
大丽轮枝菌微丝荧光标记载体构建及应用
2019-08-27 01:52陈斌田娟冯志迪王欢李梅兰孔照胜
生物工程学报 2019年8期
陈斌,田娟,冯志迪,王欢,李梅兰,孔照胜
大丽轮枝菌微丝荧光标记载体构建及应用
陈斌1,2,田娟2,冯志迪2,王欢2,李梅兰1,孔照胜1,2
1 山西农业大学 园艺学院,山西 太谷 030801 2 中国科学院微生物研究所 植物基因组学国家重点实验室,北京 100101
微丝在真菌生长发育、胞质分裂等生命过程中具有重要功能。通过农杆菌介导遗传转化方法,将荧光mCherry标记微丝的表达载体pSULPH-Lifeact-mCherry转入大丽轮枝菌(Kleb.) 野生型V592,获得稳定的微丝荧光标记菌株V592/Lifeact-mCherry,并检测了其生物学表型和孢子萌发、菌丝生长等过程中的微丝荧光动态变化。结果表明:微丝荧光标记菌株的菌落形态、生长速率、产孢量、萌发率等表型与野生型没有显著差异;且可以观察到微丝荧光信号在分生孢子和菌丝的顶端及隔膜都有清晰定位,同时对该菌株隔膜形成过程微丝动态观察发现,微丝参与胞质分裂进程中肌动球蛋白收缩环CAR (Contractile actomyosin ring) 的形成。微丝荧光标记菌株可用于微丝在真菌发育中的动力学研究,这为深入研究微丝在真菌发育及致病过程中的作用机制提供理论与实践支撑。
大丽轮枝菌,微丝标记,生长发育,隔膜,顶端生长
大丽轮枝菌 (Kleb.) 是一种土传维管束病原真菌,它严重影响包括棉花spp.、茄子L.和莴苣L.等在内的38科660余种栽培作物和野生植物的生长[1-3]。侵染循环起始于菌丝生长接触并吸附植物根表面,菌丝形成特异侵染结构附着枝进而形成侵染钉刺穿根部表皮[4-5]。菌丝和分生孢子随后在植物的维管组织中定殖,造成维管组织系统褐变,最终导致植物死亡[6-8]。该病原菌具有致病力强、寄主广、生理小种多等特征,而且微菌核可不依赖于寄主在土壤中存活14年之久[9],发病后很难高效地控制病情蔓延而导致欠收甚至绝收,故对致病机制深入研究进而有效防治黄萎病已成为当前植物病害防控领域的研究热点。
细胞骨架微丝对丝状真菌生命活动的正常进行至关重要,包括菌丝顶端极性生长[10]、细胞分裂[11]、胞吞作用[12]和生物大分子的运输[13-14]等。其中丝状真菌顶端生长进程中囊泡可沿着微丝移动至顶体,继而被肌球蛋白沿微丝输送至顶端质膜[10];模式丝状真菌构巢曲霉隔膜形成过程中微丝与Ⅱ型肌球蛋白MyoB共定位,且微丝解聚后胞质分裂中肌动球蛋白收缩环 (Contractile actomyosin ring,CAR) 收缩和MyoB的定位均被阻断[15]。荧光蛋白标记微丝已成为研究细胞骨架结构及动态的常见细胞生物学方法[16-17]。Lifeact只有17个氨基酸大小,可与微丝稳定结合且不影响微丝动态[18]。mCherry红色荧光蛋白具有较高荧光信噪比,可用于目的蛋白定位、实时追踪以及基因表达等研究[19-22]。因此,Lifeact-mCherry荧光蛋白可作为真核生物广泛存在的微丝骨架及其互作细胞组分的示踪标记物。
真菌微丝在其胞质分裂、菌丝顶端生长、菌丝形态建成及定殖宿主等生命活动中具有非常重要的功能[23]。然而微丝在其生长发育及与宿主互作中的作用机制还不是很清楚。本研究中,我们利用农杆菌遗传转化法获得微丝荧光标记菌株V592/Lifeact-mCherry,且可以稳定清晰地观察其发育进程中微丝动态,这为发育机理、与环境互作机制及微丝互作蛋白筛选等方面的深入研究奠定了较好的基础。
1 材料与方法
1.1 材料
1.1.1 菌株及质粒
野生型V592、农杆菌AGL1菌株、质粒pentry-Lifeact-mCherry和pSULPH-mut-RG# PB均由本实验室保存,感受态大肠杆菌Trans10购自北京全式金生物技术有限公司。
1.1.2 主要试剂、培养基
Gene Ruler DNA Ladder Mix、CloneExpress®Ⅱ One Step Cloning Kit、2×PCR Master Mix、限制性内切酶RⅠ和HⅠ、PCR产物纯化和DNA胶回收试剂盒EasyPure®Quick Gel Extraction Kit、TIANprep Mini Plasmid Kit质粒小提试剂盒 (离心柱型)。培养于查氏培养基和PDA培养基;大肠杆菌和农杆菌分别培养于LB培养基和YEP培养基。所用硫酸卡那霉素(Kanamycin,Kan)的浓度为50 μg/mL,利福平(Rifampicin,Rif)的浓度为25 μg/mL[24]。
1.2 方法
1.2.1 微丝荧光标记菌株V592/Lifeact-mCherry的获得
以Lifeact-mCherry-F/Lifeact-mCherry-R从质粒pentry-Lifeact-mCherry中PCR扩增目的基因片段 (引物见表1,在5′端和3′端分别引入HⅠ和RⅠ酶切位点,在插入片段正反向引物的 5′端各引入约15 bp的线性化载体两末端同源序列);质粒载体pSULPH-mut-RG#PB使用限制性内切酶HⅠ和RⅠ进行双酶切线性化后,用One Step Cloning Kit将克隆所得的目的基因片段连入表达载体;菌落PCR鉴定为阳性克隆的菌落,提质粒酶切鉴定或测序进一步确认。电击转化法将连有目的片段的重组质粒转入农杆菌AGL1中,取阳性克隆进行培养;利用农杆菌介导的遗传转化(-mediated transformation,ATMT)方法将pSULPH-Lifeact- mCherry微丝标记表达载体转入野生型V592,单孢分离后经抗生素筛选获得微丝荧光标记菌株V592/Lifeact-mCherry。
1.2.2 微丝荧光标记菌株表型鉴定
菌株生长速率测定:将野生型菌株V592和微丝荧光标记菌株V592/Lifeact-mCherry接种于PDA培养基培养7 d,用打孔器取0.5 cm菌块放置于新的PDA培养基的中央,26 ℃黑暗培养,每3 d测量一次菌落横纵直径并计算平均值。每个菌株均重复6次。
菌株产孢量统计:将V592和V592/Lifeact- mCherry接种于PDA培养基上培养7 d,打孔器在各自菌株的菌落边缘同一位置取0.5 cm的菌块置于1 mL无菌双蒸水中,迅速振荡使分生孢子从培养基上洗脱下来并均匀分布于无菌水中,用血球计数板统计分生孢子浓度。每个菌株均重复6次。
菌株萌发率统计:用液体查氏培养基将不同类型菌株的分生孢子从PDA培养基上洗脱下来,利用血球计数板调整分生孢子浓度至106孢子/mL左右,将孢子悬浮液滴于载玻片上,26 ℃培养12 h统计分生孢子萌发情况。每个菌株均重复 6次。
1.2.3 微丝荧光标记菌株的分生孢子与营养生长菌丝中微丝荧光信号观察
用液体查氏培养基将分生孢子从PDA培养基洗脱下来并稀释至106孢子/mL;取10 μL孢子液滴于载玻片,盖上盖玻片,四周封口,然后激光共聚焦显微镜观察孢子中微丝定位情况;取20 μL孢子液滴于培养皿中央,盖上培养皿,封口,置于26 ℃的恒温培养箱里保湿培养一定时间后观察菌丝中微丝分布情况。
1.2.4 数据分析
GraphPad Prism7、Image J软件进行实验数据分析与作图。

表1 文中所用引物列表
The underlined line sequences indicate restriction enzyme site and the wavy line indicate the homologous sequence on the vector.
2 结果与分析
2.1 V. dahliae微丝荧光标记菌株V592/ Lifeact-mCherry的获得
以质粒pentry-Lifeact-mCherry为模板PCR扩增出最终带有HⅠ和RⅠ的酶切位点的基因片段,琼脂糖凝胶电泳显示基因编码序列和酶切位点全长为816 bp,说明基因克隆成功。pSULPH-mut-RG#PB载体质粒进行双酶切后用ExnaseⅡ酶进行一步法连接,重组产物转化大肠杆菌后菌落PCR验证阳性克隆,进一步测序验证。重组质粒通过电击转化法转入农杆菌AGL1,抗生素筛选获得阳性单克隆。农杆菌介导T-DNA随机插入法将基因片段插入基因组,单孢分离后经抗生素筛选获得微丝荧光标记菌株V592/Lifeact-mCherry。
2.2 微丝荧光标记菌株的菌落形态和生长速率与野生型V592相近
如图1A所示,PDA培养基培养16 d的微丝荧光标记菌株V592/Lifeact-mCherry和野生型菌株V592的菌落形态无明显差异,都有发达的白色气生菌丝和致密的黑色微菌核。从图1B可看出,微丝荧光标记菌株的菌落生长速率与野生型菌株相似;微丝荧光标记菌株最终直径约为8.23 cm,而野生型菌落最终直径约为8.17 cm,二者之间无显著差异 (>0.05)。综上,我们认为微丝荧光标记菌株在PDA培养基的菌落形态和生长速率与野生型菌株相近。
2.3 微丝荧光标记菌株产孢量、萌发率都与野生型V592无差异
对微丝荧光标记菌株V592/Lifeact-mCherry和野生型菌株V592产孢量和萌发率进行了统计。如图2A所示,微丝荧光标记菌株产孢量约为 (3.73±0.58)×106孢子/mL,约相当于野生型的91.60%,检验对二者产孢量进行差异分析,说明微丝荧光标记菌株和野生型菌株的产孢量没有显著差异 (>0.05)。如图2B所示,微丝荧光标记菌株萌发率约为93.86%,约为野生型的97.90%,检验对二者萌发率进行差异分析,说明微丝荧光标记菌株和野生型菌株的萌发率也没有显著差异 (>0.05)。综合以上结果表明微丝荧光标记菌株产孢量、萌发率都与野生型V592无差异。
2.4 微丝荧光标记菌株分生孢子和营养菌丝中的微丝分布
细胞骨架微丝即肌动蛋白丝参与真菌顶端生长、胞质分裂、细胞内物质运输等细胞活动。为探究中微丝分布情况,我们用激光共聚焦显微镜观察了微丝荧光标记菌株发育过程中微丝的亚细胞定位。如图3所示,分生孢子中微丝主要以点状分布于将要萌发的孢子顶端细胞活动旺盛区域。如图4所示,营养菌丝生长过程中微丝主要沿菌丝生长方向分布于菌丝细胞两侧内壁,且在顶端有聚集现象,或 呈环状分布于正在形成的隔膜。从图5可看出,菌丝胞质分裂过程中微丝从菌丝细胞膜内侧两端不断聚合从而参与肌动球蛋白收缩环的组装;接着环开始渐渐收缩并在第6 min时微丝相对荧光强度达到峰值,继而环逐渐降解荧光强度也随着下降,整个过程约持续14 min。
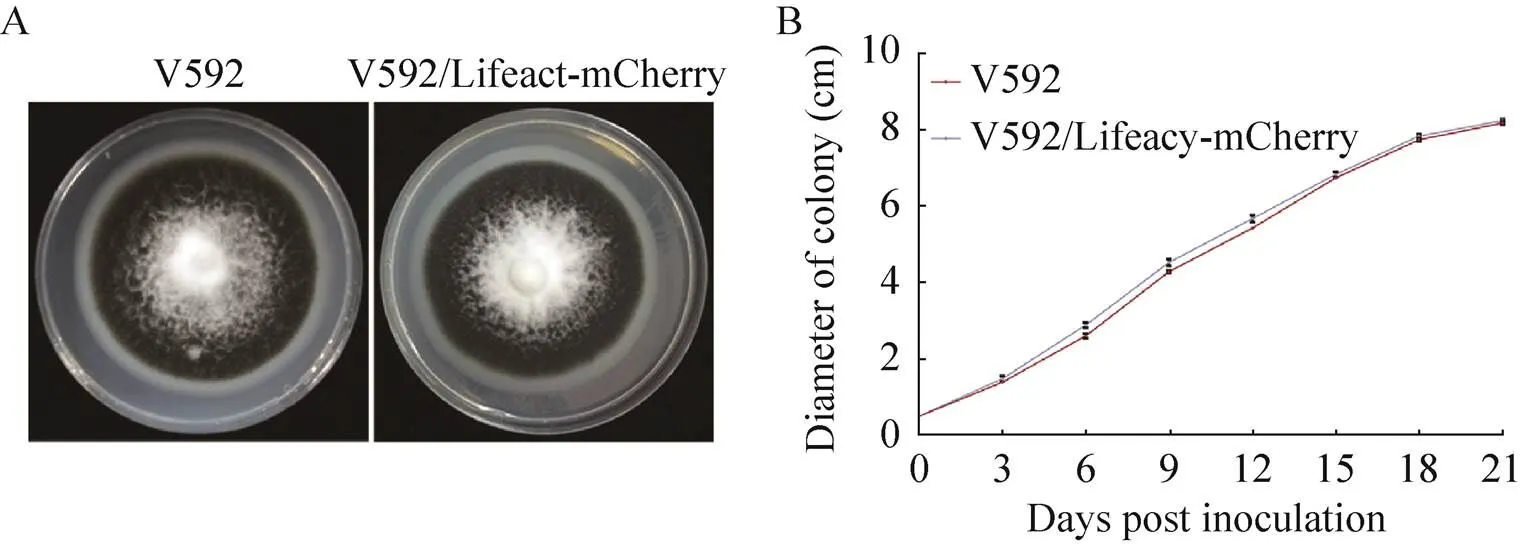
图1 野生型与微丝荧光标记菌株的菌落形态与生长特性
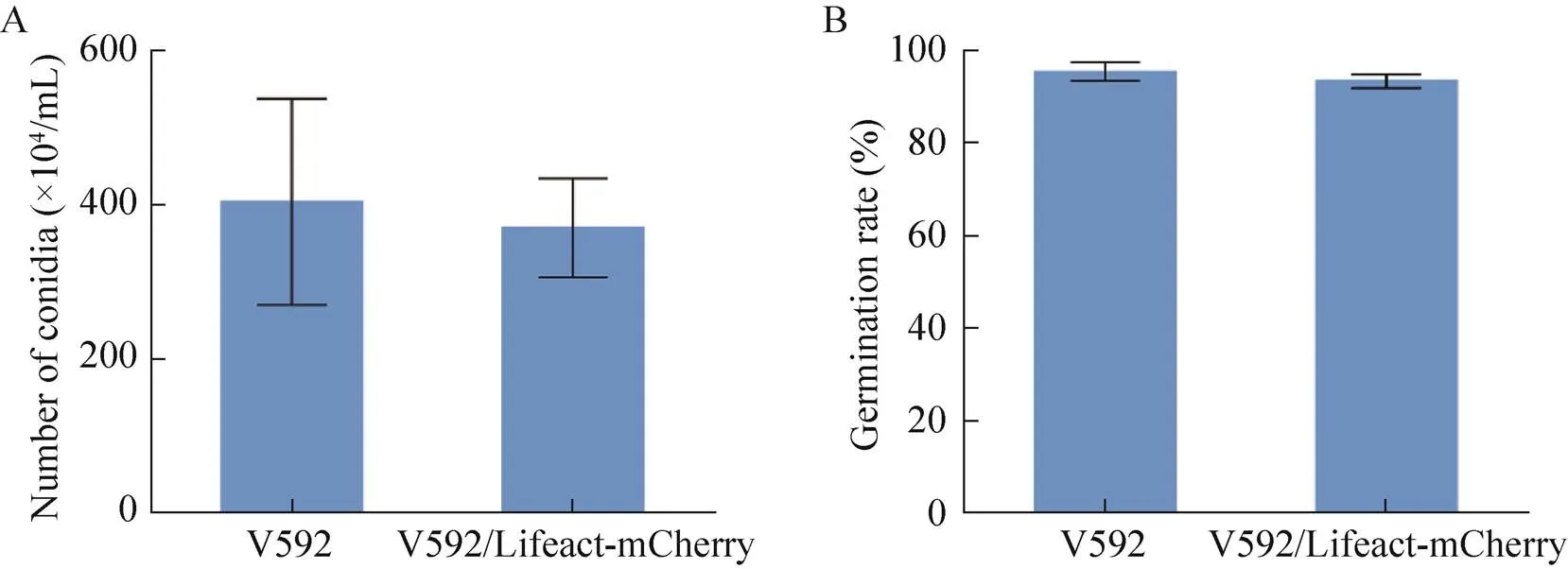
图2 野生型与微丝荧光标记菌株的产孢量和萌发率
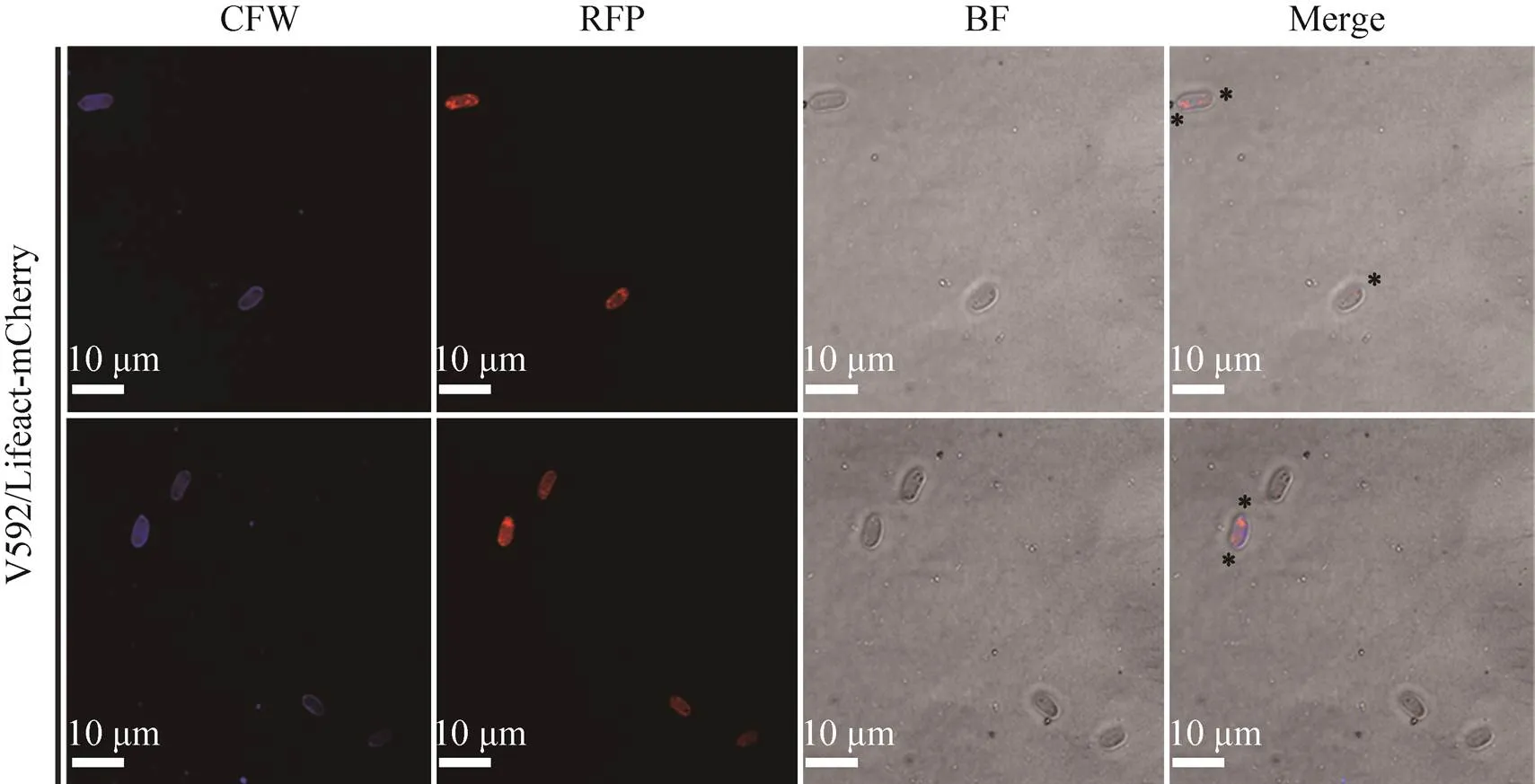
图3 微丝荧光标记菌株孢子中微丝的亚细胞定位
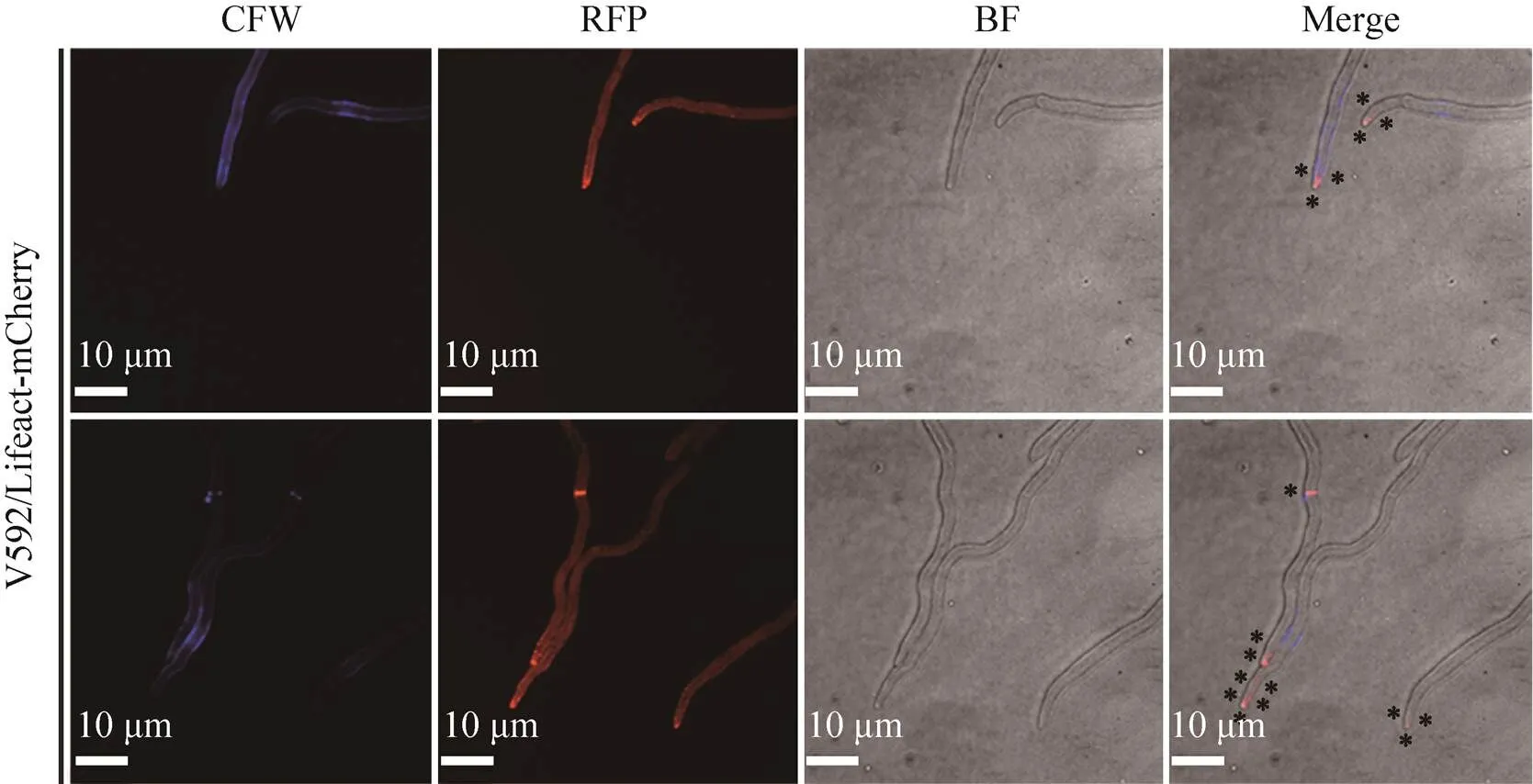
图4 微丝荧光标记菌株菌丝中微丝的亚细胞定位

图5 微丝荧光标记菌株菌丝隔膜形成过程中微丝动态及微丝相对荧光强度
3 讨论
是棉花等众多重要双子叶植物最具威胁的病原之一,所以的生长发育及致病机制一直是研究热点。真菌微丝在维持菌丝顶端极性生长、胞质分裂、胞吞及与宿主互作等一系列细胞活动中具有重要的生物学功能[13, 25]。鬼笔环肽微丝染色法具有操作复杂、毒性较大、抑制微丝功能等局限,因此我们构建微丝标记表达载体pSULPH-Lifeact-mCherry,利用农杆菌介导遗传转化野生型V592获得稳定的微丝荧光标记菌株V592/Lifeact-mCherry。标记菌株的菌落形态、生长速率、产孢量、萌发率等表型与野生型V592没有显著差异 (图1和图2)。因此,微丝荧光标记菌株可以用来观察生长发育中微丝动态。此外,使用激光共聚焦显微镜观察了微丝在分生孢子和营养菌丝中的分布情况。
微丝骨架对菌丝顶端极性建立与维持是必要的。肌动蛋白及其结合蛋白如冠蛋白等和肌动蛋白马达蛋白肌球蛋白对真菌顶端生长至关重要,肌动蛋白、冠蛋白、丝束蛋白、Arp2/3 (Actin-relatedprotein 2/3,Arp2/3) 复合物等共同组成内吞颈圈[26],构巢曲霉 ()Ⅰ型肌球蛋白MYOA在芽管顶端与肌动蛋白共定位,且在菌丝极性生长与分泌中起关键作用[27];而Ⅴ型肌球蛋白MyoE在顶体的定位依赖于微丝,且对菌丝形态起重要作用[28];且Ⅴ型几丁质合成酶(Chitin synthase with a myosin motor-like domain,CsmA)通过MMD (Myosin motor-like domain,MMD) 结构域与微丝直接相互作用,参与丝状真菌极性生长过程细胞壁合成和菌丝形态建成[29]。
本研究发现中微丝主要以点状分布于将萌发状态孢子顶端或菌丝顶端的细胞活动旺盛区域 (图3和4)。这可能与细胞顶端极性生长过程中活跃的微丝动态重构有关,菌丝顶端生长涉及的内吞作用和细胞壁合成相关的酶等物质的极性运输两大过程中都离不开微丝的参与。
胞质分裂对绝大多数生物生存都极其重要,它需要细胞信号、细胞外基质重构等活动协调作用,出芽酵母细胞分裂过程包括CAR的组装、收缩和解体、隔膜的形成、脱落及其时空协调[30]。肌动蛋白是肌动球蛋白收缩环组装与维持所必需,在胞质分裂过程中起关键作用[31-34]。研究表明,裂殖酵母肌动蛋白丝和Ⅱ型肌球蛋白是胞质分裂中收缩环动力来源的保守组分[31];中肌动蛋白丝从胞质分裂节点开始延伸并且形成束进而形成收缩环,收缩环收缩与隔膜形成同步[32]。胞质分裂时肌动球蛋白环的收缩主要由肌动蛋白解聚驱动[11],肌动蛋白在芽颈处与Ⅱ型肌球蛋白结合形成CAR,然后再进行胞质分裂,CAR收缩同时开始形成初级隔膜,随后质膜向颈部内陷,CAR收缩依赖于微丝[33-34]。
丝状真菌菌丝中胞质分裂与酵母不同,它们不涉及母细胞与子细胞的分离,只是在菌丝中形成隔膜从而维持适当的细胞体表面积比,但微丝在胞质分裂CAR组装、维持、收缩等过程中的功能相对保守[15,23,31-32,35]。模式丝状真菌稻瘟菌中CAR首先从菌丝两侧的细胞壁隔膜起始位点开始组装,随着隔膜的形成,收缩环慢慢向中心聚集,隔膜完全形成后环会在隔膜中心部位形成一个亮点,之后渐渐解体消失[36]。同样,禾谷镰孢菌、粗糙脉孢菌胞质分裂CAR的动态过程也类似[35, 37]。胞质分裂是真菌增殖和分化等重要生命活动进程的基础。
本研究发现营养生长菌丝中微丝主要沿菌丝生长方向分布于菌丝细胞两侧内壁;或呈环状分布于正在形成中的菌丝隔膜 (图4)。我们对菌丝胞质分裂中微丝动力学进行分析发现,微丝先分布于菌丝细胞膜内侧两端;接着不断聚合且与其他相关蛋白组装成肌动球蛋白收缩环;之后环开始收缩逐渐加粗,且持续一段时间,最后慢慢降解消失 (图5)。这与Guo等和Song等的研究结论基本一致[36-37]。说明菌丝胞质分裂过程中微丝参与组装肌动球蛋白收缩环,从而为隔膜形成提供动力和轨道。
微丝荧光标记菌株V592/Lifeact- mCherry菌落形态、生长速率、产孢量、萌发率等表型与野生型V592没有显著差异,且可以稳定清晰地观察其生长发育进程中的微丝动态变化,这为深入研究其发育机理及致病机制提供理论与实践基础。
[1] Klosterman SJ, Atallah ZK, Vallad GE, et al. Diversity, pathogenicity, and management ofspecies. Annu Rev Phytopathol, 2009, 47: 39–62.
[2] Xu Z, Ali Z, Hou X, et al. Characterization of Chinese eggplant isolates of the fungal pathogenfrom different geographic origins. Genet Mol Res, 2013, 12(1): 183–195.
[3] Yue YL. Identification of the pathogens causing seven kinds of agricultural and forestry plantswilt and interaction infection test[D]. Shihezi: Shihezi University, 2015 (in Chinese). 岳永亮. 七种农林植物黄萎病病原鉴定及其交互感染试验[D]. 石河子: 石河子大学, 2015.
[4] Zhao YL, Zhou TT, Guo HS. Hyphopodium-specific VdNoxB/VdPls1-dependent ROS-Ca2+signaling is required for plant infection by. PLoS Pathog, 2016, 12(7): e1005793.
[5] Klimes A, Dobinson KF, Thomma BPHJ, et al. Genomics spurs rapid advances in our understanding of the biology of vascular wilt pathogens in the genus.Annu Rev Phytopathol, 2015, 53: 181–198.
[6] Zhao P, Zhao YL, Jin Y, et al. Colonization process ofroots by a green fluorescent protein-tagged isolate of. Protein Cell, 2014, 5(2): 94–98.
[7] Yadeta KA, Thomma BPHJ. The xylem as battleground for plant hosts and vascular wilt pathogens. Front Plant Sci, 2013, 4: 97.
[8] Vallad GE, Subbarao KV. Colonization of resistant and susceptible lettuce cultivars by a green fluorescent protein-tagged isolate of. Phytopathology, 2008, 98(8): 871–885.
[9] Wilhelm S. Longevity of thewilt fungus in the laboratory and field. Phytopathology, 1955, 45: 180–181.
[10] Riquelme M. Tip growth in filamentous fungi: a road trip to the apex. Annu Rev Microbiol, 2013, 67: 587–609.
[11] Pinto IM, Rubinstein B, Kucharavy A, et al. Actin depolymerization drives actomyosin ring contraction during budding yeast cytokinesis. Dev Cell, 2012, 22(6): 1247–1260.
[12] Steinberg G. On the move: endosomes in fungal growth and pathogenicity. Nat Rev Microbiol, 2007, 5(4): 309–316.
[13] Steinberg G. Hyphal growth: a tale of motors, lipids, and the spitzenkörper. Eukaryot Cell, 2007, 6(3): 351–360.
[14] Weber I, Gruber C, Steinberg G. A class-V myosin required for mating, hyphal growth, and pathogenicity in the dimorphic plant pathogen. Plant Cell, 2003, 15(12): 2826–2842.
[15] Hill TW, Jackson-Hayes L, Wang X, et al. A mutation in the converter subdomain ofMyoB blocks constriction of the actomyosin ring in cytokinesis. Fungal Genet Biol, 2015, 75: 72–83.
[16] Kiyonari H, Kaneko M, Abe T, et al. Dynamic organelle localization and cytoskeletal reorganization during preimplantation mouse embryo development revealed by live imaging of genetically encoded fluorescent fusion proteins. Genesis, 2019, 57(2): e23277.
[17] Lippincott-Schwartz J, Patterson GH. Development and use of fluorescent protein markers in living cells. Science, 2003, 300(5616): 87–91.
[18] Riedl J, Crevenna AH, Kessenbrock K, et al. Lifeact: a versatile marker to visualize F-actin. Nat Methods, 2008, 5(7): 605–607.
[19] Ramirez-Mata A, Pacheco MR, Moreno SJ, et al. Versatile use ofstrains tagged withandgenes for the visualization of biofilms associated with wheat roots. Microbiol Res, 2018, 215: 155–163.
[20] Ransom EM, Ellermeier CD, Weiss DS. Use of mCherry red fluorescent protein for studies of protein localization and gene expression in. Appl Environ Microbiol, 2015, 81(5): 1652–1660.
[21] Xiao CW, Li L, Lao LM, et al. Application of the red fluorescent protein mCherry in mycelial labeling and organelle tracing in the dermatophyte. FEMS Microbiol Lett, 2018, 365(6): fny006.
[22] Noble S, Godoy R, Affaticati P, et al. Transgenic zebrafish expressing mCherry in the mitochondria of dopaminergic neurons. Zebrafish, 2015, 12(5): 349–356.
[23] Berepiki A, Lichius A, Read ND. Actin organization and dynamics in filamentous fungi. Nat Rev Microbiol, 2011, 9(12): 876–887.
[24] Gao F, Zhou BJ, Li GY, et al. A glutamic acid-rich protein identified infrom an insertional mutagenesis affects microsclerotial formation and pathogenicity. PLoS ONE, 2010, 5(12): e15319.
[25] Wolkow TD, Harris SD, Hamer JE. Cytokinesis inis controlled by cell size, nuclear positioning and mitosis. J Cell Sci, 1996, 109: 2179–2188.
[26] Echauri-Espinosa RO, Callejas-Negrete OA, Roberson RW, et al. Coronin is a component of the endocytic collar of hyphae ofand is necessary for normal growth and morphogenesis. PLoS ONE, 2012, 7(5): e38237.
[27] McGoldrick CA, Gruver C, May GS.ofencodes an essential myosin I required for secretion and polarized growth. J Cell Biol, 1995, 128(4): 577–587.
[28] Taheri-Talesh N, Xiong Y, Oakley BR. The functions of myosin II and myosin V homologs in tip growth and septation in. PLoS ONE, 2012, 7(2): e31218.
[29] Takeshita N, Ohta A, Horiuchi H. CsmA, a class V chitin synthase with a myosin motor-like domain, is localized through direct interaction with the actin cytoskeleton in. Mol Biol Cell, 2005, 16(4): 1961–1970.
[30] Schwayer C, Sikora M, Slováková J, et al. Actin rings of power. Dev Cell, 2016, 37(6): 493–506.
[31] Takaine M, Numata O, Nakano K. An actin-myosin-II interaction is involved in maintaining the contractile ring in fission yeast. J Cell Sci, 2015, 128(15): 2903–2918.
[32] Pollard TD, Wu JQ. Understanding cytokinesis: lessons from fission yeast. Nat Rev Mol Cell Biol, 2010, 11(2): 149–155.
[33] Lippincott J, Li R. Sequential assembly of myosin II, an IQGAP-like protein, and filamentous actin to a ring structure involved in budding yeast cytokinesis. J Cell Biol, 1998, 140(2): 355–366.
[34] Bi EF, Maddox P, Lew DJ, et al. Involvement of an actomyosin contractile ring incytokinesis. J Cell Biol, 1998, 142(5): 1301–1312.
[35] Delgado-Álvarez DL, Bartnicki-García S, Seiler S, et al. Septum development in: the septal actomyosin tangle. PLoS ONE, 2014, 9(5): e96744.
[36] Guo M, Tan LY, Nie X, et al. A class-II myosin is required for growth, conidiation, cell wall integrity and pathogenicity of. Virulence, 2017, 8(7): 1335–1354.
[37] Song B, Li HP, Zhang JB, et al. Type II myosin gene inis required for septation, development, mycotoxin biosynthesis and pathogenicity. Fungal Genet Biol, 2013, 54: 60–70.
Construction and application of actin fluorescent marker inKleb.
Bin Chen1,2, Juan Tian2, Zhidi Feng2, Huan Wang2, Meilan Li1, and Zhaosheng Kong1,2
1 College of Horticulture, Shanxi Agricultural University, Taigu 030801, Shanxi, China 2 State Key Laboratory of Plant Genomics, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
Actin filaments play an important role in fungal life processes such as growth, development and cytokinesis. The expression vector pSULPH-Lifeact-mCherry of fluorescent mCherry-labeled actin was transferred intoKleb. wild type V592 by the genetic transformation system mediated byto obtain the stable fluorescent labeled actin strain V592/Lifeact-mCherry. Then we detected its biological phenotype and the dynamic changes of actin fluorescence during the process of spore germination, mycelial growth and development. There was no significant difference in the colony morphology, colonial growth rate, sporulation and germination rate between the fluorescent labeled actin strain and the wild type. The actin fluorescence signal was observed at the tip of the conidia and hyphae and the septum clearly. Actin participated in the formation of the contractile actomyosin ring (CAR) during cytokinesis by observing the dynamic behavior of the actin in the process of hyphal septum formation. The fluorescent labeled actin strain can be used to study the dynamics of actin in fungal development to provide theoretical and practical support for further study of the mechanism of actin in fungal development and pathogenesis.
, actin marker, growth and development, septum, tip-growth
May 10, 2019;
June 3, 2019
Supported by:National Key Research and Development Program of China (No. 2017YFD0200600), Shanxi Province Key Research and Development Program Key Projects (No. 201703D211001-04-01).
Zhaosheng Kong. Tel/Fax: +86-10-64806099; E-mail: zskong@im.ac.cn
Meilan Li. Tel/Fax: +86-354-6288771; E-mail: 15935485975@163.com
国家重点研发计划项目 (No. 2017YFD0200600),山西省重点研发计划重点项目(No. 201703D211001-04-01) 资助。
2019-06-11
http://kns.cnki.net/kcms/detail/11.1998.Q.20190610.1120.003.html

陈斌, 田娟, 冯志迪, 等. 大丽轮枝菌微丝荧光标记载体构建及应用.生物工程学报, 2019, 35(8): 1520–1528.Chen B, Tian J, Feng ZD, et al. Construction and application of actin fluorescent marker in Verticillium dahliae Kleb. Chin J Biotech, 2019, 35(8): 1520–1528.
(本文责编 陈宏宇)
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
河南化工(2021年9期)2021-10-13
科学技术创新(2021年26期)2021-09-15
浙江农业学报(2021年5期)2021-05-31
合成树脂及塑料(2021年1期)2021-03-01
中国新技术新产品(2019年2期)2019-04-12
新农业(2017年15期)2017-09-11
西南农业学报(2017年6期)2017-08-08
上海农业学报(2017年4期)2017-04-10
天津医药(2016年9期)2016-10-20
