
冷藏鲈鱼片优势腐败菌的分离鉴定及致腐能力分析
2019-09-04 09:54赵宏强蓝蔚青孙晓红刘书成
中国食品学报 2019年8期
赵宏强 蓝蔚青* 孙晓红 刘书成 谢 晶
(1 上海海洋大学食品学院 上海水产品加工及贮藏工程技术研究中心上海冷链装备性能与节能评价专业技术服务平台 食品科学与工程国家级实验教学示范中心(上海海洋大学)上海201306 2 广东海洋大学食品科技学院 广东湛江524088)
鲈鱼(Lateolabrax japonicus)又名鲈板、花鲈、四肋鱼等,属鲈形目鲈科鲈属经济鱼类,主要分布于太平洋西部与中国沿海,其中又以我国黄海、渤海较多。鲈鱼肉质鲜嫩,营养价值丰富,其体内富含蛋白质、VA、VB、钙、镁、锌、硒等营养元素,具补肝肾、益脾胃、化痰止咳功效,深受广大消费者喜爱[1]。然而,由于鱼肉中的营养物质与水分含量较高,鱼体组织易受内源酶与微生物的作用,会代谢产组胺、硫吲哚、醛等腐败物质,使鱼肉产生不可接受的气味,影响其食用与销售。
水产品优势腐败菌为水产品贮藏期间,对其腐败过程起着主导作用的特定菌,其致腐能力要明显强于其它细菌,对水产品货架期有着密切关系[2]。近年来,国内外部分学者开始定量研究腐败菌致腐能力。如Dalaaard 等[3]利用腐败菌产生的腐败代谢产物作为其腐败能力的定量指标。许振伟等[4-5]进行腐败菌致腐能力定量研究时,将腐败菌接种至无菌鱼块,由贮藏过程中的感官与腐败产物变化,以腐败代谢产物——挥发性盐基氮(Total Volatile Basic Nitrogen,TVB-N)与微生物生长量间的比例关系,表征优势腐败菌的致腐能力。基于此,本文通过前期冷藏鲈鱼优势腐败菌的分离筛选,结合菌落形态观察、生理生化实验与16S rDNA 分子鉴定技术等方法对其分离菌株进行鉴定,以TVB-N 值与菌落总数作为定量指标,致腐因子YTVB-N/CFU 为定量标准,综合评价优势腐败菌菌株致腐能力,为后期研究鲈鱼优势腐败菌的作用机理提供理论参考。
1 材料与方法
1.1 原料
鲜活鲈鱼购于上海农工商港城超市,将原料保持鲜活状态在30 min 内运回实验室。
1.2 主要药品试剂
营养琼脂培养基、假单胞菌CFC 选择性培养基、VRBDA-肠道菌琼脂培养基、MRS 琼脂培养基、铁琼脂培养基,青岛高科技工业园海博生物技术有限公司;琼脂糖、PCR 扩增试剂与合成扩增引物,生工生物工程(上海)股份有限公司;天根细菌基因组DNA 提取试剂盒、PCR 产物回收试剂盒,天根生化科技(北京)有限公司等。
1.3 主要仪器与设备
DHP-9162 型电热恒温培养箱,上海一恒科技有限公司;PowerPac Basic 电泳仪、Universal Hood II 凝胶成像仪,BIO-RAD 公司;Mastercycler6325 PCR 扩 增 仪、5427R 离 心 机,EPPENDORF 公司;AUW320 型分析天平,日本岛津公司;TGL-16M 台式高速冷冻离心机,湖南湘仪实验室仪器开发有限公司;FJ200-S 数显高速均质机,杭州齐威仪器有限公司;SW-CJ-1D 型单人净化工作台,苏州净化设备有限公司;Kjeltec2300 型凯氏定氮仪,丹麦FOSS 公司;LDZM-40KCS-Ⅲ型立式压力蒸汽灭菌器,上海申安医疗器械厂等。
1.4 试验方法
1.4.1 优势腐败菌分离纯化 在无菌工作台中随机选取10 g 腐败末期的鲈鱼样品,将其置于无菌均质袋中,加入90 mL 无菌生理盐水封口后拍打2 min。取上清液1 mL,10 倍梯度稀释,选择3 个合适的稀释度,每个稀释度做3 次重复,倾注于含有不同选择培养基的平板中。培养基选择及培养条件见表1。

表1 不同微生物的选择性培养基及培养条件Table 1 Selective culture media and conditions of different bacteria
从上述培养基的平板中挑取各种典型生长的菌落,在TSA 培养基上反复进行平板划线分离,至少3 次划线分离后得到纯化单菌落[6]。最后,于TSB 肉汤中培养,30 ℃摇床培养18 h,使其菌数达到108CFU/mL 后低温保存。
1.4.2 优势腐败菌筛选 将分离纯化多次的菌株培养至对数期后,用无菌生理盐水制成菌悬液,摇匀,利用血球计数板直接测定菌液浓度,调整菌液浓度为105~106CFU/mL[7]。用无菌手术刀将鲈鱼背部肉分割成约40 g 大小均匀的肉块,先用75%酒精喷其表面,静置10 s 后用无菌超纯水清洗,然后使用酒精灯对鲈鱼表面进行灭菌,再将鲈鱼鱼块浸泡于调整过的菌液30 s 后取出放入无菌自封袋中,每种菌株接种2 个肉块,以2 个未接种菌液的肉块为对照,置于4 ℃冰箱下贮藏。
同时,参照Parlapani 等[8]法,由10 名训练有素的感官评价人员组成评定小组,分别从样品外观、气味与弹性3 个方面进行感官评价,将腐败最快肉块所接种的菌株作为筛选出的优势腐败菌[9-10],选取其中的优势腐败菌进行菌种鉴定与致腐能力测定。
1.4.3 优势腐败菌鉴定
1.4.3.1 菌落特征与菌体形态观察 在TSA 培养基上多次纯化的细菌,选取长势比较好的单菌落,通过革兰氏染色等方法对其菌落特征与菌体形态进行观察。
1.4.3.2 传统生理生化试验 根据 《常见细菌系统鉴定手册》[11]与《伯杰细菌鉴定手册》[12]对菌株进行氧化酶、产硫化氢、葡萄糖发酵、过氧化氢酶、明胶与山梨醇生理生化鉴定。
1.4.3.3 16S rDNA 鉴定 挑取重新培养18 h 的单菌落至液体培养基中过夜,采用天根细菌基因组DNA 快速抽提试剂盒直接提取DNA,于-20 ℃保存备用。

表2 鲈鱼感官评价表Table 2 Sensory evaluation standards of Lateolabrax japonicus
以提取的菌液总DNA 作为模板,采用通用引物27F (5’-AGAGTTTGATCCTGGCTCAG-3’)和1492R (5’-AAGGAGGTGATCCAGCCGCA-3’)来扩增目的基因片段。采用25 μL 体系进行PCR 扩增,即Taq PCR Master Mix (2×)12.5 μL、ddH2O 9.5 μL、上、下游引物各1 μL 及模板DNA 1 μL。PCR 扩增程序为:94 ℃预变性4 min,94 ℃变性1 min,58 ℃退火1 min,72 ℃延伸2 min,进行35 个循环;最后72 ℃延伸10 min。用1×TAE 缓冲液配制1%琼脂糖凝胶[13],取5μL 扩增产物进行琼脂糖凝胶电泳,再用溴化乙锭(EB)对其染色后,在凝胶成像仪中观察电泳结果并拍摄电泳图谱。每种菌株均获得长度1.5 kb 左右的条带,将PCR 成功的样品委托生工生物工程(上海)股份有限公司测序。
登录网站(http://blast.ncbi.nlm.nih.gov/Blast.cgi),将测序所得16S rDNA 序列与核酸序列数据库中已知序列进行同源比对,从中选取若干个相似性高的菌株序列,使用MEGA5.1 软件,通过最大似然法(Maximum-Likelihood)做1 000 次随机抽样构建系统发育树。
1.4.4 优势腐败菌腐败能力测定 将筛选所得的优势腐败菌按照1.4.2 节方法制得一定浓度的菌悬液,接种于灭菌的肉块后放入无菌自封袋中4℃贮藏。将筛选得到的优势腐败菌菌株接种到无菌的鲈鱼鱼块上,以未接腐败菌肉块作对照,贮藏在4 ℃冰箱中,定期进行TVB-N 和菌落总数的测定。将TVB-N 产量因子YTVB-N 作为各优势腐败菌腐败能力的定量指标[14]。

1.4.4.1 TVB-N 值 参照食品安全国家标准——GB 5009.228-2016 《食品中挥发性盐基氮的测定》[15]进行鲈鱼肉TVB-N 值测定。
1.4.4.2 菌落总数 参照食品安全国家标准——GB 4789.2-2016《食品微生物学检验菌落总数法》[16]进行鲈鱼肉菌落总数测定。
1.4.5 数据分析 使用MEGA 5.1 软件构建系统发育树。数据采用3 次平行试验的平均值,结果用平均值±标准偏差表示。采用OriginPro 9.0 绘制数据图。
2 结果与分析
2.1 优势腐败菌的分离筛选
2.1.1 优势腐败菌分离 对菌液采用选择性培养基,分别在不同的培养环境下培养,然后根据菌落生长情况,共选取12 株长势较好、形态典型的菌落,其中营养琼脂培养基1 株(T1),VRBDA-肠道菌琼脂培养基2 株(E1、E2),铁琼脂培养基4 株(S1、S2、S3、S4),假单胞菌CFC 选择性培养基3株(P1、P2、P3),MRS 琼脂培养基2 株(L1、L2)。
2.1.2 优势腐败菌筛选 将选取的12 株典型菌株进行24 h 摇床活化,制成适量菌液,分别接种在无菌鲈鱼样品上,以未接种菌液的灭菌样品作为对照组,分别进行感官分析。结果如图1所示。
由图1可知,贮藏初期,鲈鱼样品表面色泽鲜亮,具有固有清香,肌肉弹性坚实,指压后凹陷立即消失,样品新鲜度较好,接种细菌与未接种细菌的鲈鱼样品无差别,样品初期品质相对一致。随着货架期的延长,鲈鱼鱼块品质下降,样品表面色泽亮度降低,逐渐产生不愉快的气味,其中接种过菌液的鱼块感官分值明显下降。S1、S2、P1、P2 与P3样品的感官分值显著下降,第7 天时,S2 组样品的感官分值已腐败严重,样品表面黏液丰富,色泽暗黄,腐臭味明显,感官呈不可接受。不同的微生物由于其生物学特性不同,对营养物质的利用也有所差异,其选择性消耗鱼肉中的营养物质,造成鱼肉肉质腐败速度的差异[17]。综合感官评价结果,筛选出使鲈鱼鱼块腐败速度较快的5 株优势腐败菌,分别为S1、S2、P1、P2 与P3,后期分别进行菌种鉴定和致腐能力测定。
2.2 优势腐败菌鉴定
2.2.1 细菌形态观察 通过对上述优势腐败菌的形态观察,并作革兰氏染色后镜检,观察结果如表3所示。

图1 不同优势腐败菌对冷藏鲈鱼感官分值的影响Fig.1 Effect of different spoilage organisms on sensory scores of Lateolabrax japonicus during chilled storage

表3 主要腐败菌的菌落特征与菌体形态Table 3 Characteristics of bacterial colony and shape of strains in spoilage organisms
2.2.2 细菌生理生化特征 对优势腐败菌进行氧化酶试验、明胶试验、山梨醇试验、硫化氢试验、葡萄糖氧化发酵、3%过氧化氢酶等6 种传统生理生化试验,结果如表4所示。

表4 主要腐败菌的生理生化鉴定表Table 4 Physiological-biochemical characteristics of mainspoilage bacteria
根据食品微生物鉴定图谱,并结合《常见细菌系统鉴定手册》与《伯杰细菌鉴定手册》,通过镜检结果与生理生化试验综合评价,初步鉴定S1、S2为希瓦式菌属,P1、P2、P3 为假单胞菌属。冷藏鲈鱼贮藏末期的优势腐败菌分别为希瓦式菌属与假单胞菌属。
2.2.3 分子鉴定 对筛选的5 株优势腐败菌进行分子鉴定,以5 株优势腐败菌的总DNA 为模板进行PCR 扩增,并作凝胶电泳检测。
从图2可见,样品在1.5kb 附近可见清晰明亮条带,表明其PCR 扩增成功,可用于后期测序。将PCR 产物送至上海生工公司进行分子检测,将检测结果通过NCBI 软件选取同源性最高的菌株序列,用MEGA5.1 进行多重序列对比,并以Maximum-Likelihood 法构建系统发育树。
由图3发育树并结合NCBI 网站最高相似度菌种可知,P1 与假单胞菌属亲缘关系最近,P2、P3与草莓假单胞菌亲缘关系最近,S1 与腐败希瓦氏菌亲缘关系最近,S2 与波罗的海希瓦式菌亲缘性最近。综合菌落形态特征、生理生化鉴定与分子鉴定,可知5 株优势腐败菌中有1 株假单胞菌属,2株草莓假单胞菌,2 株希瓦式菌属。在前人研究中,杨宪时等[18]对罗非鱼冷藏过程菌群变化研究发现,假单胞菌是罗非鱼在0~10 ℃冷藏条件下的优势腐败菌。郭全有等[19]对0 ℃和5 ℃贮藏条件下大黄鱼货架终点时菌相研究发现,假单胞菌与希瓦式菌是冷藏养殖大黄鱼的优势腐败菌。希瓦式菌属为革兰氏阴性菌,运动杆菌,嗜冷性,是水产品中常见的典型腐败菌,其腐败能力较强,具有一定的还原能力,可将氧化三甲胺还原成三甲胺,伴随着硫化氢的产生[20]。在有氧冷藏中,鱼、贝类和甲壳类的特定腐败菌 (Specific Spoilage Organisms,SSO)多为假单胞菌或腐败希瓦氏菌,其中腐败希瓦氏菌多为海洋温带水域的SSO,假单胞菌多为热带淡水鱼的SSO。王慧敏等[21]研究了鲈鱼在冷藏条件下,不同贮藏时间的腐败菌菌群变化,发现希瓦氏菌属(Shewanella spp.)和假单胞菌属(Pseudomonas spp.) 细菌在整个贮藏时间内均存在,且随贮藏时间的延长逐渐成为优势腐败菌。

图2 5 种菌株的16S rDNA 电泳图谱(Marker:200bp)Fig.2 16S rDNA electrophoresis map of 5 strains(Marker:200bp)

图3 基于16S rRNA 序列同源性的5 菌株的系统发育树Fig.3 Phylogenetic tree of 5 strains base on 16S rRNA gene analysis
2.3 优势腐败菌的致腐能力分析
2.3.1 TVB-N 值 挥发性盐基氮(TVB-N)是指肉类及水产品在贮藏期间肉中的蛋白质等含氮物质细菌与酶的作用下,分解产生氨类和胺等挥发性碱性含氮物质,是评价水产品新鲜度的重要指标,可反映分解产物的数量,评价水产品的新鲜度[22]。由国标可知,鲈鱼肉的TVB-N 值<13 mgN/100 g 为一级鲜度,TVB-N 值>30 mgN/100 g 为已腐败[23]。
由图4可知,样品在贮藏初期的TVB-N 值为(8.16±0.26)mgN/100 g,无显著性差异(P>0.05),在一级鲜度水平,表明接种初期,样品中的腐败菌未见增多。可能由于刚接种至样品表面,细菌还未适应生长环境,并未快速生长繁殖产生代谢产物。然而,随着货架期的延长,接种腐败菌的鱼块在第3 天时开始出现显著性差异,表明此时样品腐败加剧,代谢产物增多。贮藏第7 天时,P3 组样品的TVB-N 值达(49.88±0.84)mg N/100 g,处于极度腐败,产生不可接受的刺激性气味,这与感官评价结果一致,表明P3 在贮藏期间,微生物生长繁殖较快,消耗鲈鱼营养物质产生代谢产物增多。S1,S2 组样品的TVB-N 值分别是 (42.72±1.09)mg N/100g 与(43.05±0.53)mg N/100 g,两组间无显著差异(P>0.05),可能由于两种细菌同为同一种属,其致腐能力相似。
2.3.2 菌落总数 菌落总数变化可反映蛋白质和氨基酸分解代谢情况,测定菌落总数是衡量鲈鱼腐败程度的重要参数。Al-Daqal 等[24]提出水产品可食用的菌落总数上限是6.0 lg(CFU/g)。当TVC值超过上限值,表明鱼肉腐败。
由图5可知,贮藏初期,对照组样品的菌落总数为(2.02±0.01)lg(CFU/g),符合要求。随着贮藏时间的延长,接种腐败菌鱼肉的菌落总数均明显高于对照组。S1 处理组样品在贮藏末期的菌落总数增长缓慢,可能由于营养物质的消耗,样品中的菌落生长逐渐趋于稳定,从而维持在相对稳定的数量级。第5 天时,接种腐败菌鲈鱼样品的菌落总数均超过6.0lg(CFU/g),样品已腐败。不同菌株的生长情况会随贮藏时间呈现显著差异(P<0.05),这可能与不同菌种的致腐机制有关。P1 组样品的菌落总数在贮藏末期达(9.1l±0.05)lg(CFU/g),处于高度腐败状态。从图中还可看出,各接种腐败菌样品的菌落总数在5 d 后处于生长平缓期。这可能由于鲈鱼鱼肉中营养物质随着贮藏时间的延长而逐渐消耗,导致有机酸、胺类物质等代谢产物的积累,从而抑制其生长[25]。
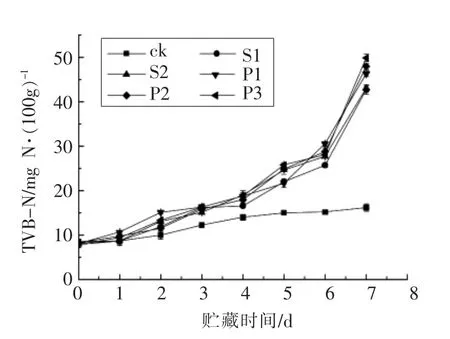
图4 冷藏条件下接种腐败菌的鲈鱼肉的TVB-N 值变化Fig.4 TVB-N values of the Lateolabrax japonicus inoculated with spoilage bacteria during chilled storage

图5 冷藏条件下接种腐败菌的鲈鱼鱼肉的TVC 值变化Fig.5 TVC values of Lateolabrax japonicus inoculated with spoilage bacteria during chilled storage
2.4 腐败能力分析
由于不同腐败菌所需营养物质不同,即生长代谢存在一定差异,因此将菌落总数与挥发性盐基氮相结合,通过产量因子Y(TVB-N/CFU)定量表征各优势腐败菌的腐败能力。
从表5可看出,通过YTVB-N可判断希瓦式菌属的致腐能力高于假单胞菌属,这可能由于希瓦式菌属在对冷藏淡水鲈鱼鱼肉的利用特性上强于假单胞菌属,其分解鱼肉中的蛋白质产生的代谢产物有所不同,鱼肉腐败程度也有所差异。现有研究表明,希瓦式菌属(Shewanella sp)是水产品腐败菌中的一种,其腐败活性强,能把氧化三甲胺还原成三甲胺,产生硫化氢等挥发性气体[26]。此外,S1的致腐因子是16.30×10-8mgTVB-N/CFU、S2 的致腐因子是14.84×10-8mgTVB-N/CFU,两者无显著差异,表明其同源性很近,这与分子鉴定结果相符合。冷藏鲈鱼的优势腐败菌中,假单胞菌属中草莓假单胞菌属的致腐能力要强于一般假单胞菌属,从P1、P2与P3 的产量因子大小可分析得出。
3 结论
通过对冷藏鲈鱼优势腐败菌的分离、筛选与鉴定,观察菌落菌落形态,做生理生化实验与16SrDNA 序列分析。所筛选的优势腐败菌中,有1株假单胞菌属(Pseudomonas sp.WCS374),2 株草莓假单胞菌(Pseudomonas fragi P121、Pseudomonas fragi B-727)和2 株希瓦式菌属(Putrefaciens Shewanella、Shewanella baltica OS185)。

表5 各种腐败菌的TVB-N 产量因子Table 5 Yield factors of TVB-N of different spoilage bacteria
将筛选所得5 株菌株分别接种到灭菌鲈鱼鱼块上,置于4 ℃条件下贮藏,通过TVB-N 产量因子(YTVB-N)比较得出,冷藏鲈鱼的优势腐败菌中,希瓦式菌属细菌的致腐能力强于假单胞菌属;假单胞菌属中,草莓假单胞菌的致腐能力强于一般假单胞菌。
猜你喜欢
中国实验诊断学(2022年5期)2022-05-27
当代水产(2022年3期)2022-04-26
实用老年医学(2021年12期)2021-12-05
华人时刊(2021年15期)2021-11-27
中国现代医药杂志(2020年10期)2020-12-14
环球时报(2019-01-07)2019-01-07
中成药(2017年9期)2017-12-19
食品工业科技(2014年23期)2014-03-11
青年时代(2012年4期)2012-08-30
