
低氧或亮氨酸干预对运动心肌Sestrin2/AMPKα2信号通路激活的影响
2019-08-21 00:56杨赟王蕴红王智强
中国运动医学杂志 2019年7期
杨赟 王蕴红 王智强
1 首都体育学院研究生部(北京100191)
2 首都体育学院运动科学与健康学院
Sestrin2 是应激诱导蛋白Sestrins 家族中的重要成员,在机体受到应激刺激,如DNA 损伤、氧化应激、缺氧等情况下诱导产生[1]。AMPK(Adenosine Monophosphate Activated Protein Kinase,磷酸腺苷活化蛋白激酶)作为一种重要的蛋白激酶参与多种代谢过程,其活性主要受AMP/ATP 比值调控。当机体处于应激状态如运动、组织缺血、缺氧等,都可以使AMP/ATP 比值升高,启动AMPK激酶系统,刺激葡萄糖转运和脂肪酸氧化等增加ATP 产生的代谢过程,同时阻断脂类、蛋白质合成以及糖异生等需消耗ATP 的代谢通路[2]。有研究发现细胞Sestrin2 的表达可显著提高能量状态感受因子AMPK Thr172位点磷酸化水平[3],且4周和6周跑台运动能引起小鼠骨骼肌Sestrin2蛋白表达和AMPKα 2 磷酸化水平显著增加,提示运动可通过Sestrin2 激活AMPKα2信号分子[4]。也有研究表明,低氧应激能够显著提高人神经母细胞瘤细胞Sestrin2 蛋白表达[5]。Sestrin2 是一种高度特异的亮氨酸传感器,机体内的亮氨酸能够通过Sestrin2 调控mTORC1 信号通路[6,7]。但目前有关运动心肌以及低氧/亮氨酸干预能否通过调控运动心肌Sestrin2/AMPKα2 信号通路来调控心肌能量代谢的研究还未见报道。为此,本实验通过建立单纯运动模型、低氧运动模型和运动补充亮氨酸模型,探讨Sestrin2/AMPKα2 信号通路在运动心肌中的时相变化以及低氧或亮氨酸干预时运动心肌Sestrin2/AMPKα2信号通路的表达规律。
1 对象与方法
1.1 研究对象
8 周雄性SPF 级SD 大鼠,由北京维通利华实验动物技术中心提供(动物合格证号SCXK(京)2015-0001)。以国家标准啮齿类动物饲料喂养,自由进食和饮水。动物饲养环境为温度23℃~25℃,相对湿度在41%~53%。自然昼夜节律变化,光照和黑暗交替各12 h(光照时间在8:00-20:00)。
1.2 建立单纯运动模型
1.2.1 动物分组
本实验分为急性耐力运动模型和4 周运动模型,80只大鼠适应性饲养后随机分组。急性耐力运动模型分为安静组(Con)、运动后0小时取材组(E0)、运动后3小时取材组(E3)、运动后24小时取材组(E24)。4周耐力运动模型分为安静组(4Con)、运动后0 小时取材组(4E0)、运动后3小时取材组(4E3)、运动后24小时取材组(4E24)。每组10只。
1.2.2 运动方案
急性耐力运动模型运动方案参照Bedford[8]渐增负荷跑台运动建立模型,只进行一次耐力运动。运动方案为:初始坡度5°,速度10 m/min,每运动5 min 速度增加5 m/min。15 min 后速度增加至19 m/min,继续运动45min。4周运动模型训练方案:适应性训练周坡度由5°递增至10°,速度为14 m/min,运动时间由15 min 递增至25 min。训练周从第1 周至第4 周坡度保持10°,速度由14 m/min 逐渐递增到24 m/min。运动时间从第1 周到第3 周由30 min 缓慢递增到60 min,并保持到训练结束。动物每周训练5天,休息2天。
急性耐力运动模型和4周运动模型在运动后按时间点取材,安静组同时取材。大鼠取材前12小时开始禁食,腹腔注射麻醉,打开胸腔取心脏,用预冷生理盐水清洗,滤纸吸净血液和生理盐水,心脏迅速称重,分离左心室,液氮冻存。所有操作均在冰上完成,样本于-80℃冰箱保存。
1.3 建立低氧运动模型
1.3.1 动物分组
60 只大鼠适应性饲养后,随机分为:常氧安静组(Con)、常氧耐力运动组(En)、常氧力竭运动组(Ex)、低氧安静组(Hypo)、低氧耐力运动组(Hypo+En)和低氧力竭运动组(Hypo+Ex)。每组10只。
1.3.2 运动方案
低氧运动模型中的常氧组在正常氧含量下安静放置或运动,低氧组在北京体育大学动物房低氧房模拟4500米海拔氧环境(氧含量约11.8%)下放置或运动,低氧安静组在低氧环境下安静放置,低氧运动组在低氧环境下运动。运动方案参照Leandro[9]渐增负荷耐力运动方案:坡度10°,初始速度5 m/min,运动4 min后速度增加5 m/min。之后每运动3 min速度增加5 m/min。耐力运动组终速度为40 m/min,总运动时间为25 min。力竭运动组速度递增到50 m/min后运动至力竭。
低氧运动模型所有运动组在运动后即刻取心脏,安静组同时取材。取材方法同实验一。
1.4 建立运动补充亮氨酸模型
1.4.1 动物分组
本实验分为补充亮氨酸急性耐力运动模型和4周运动补充亮氨酸模型,80 只大鼠适应性饲养后随机分组。急性耐力运动补充亮氨酸模型分为安静组(Con)、单纯运动组(E)、单纯补充亮氨酸组(Leu)和补充亮氨酸运动组(Leu+E)。4 周运动补充亮氨酸模型分为安静组(4Con)、单纯运动组(4E)、单纯补充亮氨酸组(4Leu)和运动补充亮氨酸组(4Leu+E)。每组10只。
1.4.2 运动方案
急性耐力运动补充亮氨酸模型运动方案同实验一急性耐力运动模型运动方案,4周运动补充亮氨酸模型运动方案同实验一的4周运动模型训练方案。
1.4.3 亮氨酸灌胃方案
补充亮氨酸模型的灌胃量参照Campos[10]补充亮氨酸大鼠模型,依据《实验动物科学》中动物与人的每公斤体重剂量折算系数表以及人类每天亮氨酸需要量计算出大鼠每天亮氨酸的需求补给量为88 mg/g,据此计算出大鼠每日灌胃量灌胃。
急性耐力运动补充亮氨酸模型大鼠在运动前10 min进行灌胃,安静对照组和单纯运动组灌胃等体积蒸馏水。补充亮氨酸组灌胃166 mg/kg,浓度83 mg/ml的亮氨酸溶液。4 周运动补充亮氨酸模型大鼠在每次运动后10 min进行灌胃,安静对照组和单纯运动组灌胃等体积蒸馏水。补充亮氨酸组灌胃166 mg/kg,浓度83 mg/ml 亮氨酸溶液。每周五下午结束末次运动后,称量记录体重并计算亮氨酸补剂量。
急性耐力运动补充亮氨酸模型的运动组在运动后即刻取材,不运动组同时取材。4周运动补充亮氨酸模型的运动组在运动后24 小时取材,不运动组同时取材。取材方法同实验一。
1.5 Western Blot
称取一定量心肌组织,按重量:体积比1∶9加入预冷的细胞裂解液,冰上匀浆,静置20 min,4℃,13000 rpm 离心20 min。取上清BCA 法蛋白定量。以裂解液调整蛋白浓度,加入5X 上样缓冲液,使样品终浓度为4 μg/μl。95℃,煮沸10 min 后,制成电泳上样液。配置12%的SDS-PAG分离胶和5%浓缩胶,加样电泳,电泳后将蛋白转至NC 膜。将膜在一抗中孵育,4℃过夜,一抗溶于封闭液,Sestrin2(1∶1000,兔抗,abcam 公司)、AMPKα2(1∶1000,兔抗,天德悦公司)、p-AMPKα(Thr172)(1∶2000,兔抗,abcam 公司)、β-actin(1∶20000,鼠抗,天德悦公司)。二抗使用山羊抗兔IgG(H+L),HRP(1∶5000,天德悦公司)和山羊抗鼠IgG(H+L),HRP(1∶10000,天德悦公司),室温孵育40 min。洗膜后采用ECL发光试剂与膜在暗室中反应曝光、显影、定影。胶片扫描后,运用ImageJ软件处理后,使用Tota-Lab Quant软件系统进行图像分析,计算目的蛋白和内参蛋白条带灰度值,并用目的蛋白的灰度值与相应的内参蛋白灰度值对比,得出每个样品目的蛋白的相对含量。
1.6 统计学方法
实验所得数据使用SPSS 进行双因素方差分析(Two-way ANOVA),数据以平均数±标准差表示,对运动和各干预因素(时间、低氧和亮氨酸)进行主效应及两者的交互作用分析。如果主效应显著(P<0.05)则采用Tukey’s test 进行事后多重比较分析(post-hoc test)。P<0.05 表示有显著性差异。
2 实验结果
2.1 耐力运动对心肌Sestrin2 表达和AMPKα2 激活的影响
2.1.1 急性耐力运动对心肌Sestrin2蛋白表达的影响
与安静组相比,运动后0 h 组心肌Sestrin2 蛋白含量显著升高(P<0.05),在运动后3 h逐渐恢复到安静水平。表明急性耐力运动能够提高心肌Sestrin2 蛋白表达(见图1)。

图1 急性耐力运动后不同时间心肌Sestrin2蛋白表达
2.1.2 急性耐力运动对心肌AMPKα2激活的影响
与安静组相比,运动后0 h组心肌AMPKα2蛋白活性显著升高(P<0.05),在运动后3 h逐渐恢复到安静水平。表明急性耐力运动能够显著激活AMPKα2(见图2)。

图2 急性耐力运动后不同时间心肌AMPKα2蛋白活性
2.1.3 4周运动对心肌Sestrin2蛋白表达的影响
经过4 周运动训练,运动后各时间段心肌Sestrin2蛋白含量与安静组相比没有显著性差异(P>0.05)。说明4周运动训练后心肌Sestrin2蛋白表达在运动后没有显著性变化(见图3)。

图3 4周运动后不同时间心肌Sestrin2蛋白表达
2.1.4 4周运动对心肌AMPKα2激活的影响
经过4周运动训练,运动后各时间段心肌AMPKα2蛋白活性与安静组相比没有显著性差异(P>0.05)。说明4周耐力运动没有激活心肌AMPKα2(见图4)。
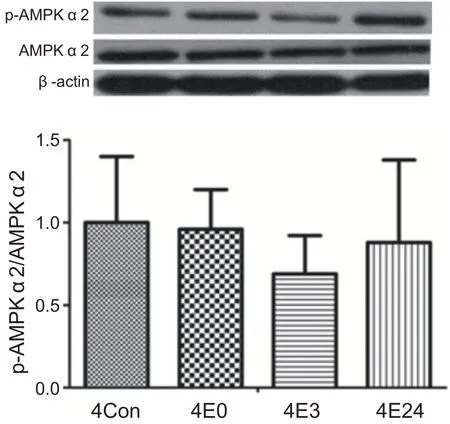
图4 4周运动后不同时间心肌AMPKα2蛋白活性
2.2 低氧运动对心肌Sestrin2 表达和AMPKα2 激活的影响
2.2.1 低氧力竭运动对心肌Sestrin2蛋白表达的影响
与安静组相比,常氧力竭运动组和低氧力竭运动组在运动后即刻心肌Sestrin2蛋白含量显著性增高(P<0.05)。低氧安静组和安静组相比心肌Sestrin2 蛋白含量没有显著性差异。表明力竭运动和低氧力竭运动能够提高心肌Sestrin2蛋白表达(见图5)。

图5 低氧力竭运动心肌Sestrin2蛋白的表达
2.2.2 低氧力竭运动对心肌AMPKα2激活的影响
与安静组相比,常氧力竭运动组和低氧力竭运动组在运动后即刻心肌AMPKα2 蛋白活性显著性增高(P<0.05)。低氧安静组与安静组相比心肌AMPKα2蛋白活性没有显著性变化。说明力竭运动和低氧力竭运动能够显著激活心肌AMPKα2(见图6)。
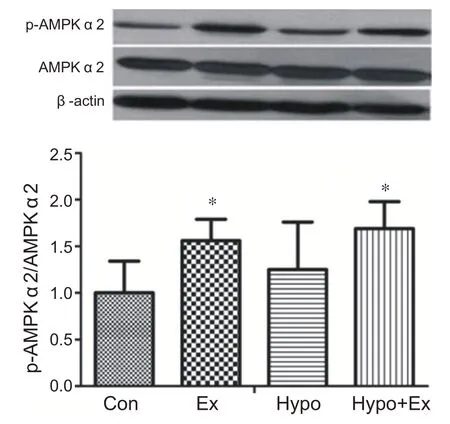
图6 低氧力竭运动对心肌AMPKα2的影响
2.2.3 低氧耐力运动对心肌Sestrin2蛋白表达的影响
与安静组相比,常氧耐力运动组、低氧安静组和低氧耐力运动组心肌Sestrin2 蛋白含量没有显著性变化(P>0.05)。说明耐力运动和低氧耐力运动对心肌Sestrin2表达没有影响(见图7)。
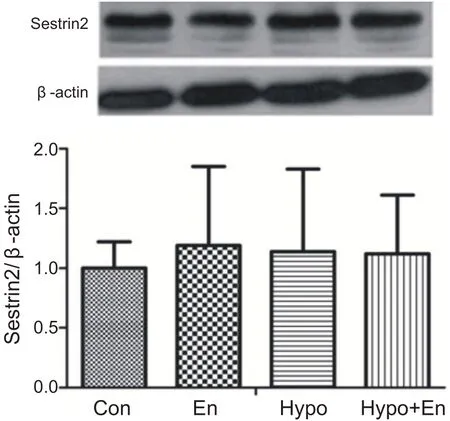
图7 低氧耐力运动心肌Sestrin2蛋白的表达
2.2.4 低氧耐力运动对心肌AMPKα2激活的影响
与安静组相比,常氧耐力运动组、低氧安静组和低氧耐力运动组心肌AMPKα2蛋白激活水平没有显著性变化(P>0.05)。表明耐力运动和低氧耐力运动没有激活心肌AMPKα2(见图8)。
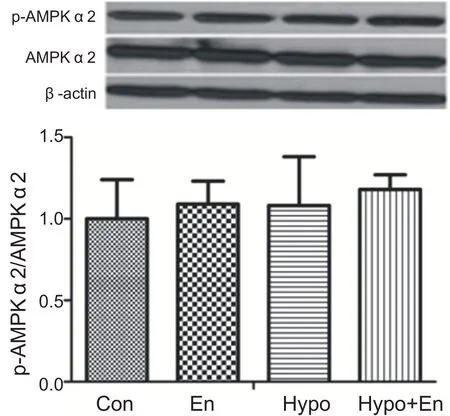
图8 低氧耐力运动运动对心肌AMPKα2激活的影响
2.3 运动补充亮氨酸对心肌Sestrin2 表达和AMPKα2激活的影响
2.3.1 补充亮氨酸急性运动对心肌Sestrin2蛋白表达的影响
与安静组相比,单纯运动组心肌Sestrin2蛋白含量显著升高(P<0.05)。补充亮氨酸急性耐力运动组与安静组和补充亮氨酸安静组相比Sestrin2 蛋白含量没有出现显著性变化(P>0.05)。说明补充亮氨酸急性耐力运动对心肌Sestrin2表达没有影响(见图9)。
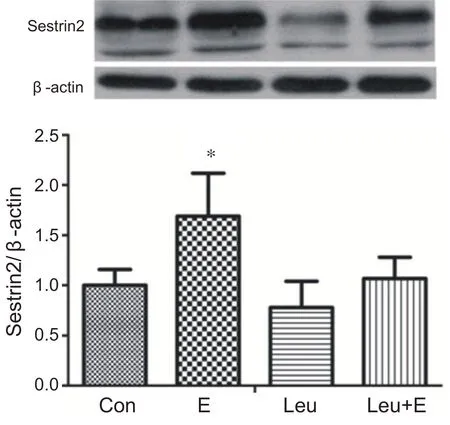
图9 补充亮氨酸急性耐力运动对心肌Sestrin2蛋白表达的影响
2.3.2 补充亮氨酸急性运动对心肌AMPKα2激活的影响
与安静组相比,单纯运动组心肌AMPKα2 活性显著升高(P<0.05)。单纯补亮氨酸组和补亮氨酸运动组与安静组相比心肌AMPKα2活性没有出现显著性变化(P>0.05)。表明补充亮氨酸急性耐力运动没有激活心肌AMPKα2(见图10)。

图10 急性耐力运动补充亮氨酸对心肌AMPKα2激活的影响
2.3.3 4周运动补充亮氨酸对心肌Sestrin2蛋白表达的影响
经过4周运动训练和补亮氨酸,单纯运动组、补亮氨酸安静组和补亮氨酸运动组与安静组相比心肌Sestrin2蛋白含量没有显著性变化(P>0.05)。表明4周运动补充亮氨酸对心肌Sestrin2蛋白表达没有影响(见图11)。

图11 4周运动补充亮氨酸对心肌Sestrin2蛋白表达的影响
2.3.4 4周运动补充亮氨酸对心肌AMPKα2激活的影响
经过4周运动训练和补亮氨酸,单纯运动组、补亮氨酸安静组和补亮氨酸运动组与安静组相比心肌AMPKα2 蛋白活性没有显著性变化(P>0.05)。表明4周运动补充亮氨酸没有激活心肌AMPKα2(见图12)。

图12 4周运动补充亮氨酸对心肌AMPKα2激活影响
3 讨论
3.1 耐力运动对心肌Sestrin2/AMPKα2 信号通路激活的影响
研究发现,细胞Sestrin2表达的升高可显著提高细胞能量状态感受因子AMPK Thr172 位点磷酸化水平[3]。也有研究发现,急性耐力运动能够显著激活心肌AMPKα2[11]。但有关急性耐力运动对心肌Sestrin2/AMPKα2信号通路的影响研究还未见报道。本研究发现,运动后即刻心肌Sestrin2 蛋白表达显著升高,说明急性耐力运动作为一种应激能够提高心肌Sestrin2 蛋白表达。同时在急性耐力运动模型中发现运动后即刻心肌AMPKα2 活性也显著升高,提示急性耐力运动中心肌能通过Sestrin2 激活AMPKα2 调节心肌能量代谢。而运动后3 小时心肌Sestrin2 的表达和AMPKα2的激活水平恢复到安静水平,说明运动对心肌Sestrin2/AMPKα2信号通路的激活是一个短期的激活。
在本实验中发现经过四周的运动训练,心肌Sestrin2的表达和AMPKα2的激活水平在运动后各时间点没有显著性差异,提示经过四周耐力运动训练心肌Sestrin2/AMPKα2信号通路表现出对运动的适应性改变。
3.2 低氧运动对心肌Sestrin2/AMPKα2 信号通路激活的影响
心肌供能以有氧代谢为主,对低氧刺激比较敏感。急、慢性低氧能使心肌糖原减少,葡萄糖代谢增强,乳酸增加,脂肪酸代谢增强等[12]。有研究表明,低氧暴露能使心肌AMPKα2磷酸化水平显著增加[13]。还有研究显示,低氧应激能够显著提高人神经母细胞瘤细胞Sestrin2蛋白表达[5]。但有关低氧应激以及低氧环境运动能否引起心肌Sestrin2 表达和AMPKα2 活性改变还未见报道。本实验结果显示,低氧力竭运动和常氧力竭运动均能使心肌Sestrin2 蛋白表达和AMPKα2活性显著升高,但两组之间没有显著性差异,并且单纯低氧对心肌Sestrin2/AMPKα2信号通路没有明显影响,表明低氧干预对心肌Sestrin2/AMPKα2 信号通路的激活作用不明显。而低氧耐力运动组心肌该信号通路激活也没有发现变化,进一步说明低氧环境下运动诱发心肌该信号通路的变化只与运动强度和时间有关,而与缺氧环境无明显的联系。尽管发生的机制尚不清楚,但有研究显示,大鼠颈总动脉分别结扎150 min 或100 min,造成两个不同程度(严重或中等)的缺氧模型,结果发现,只有严重的缺氧模型Sestrin2的表达显著升高,而中等程度缺氧模型Sestrin2表达没有变化[14],表明缺氧不是刺激Sestrin2 表达所必需的因素。另有研究显示,一些降低细胞ATP浓度的物质,如2-脱氧葡萄糖(糖酵解抑制剂)和二甲双胍(线粒体呼吸抑制剂),也能够诱导Sestrin2表达[15],提示Sestrin2表达与能量供应下降有关。此外,Sestrin2的表达虽然在一些细胞是受缺氧诱导因子(HIF-1)调控,而在另一些细胞类型,缺氧所诱导的Sestrin2 表达不依赖HIF-1,并且显示出与其他HIF-1靶基因不同的表达动力学[5,16]。从上述研究结果可以得出,Sestrin2的转录不是由缺氧本身所诱导,而可能是由缺氧引起的能量缺乏所致。因此可以推测,在本实验中,单纯低氧暴露和低氧非力竭耐力运动未引起心肌Sestrin2/AMPKα2激活,可能是由于未出现能量供应缺乏,而低氧力竭运动心肌是否出现了能量供应缺乏有待进一步研究。
3.3 运动补充亮氨酸对心肌Sestrin2/AMPKα2 信号通路激活的影响
支链氨基酸是用于运动员恢复的常用补剂[17],也是心脏摄取的主要氨基酸,虽然氨基酸在心脏ATP 的产生中比重较小,但它们对蛋白质合成和细胞信号传导等过程至关重要。其中亮氨酸是重要的供能氨基酸,能调节糖代谢和蛋白质代谢[18]。近来的研究显示,Sestrin2是一种高度特异的亮氨酸传感器[7]。当亮氨酸缺乏时,Sestrin2 与蛋白复合体GATOR2 交互作用,抑制mTORC1信号通路激活和细胞生长。而亮氨酸存在时,能够直接结合Sestrin2,破坏这种交互作用,使mTORC1 信号分子激活[6]。但有关补充亮氨酸对心肌Sestrin2/AMPKα2 信号通路作用的研究还未见报道。为此,本研究通过给予运动大鼠口服亮氨酸,观察补充亮氨酸对运动心肌Sestrin2/AMPKα2 信号通路的急性作用和长期的效应。由于实验一结果已经显示,急性耐力运动后即刻心肌Sestrin2 的表达和AMPKα2 活性提高,因此我们观察了运动前补充亮氨酸的大鼠急性运动后即刻该信号通路的变化,结果显示,心肌Sestrin2的表达和AMPKα2活性与安静组相比没有显著性变化,而未补充亮氨酸的急性耐力运动组心肌Sestrin2/AMPKα2通路活性显著性升高。提示补充亮氨酸能够抑制运动对心肌Sestrin2/AMPKα2信号通路的刺激作用。
以往的研究表明,运动使AMPKα2 激活后可能通过磷酸化TSC2 和抑制mTOR 的活性降低p70S6K 的磷酸化,进而抑制蛋白合成,促进能量底物分解供能[19,20]。AMPKα2 活性的增加还可以抑制mTOR 上游信号分子Akt 的活性[21]。而Sestrins 能通过增强AMPKα2 活化,抑制mTORC1活性,从而使细胞合成代谢的抑制和分解代谢过程如β-氧化和自噬的增加[1]。我们以往的研究发现,4 周的递增负荷耐力训练能引起Akt/mTOR信号通路的持续性激活,为此,本实验通过建立4周耐力训练模型,观察长期耐力运动后给与亮氨酸对Sestrin2/AMPKα2通路的作用,结果显示,经过4周运动结合补充亮氨酸的大鼠心肌Sestrin2 表达和AMPKα2 活性均与安静对照组相比没有显著差异。在本实验中,4周运动训练的大鼠Sestrin2/AMPKα2 通路的活性也与安静对照组没有显著差异,并且我们在相同的模型上还观察到mTOR 的活性和心重/体重也没有明显变化。这提示,所采用的4 周运动方案训练大鼠心肌与蛋白质合成相关的信号通路没有激活,而补充氨基酸也没有对Sestrin2/AMPKα2/mTOR 信号通路产生影响。是由于补充亮氨酸只对运动应激刺激引起的Sestrin2 表达升高起抑制作用,还是运动心肌对一段时间补充亮氨酸产生了适应性改变,还不十分清楚。由于目前还没有Sestrin2 的磷酸化抗体,因此有关补充亮氨酸对Sestrin2的活性的影响,以及补充亮氨酸对运动心肌蛋白质合成的调控机制还有待进一步研究。
4 结论
4.1 急性耐力运动可以一过性激活心肌Sestrin2/AMPKα2信号通路,而对长期耐力训练后心肌Sestrin2/AMPKα2信号通路表现出对运动应激的适应。
4.2 运动的负荷量与心肌Sestrin2/AMPKα2 信号通路的激活关系密切,而低氧干预对心肌Sestrin2/AMPKα2信号通路的激活作用不明显。
4.3 补充亮氨酸可抑制急性耐力运动对心肌Sestrin2/AMPKα2信号通路的刺激作用,而经过4周运动和补充亮氨酸后的心肌Sestrin2/AMPKα2 信号通路没有显著性变化。
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年8期)2022-12-07
中学生天地(A版)(2022年9期)2022-10-31
小哥白尼(军事科学)(2022年5期)2022-08-17
天津医科大学学报(2021年3期)2021-07-21
动漫界·幼教365(小班)(2020年8期)2020-07-04
中华老年口腔医学杂志(2019年2期)2019-04-28
中国体育教练员(2018年2期)2018-07-23
散文诗(2017年18期)2018-01-31
车迷(2017年10期)2018-01-18
军事体育学报(2015年2期)2015-02-27
