
湖北海棠枝干溃疡病病原鉴定及生理学特性研究
2019-09-09 09:24张兆霞郭慧玲刘会香崔玉国申卫星王玉杰
山东林业科技 2019年4期
张兆霞,郭慧玲*,刘会香,崔玉国,申卫星,王玉杰,陈 岩
(1.泰山森林病虫害防治检疫站,山东 泰安271000;2.泰山风景名胜区管委会,山东 泰安 27100;3.山东农业大学,山东 泰安271018)
湖北海棠(Malus hupehensis(Pamp.)Rehd..)位于泰山南天门海拔1300m以上,在天街至碧霞祠一带成片分布,因为适应泰山极顶的地理气候环境,湖北海棠常被称作“泰山海棠”,是泰山高海拔地区特有的水土保持和景观树种。每年春季岱顶大片海棠绽开蓓蕾,连天开放,碧霞祠一带像是成了花的海洋,秋季则红果累累,挂满枝头。泰山现有海棠林大部分为上世纪60年代人工造林而成,由于林龄过熟和山顶特殊气候原因,普遍存在生长不良、树势衰弱,甚至出现树木干枯死亡情况。对湖北海棠林分进行有害生物种类调查的基础上,确定湖北海棠生长衰弱的原因主要是由于枝干溃疡病的病菌侵染扩散所致。本文野外采集湖北海棠枝干溃疡病的病菌病斑进行室内培养和分子鉴定,为下一步科学治理海棠溃疡病提供依据。
1 材料与方法
1.1 试验材料
从野外发生枝干溃疡病的湖北海棠树上剪取若干病枝条。
1.2 试验方法
1.2.1 病菌的分离方法
在实验室从海棠枝干病斑处切取大小约5mm病组织共15块,采用常规组织分离法。先移置于PDA培养基平板放在25℃恒温箱中培养2~3天[5]。统计各菌株出现的概率,纯化后4℃冰箱保存备用。
1.2.2 病菌培养
湖北海棠溃疡病菌分离纯化后在25℃培养5天后,观察其菌落特征,用显微镜观察病菌子实体大小、形态及孢子的形态。在菌丝体中钻取5个菌饼,将菌接种到PD(液体马铃薯)培养基中,放置于28℃,200rpm恒温摇床上培养两天,于-20℃保存。
1.2.3 病菌DNA的提取
DNA的提取采用2×CTAB法,将培养好的菌丝烘干加入液氮冷冻研磨至细小粉末,取0.25g粉末加入预热的2-ME/CTAB抽提液(1ml)中,65℃保温45~60min,颠倒混匀,加入等体积酚:氯仿:异戊醇(25:24:1)溶液混匀,12000rpm/min 离心 30min,取上清加入 0.5~0.6(350uL)倍体积冰冷的异丙醇,颠倒混匀,-20℃放置20min;12000rpm/min离心10min;去上清夜,DNA沉淀以70%乙醇洗涤并干燥后溶于50ug/mL RNase的TE缓冲液,置于-20℃保存备用。最后用10g/l琼脂糖凝胶检测提取DNA的质量。
1.2.4 rDNA及ITS序列扩增
利用真核生物的基因通用引物ITS1/ITS4进行ITS序列扩增,引物序列为 ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) 和 ITS4(5’ -TCCTCCGCTTATTGATATGC-3’)。PCR 总反应体系为 25 ul,其中10×PCR Buffer为2.5ul,Mg2+1ul,dNTP 1ul,ITS1和 ITS4各为 2uL,Taq酶 0.5uL,ddH2O14ul,DNA模板2uL,混匀后进行PCR扩增。PCR反应条件为94℃,预变性3min后进入以下循环:94℃变性 1 min,55℃退火 1 min,72℃延伸 2 min,30个循环后于72℃下10 min,并在4℃下保存备用。PCR反应结束后进行10g/l琼脂糖凝胶电泳。
1.2.5 扩增产物纯化及测序鉴定与系统发育树的构建
利用Biospin胶DNA回收试剂盒对DNA进行纯化扩增并电泳。染色后在凝胶成像系统紫外灯下观察。从NCBI数据库中下载公认Botryosphaeria dothidea序列为参考,参比菌株具体信息为KT182890.1,FJ493245.1,KF294012.1,MH715242.1,JX241645.1与供试菌株相应序列一起采用Mega5.1中ClustalW进行比对并校正。轮纹病菌以Fusarium oxysporum(JK747249.1 4-C10)为外源,基因序列自展法(Bootstrap)检测,重复 1000次,构建邻接树(Neighbor-Joining Tree)。腐烂病菌以Botryosphaeria dothidea序列为外源,其他方法相同。
1.2.6 致病性测定
采用菌饼接种法[3]。选取PDA培养基上生长5天的菌饼采取刺伤接种健康苹果果实上,以不接种病菌的苹果果实为对照,重复15次,5天后观察接种结果。
1.2.7 病菌生理学特性研究
1.2.7.1 病菌对不同温度的适应性
将海棠轮纹病和腐烂病病菌分别接种于PDA培养基,打孔器内径为0.5mm,分别于10℃、15℃、20℃、25℃、30℃、35℃和 40℃恒温培养箱中培养,每隔一天后测量菌落直径,每个菌株重复3次。
1.2.7.2 病菌对不同pH的适应性
将湖北海棠轮纹病菌接种于pH 为 3、4、5、6、7、8、9、10和 11的 PDA培养基上,打孔器内径为0.5mm,然后置于恒温培养箱中培养,每隔一天后测量菌落直径,每个菌株重复3次。
2 结果与分析
2.2 病原菌的分离
选取的15块病组织经分离纯化共获得15个菌株,放于25℃恒温箱备用。
2.3 病原菌的鉴定
2.3.1 培养学特性
2.3.1.1 海棠轮纹病菌培养学特性观察
海棠轮纹病菌在PDA培养基上生长迅速,培养初期菌落为白色,疏松,棉絮状,气生菌丝较发达,菌落边缘整齐或不整齐。培养3d后,菌落逐渐变成灰色,10d左右,菌落呈深灰色至黑色。显微镜检发现,菌丝初期无色,后为深褐色,菌丝粗细不匀,分枝发达,有隔(如图1)。
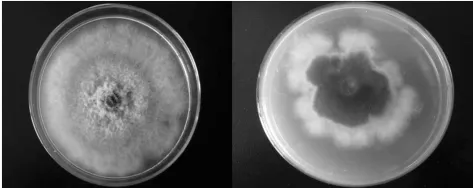
图1 海棠轮纹病菌的培养学特征(5d)
2.3.1.2 泰山海棠腐烂病菌培养学特性观察
海棠腐烂病菌在PDA培养基上初为白色,后变为浅黄色,菌丝匍匐,气生菌丝不发达,基内生长,菌落边缘整体,3~5天长满整个平皿,此后菌丝继续向培养基内生长,后期20天长出褐色的分生孢子器或者直接溢出分生孢子堆(如图2)。
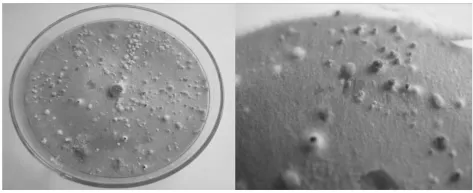
图2 海棠腐烂病菌的培养学特征(20d)
2.3.2 形态学特性
2.3.2.1 海棠轮纹病菌镜检形态学特性
解剖镜下镜检发现,海棠轮纹病菌分生孢子器呈黑褐色,初埋生于寄主表皮下,后突破表皮部分外露。分生孢子多为单生,单腔室,扁球形或球形,有孔口,大小约 110.12~154.28μm ×82.74~150.23μm,平均 129.67~114.27μm。内壁生分生孢子梗,孢子梗无色,不分支。分生孢子梭形,正直,无色,单孢,顶端钝圆,基部平截,大小为16.83~24.55μm ×5.27~8.17μm,平均 18.35 ×6.34μm(如图3)。
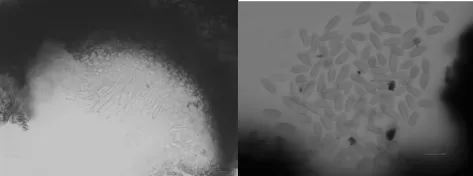
图3 海棠轮纹病菌在解剖镜下镜检形态特征
2.3.2.2 泰山海棠腐烂病菌镜检形态学特性
解剖镜下镜检发现,腐烂病菌分生孢子器着生于外子座内,扁瓶状,480~1600μm× 400~960μm,多个腔室,相通,同一孔口,室壁密生分生孢子梗;分生孢子单孢,腊肠形或香蕉形,3.6~8.0μm×0.8~1.7μm。孢子溢出形成橘黄色孢子角(如图4)。
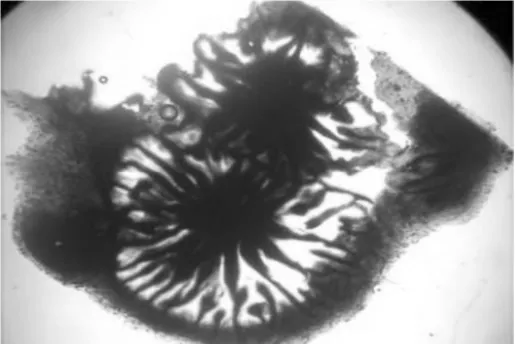
图4 海棠腐烂病菌形态特征
2.3.3 泰山溃疡病病原菌的分子鉴定
2.3.3.1 泰山海棠轮纹病菌ITS序列测定及系统树构建
海棠轮纹病菌经PCR检测,其大小介于500-750bp之间,序列测定发现海棠轮纹病菌的ITS序列长583bp。经Blast软件分析对比,分子鉴定认为泰山海棠轮纹病是子囊菌门(Ascomycotina),葡萄座腔菌(Botryosphaeria dothidea),其系统树构建如图5所示。

图5 基于ITS基因序列的海棠轮纹病菌系统发育树
由图5看出,所测定的序列确与Botryosphaeria dothidea真菌的系统发育最近,再结合病菌的培养学和形态学特征,可以明确泰山海棠轮纹病菌为Botryosphaeria dothidea。
2.3.3.2 泰山海棠腐烂病菌ITS序列测定及系统树构建
海棠腐烂病菌系统树构建如图6所示。
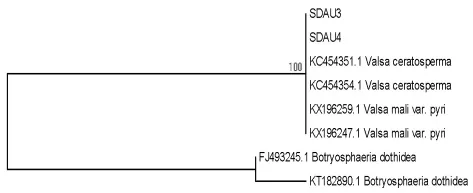
图6 基于ITS基因序列的海棠腐烂病菌系统发育树
由图6看出,海棠腐烂病菌经过序列测定和系统树构建并结合病菌的培养学和形态学特征,鉴定泰山海棠腐烂病是由Valsa ceratosperma真菌引起。
2.4 海棠溃疡病病原菌的致病性测定
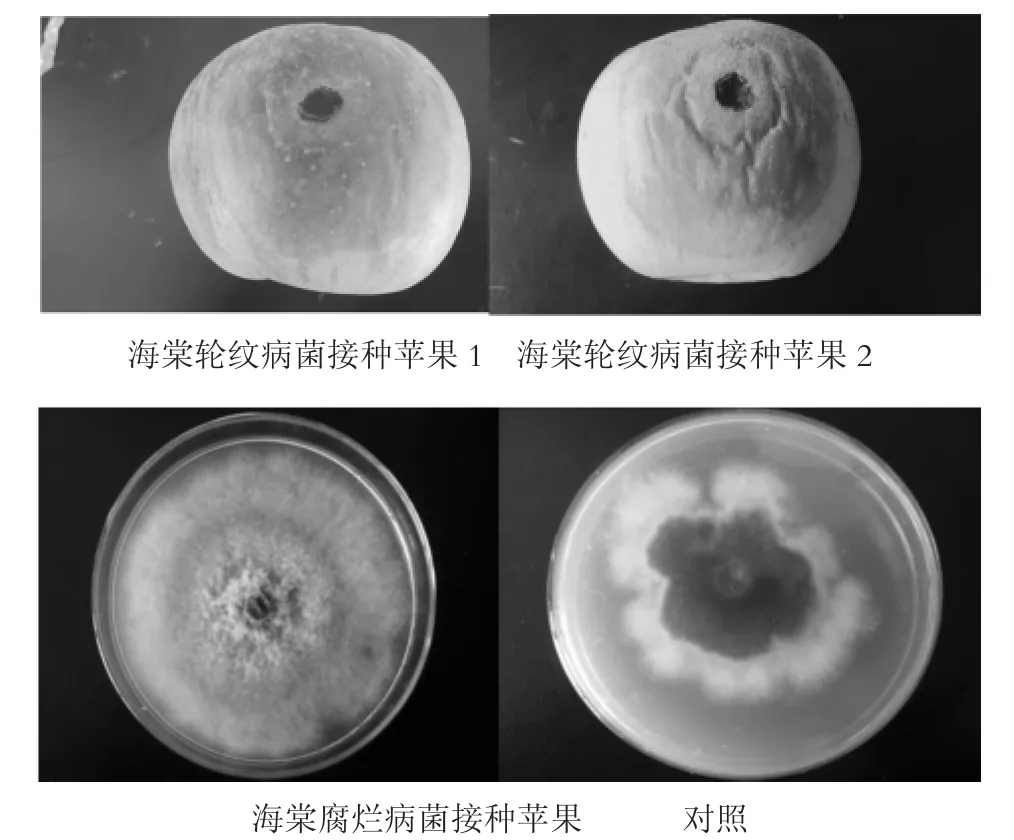
图7 海棠轮纹病和腐烂病菌接种苹果后的不同症状
室内人工接种苹果果实(如图7)发现,分离的2种病原菌对苹果果实均有致病性,其发病症状和苹果轮纹病、腐烂病病症表现一致。再次证明Botryosphaeria dothidea是泰山海棠轮纹病的病原菌;Valsa ceratosperma是泰山海棠腐烂病的病原菌。
2.5 病菌对温度的适应性
在不同温度下的PDA培养基上,湖北海棠轮纹病菌和腐烂病菌的生长特性如图8和表1所示。
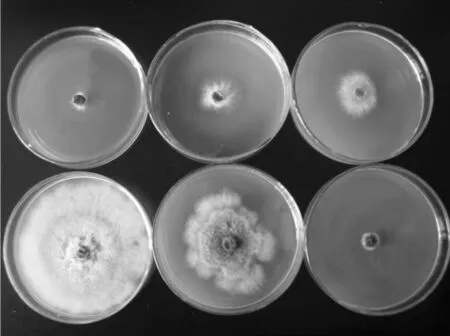
图8 湖北海棠轮纹病菌在不同温度下的生长特性(3d)(从左到右从上到下依次是在 10℃、15℃、20℃、25℃、30℃和35℃轮纹病菌的生长状况)
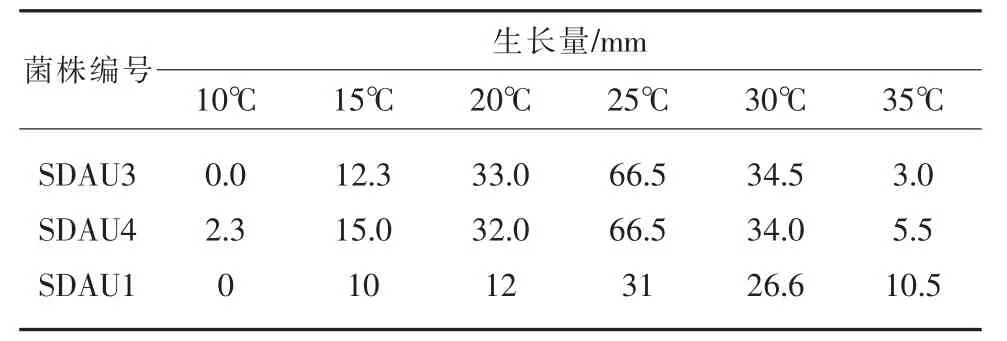
表1 湖北海棠轮纹病菌和腐烂病菌不同温度生长情况/mm
由图8和表1可以看出,泰山海棠轮纹病菌和腐烂病菌在10℃~35℃条件下均可生长,随着温度的升高,两种病菌的生长速度加快,病菌在25℃生长最好,其次是30℃、20℃和15℃,其他温度下病菌的生长速度明显减慢,腐烂病菌的耐高温能力稍强于轮纹病菌。对温度的适应性更强一些。
2.6 病菌对不同pH的适应性
泰山海棠轮纹病菌和腐烂病菌在25℃下,不同pH的PDA培养基生长结果见表2。由表4可知,不同pH条件下,海棠轮纹病菌和腐烂病菌生长特征存在差异,海棠轮纹病菌的生长范围稍窄一些,在pH4~10之间;而海棠腐烂病菌在pH3~11之间均可生长。随着pH的升高,两种病菌的生长速度加快,其中腐烂病菌在pH6时生长最快,轮纹病菌在pH5生长最快,总体表现为轮纹病菌生长稍慢于腐烂病菌生长,说明海棠腐烂病菌对酸碱度的适应性更强一些。
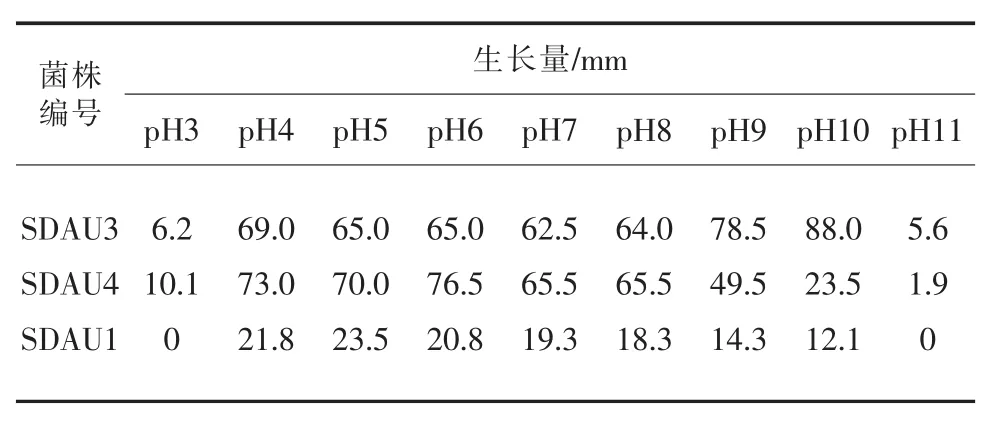
表2 湖北海棠轮纹病菌和腐烂病菌不同pH生长情况/mm
3 结论与讨论
通过海棠枝干溃疡病原菌的培养学、形态学、分子序列特征、系统树构建及致病性测定表明,海棠枝干溃疡病主要由轮纹病和腐烂病两种病害所致,鉴定病原菌2种,其中海棠轮纹病原菌为葡萄座腔菌(Botryosphaeria dothidea);海棠腐烂病原菌为 Valsa ceratosperma。生理学研究表明,两种病菌对温度和pH的要求均随着温度或者酸碱度提高而逐渐生长加速,但不同病菌的适应性不同,其中腐烂病菌对温度和酸碱度的适应性更强一些。2种病菌造成湖北海棠枝干溃疡病的发生在国内鲜有报道,尤其是大面积分布在泰山高海拔区1300m处的湖北海棠林分。本研究明确了湖北海棠枝干溃疡病原菌发生的种类及致病病原菌,为下一步综合治理湖北海棠枝干溃疡病提供了理论支持。
猜你喜欢
疯狂英语·新策略(2021年9期)2021-11-02
烟台果树(2020年1期)2020-12-17
河北果树(2020年4期)2020-11-26
河北果树(2020年4期)2020-11-26
河北果树(2020年2期)2020-01-09
热带作物学报(2019年4期)2019-06-11
今日农业(2019年14期)2019-01-04
中国森林病虫(2018年4期)2018-09-19
现代园艺(2018年3期)2018-02-10
现代园艺(2018年14期)2018-01-18
