn protein imidazole gradient elution
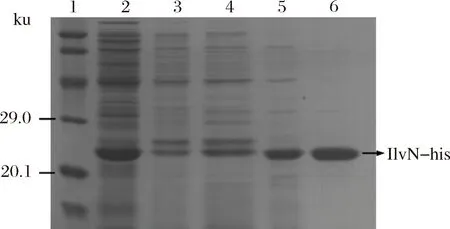
1-蛋白分子质量标准;2-自然过柱样;3~5-30、50、100 mmol/L咪唑洗涤;6-300 mmol/L咪唑洗脱图3 Ilvn蛋白纯化SDS-PAGE结果Fig.3 SDS-PAGE result of Ilvn protein purification
2.3 His-pull down进行IlvN相互作用蛋白的筛选
用BCA法测得提取的C.glutamicumXV胞内蛋白质量浓度为5.19 g/L。取1 mg纯化脱盐的IlvN-his蛋白和80 mg胞内蛋白混育过夜,获得his-IlvN-胞内蛋白相互作用复合体,以80 mg胞内蛋白作为空白对照。经过前期实验探索,选用50 mmol/L的咪唑洗涤样品,然后用300 mmol/L的咪唑洗脱液,将捕获到的含有his标签的蛋白洗脱下来,并进行SDS-PAGE,结果如图4所示。通过比较泳道4与5,发现5泳道中多出1条比较明显的条带,如图4所示。此条带中所含蛋白则很可能与IlvN具有相互作用,因此一起被洗脱(pull down)下来。
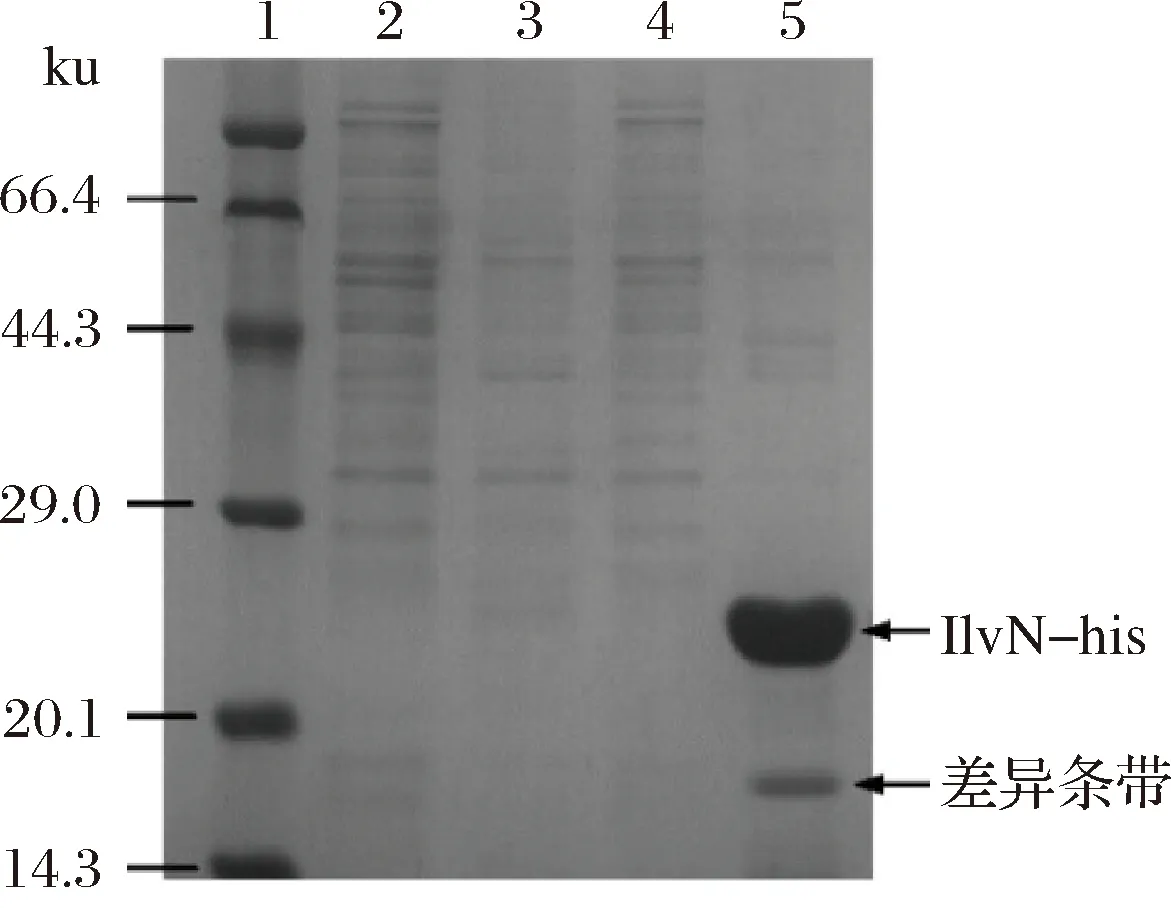
1-蛋白分子质量标准;2-空白对照50 mmol/L咪唑洗涤;3-共孵育后50 mmol/L咪唑洗涤;4-空白对照300 mmol/L咪唑洗脱;5-共孵育后300 mmol/L咪唑洗脱图4 Pull down实验SDS-PAGE结果Fig.4 SDS-PAGE result of pull down experiment
2.4 IlvN相互作用蛋白的鉴定
对上述获得的差异蛋白条带处理后通过液质联用进行鉴定。为了提高分析结果质量,降低假阳性率,Proteome Discoverer 2.2软件对检索结果做了进一步过滤:可信度在95%以上的谱肽(peptide spectrum matches,PSMs)为可信PSMs,至少包含一个特有肽段的蛋白为可信蛋白,只保留可信的谱肽和蛋白,并做FDR验证,去除FDR>5%的肽段和蛋白。
分析后共鉴定得到45种蛋白,具体信息如表3所示。通过利用COG数据库对鉴定到的蛋白质进行功能注释,以了解不同蛋白质的功能特性。由COG注释结果看(图5),大部分蛋白为参与翻译过程的核糖体蛋白,此外,还包括参与能量产生与转换、氨基酸转运与代谢、碳水化合物转运与代谢、脂转运与代谢、信号传导等过程的蛋白质。
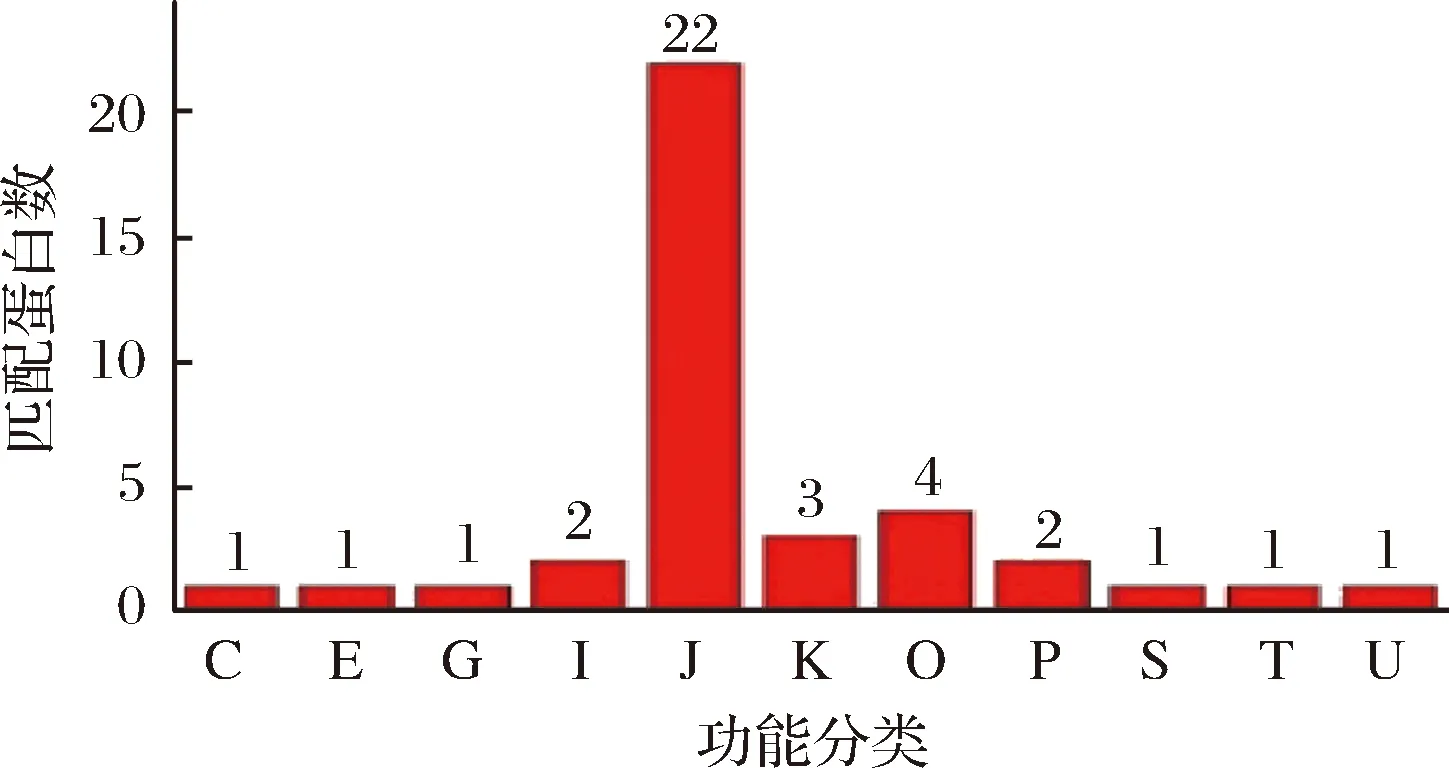
C-能量产生与转化(1);E-氨基酸转运与代谢(1);G-碳水化合物转运与代谢(1); I-脂质转运与代谢(2) ;J-翻译、核糖体结构和生物发生(22);K-转录(3);O-翻译后修饰蛋白转换,伴侣(4);P-无机离子转运与代谢(2);S-功能未知(1);T-信号传导机制(1);U-细胞运输、分泌和囊泡运输(1)图5 COG注释结果柱状图Fig.5 COG annotated results histogram
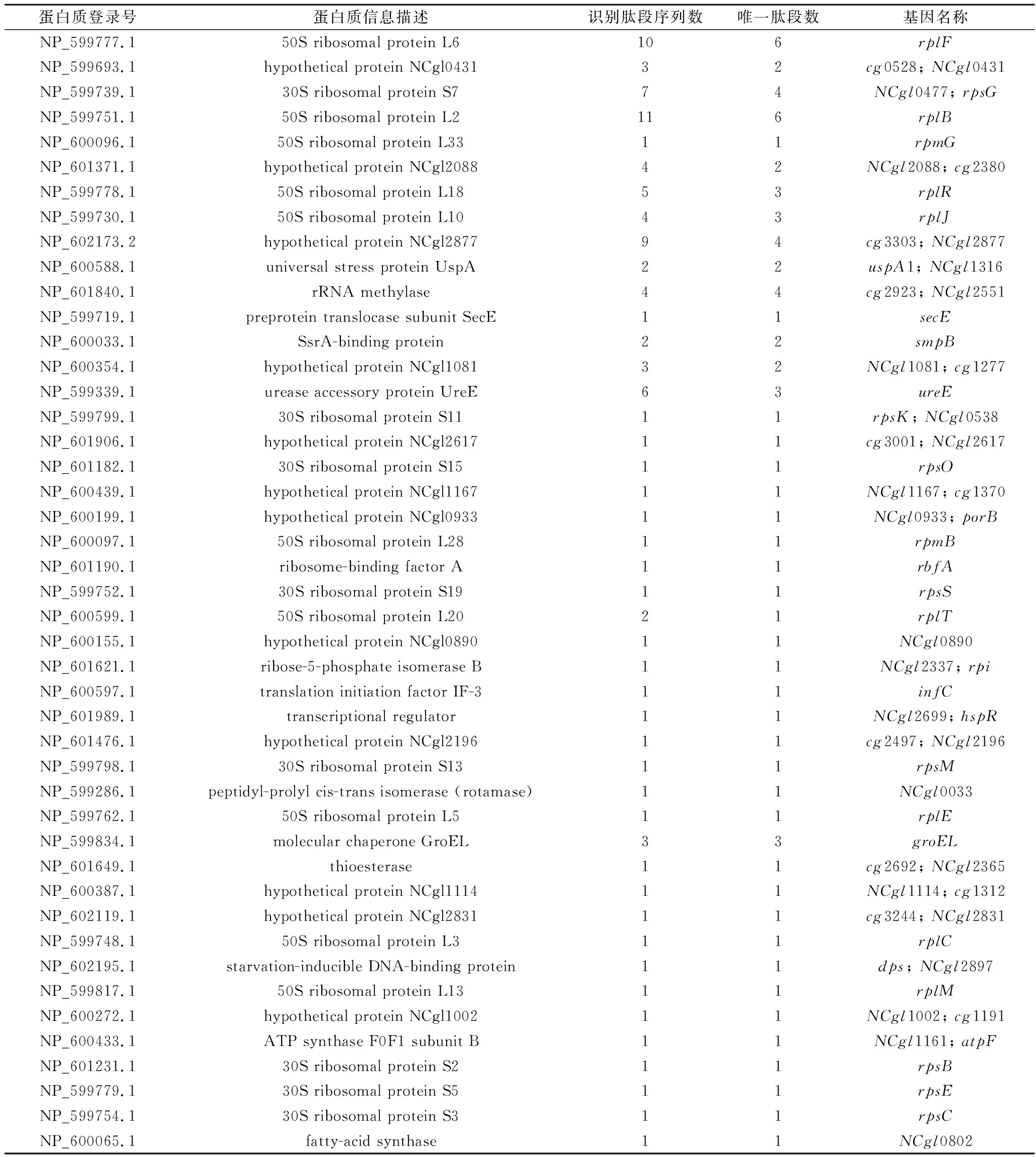
表3 IlvN相互作用蛋白鉴定结果Table 3 Identification of interactive proteins of IlvN
分支链氨基酸的合成过程涉及中心碳代谢过程与分支合成途径,其中,中心碳代谢过程为其提供了碳骨架,分支合成途径决定了具体分支链氨基酸种类的合成方向。在表3所示的相互作用蛋白列表中,Rpi (ribose-5-phosphate isomerase B),负责催化5-磷酸核糖与5-磷酸核酮糖之间的相互转化,因此,连接了磷酸戊糖途径的氧化阶段与非氧化阶段。由此可见磷酸戊糖途径与分支链氨基酸的合成具有密切的联系。此外,5-磷酸核糖是细胞内合成核糖的主要途径,因此,Rpi对于细胞内核酸合成的五碳糖原料供应也具有十分重要的作用。
脂肪酸合成过程对于细胞膜的结构与功能具有重要的作用,特别是在谷氨酸棒杆菌谷氨酸的分泌方面已有较多研究,众多研究结果表明,脂肪酸合成影响细胞膜的通透性,从而可以影响氨基酸等小分子进出细胞的过程。在谷氨酸的传统生产工艺中,采用限制生物素的供应,减弱脂肪酸的合成,使细胞通透性增强,可以明显提高谷氨酸的分泌速度。本课题组前期研究发现,细胞膜与细胞壁通透性的提高对于缬氨酸的分泌同样具有提升效果。本研究中筛查到的相互作用蛋白中,存在脂肪酸合成酶NCgl0802,进一步表明分支链氨基酸的合成与输出过程与脂肪酸的合成具有密切的联系。
UspA是一种应激蛋白,在细胞受到各种压力时,UspA的表达量会明显升高,增强细胞的存活能力。从本研究结果来看,UspA与IlvN之间也可能存在相互作用,提示分支链氨基酸的合成过程与细胞的应激性能也有一定联系,后续可再进一步展开深入研究。
3 展望
蛋白质与蛋白质之间相互作用构成了细胞生化反应网络的一个主要组成部分,蛋白之间的互作网络与转录调控网络对调控细胞及其信号有重要意义。近年来,揭示蛋白质之间的相互作用关系以及相互作用图谱的建立逐渐成为功能研究的一个主要焦点。现在已经有多种方法用于研究蛋白质的相互作用,其中基于体外相互作用的洗脱技术应用广泛,具有特异性强、操作简便、减少假阳性率等特点,结合液质联用技术可以很好地用于筛选蛋白质之间的相互作用,并且实现高通量[20]。我们可以借助这一方法来研究微生物代谢过程中关键酶的蛋白相互作用,为进一步代谢工程改造提供策略。
在前期的研究中,实验室从同一亲本出发,已经选育了3株不同分支链氨基酸的高产菌株,并且实现了工业化生产,菌株生产能力居世界前列。在此基础上,利用这3株来源于同一亲本的不同分支链氨基酸产品生产菌,以关键酶乙酰羟酸合酶调节亚基IlvN的相互作用蛋白研究为切入点,初步筛选出与其存在相互作用的宿主蛋白,也说明了这种方法的可行性。后期我们将结合蛋白组和转录组学的方法,逐渐完善其互作模式和互作网络。需要注意的是,洗脱的方法虽然可以有效地降低实验中存在的假阳性,但仍无法避免,与其中鉴定出来的某一特定蛋白的相互作用也需要通过进一步实验来证实。所得的结果对基于关键酶间的相互作用关系进行多酶协调代谢工程改造,对谷氨酸棒杆菌进行有效的人为代谢干预、获得理想的基因表型将提供有效指导,对强化谷氨酸棒杆菌分支链氨基酸的工业化生产也具有重要意义。
猜你喜欢
河北北方学院学报(自然科学版)(2022年11期)2022-02-03
中国科技纵横(2021年24期)2021-03-02
蚕桑通报(2020年1期)2020-07-10
农药科学与管理(2019年9期)2019-11-23
中成药(2018年6期)2018-07-11
中成药(2017年6期)2017-06-13
中国洗涤用品工业(2017年2期)2017-04-16
中国比较医学杂志(2017年5期)2017-01-17
中国卫生标准管理(2015年2期)2016-01-14
肿瘤影像学(2015年3期)2015-12-09
