
The lncRNA BORG: a novel inducer of TNBC metastasis, chemoresistance, and disease recurrence
2019-07-29 08:43:48AlexGoodingKimberlyParkerSabaValadkhanWilliamSchiemann
Alex J. Gooding, Kimberly A. Parker, Saba Valadkhan, William P. Schiemann
1Department of Pathology, Case Western Reserve University, Cleveland, OH 44106, USA.
2Department of Pharmacology, Case Western Reserve University, Cleveland, OH 44106, USA.
3Department of Molecular Biology and Microbiology, Case Western Reserve University, Cleveland, OH 44106, USA.
4Case Comprehensive Cancer Center, Case Western Reserve University, Cleveland, OH 44106, USA.
Abstract Although greater than 90% of breast cancer-related mortality can be attributed to metastases, the molecular mechanisms underpinning the dissemination of primary breast tumor cells and their ability to establish malignant lesions in distant tissues remain incompletely understood. Genomic and transcriptomic analyses identified a class of transcripts called long noncoding RNA (lncRNA), which interact both directly and indirectly with key components of gene regulatory networks to alter cell proliferation, invasion, and metastasis. We identified a pro-metastatic lncRNA BMP/OP-Responsive Gene (BORG) whose aberrant expression promotes metastatic relapse by reactivating proliferative programs in dormant disseminated tumor cells (DTCs). BORG expression is broadly and strongly induced by environmental and chemotherapeutic stresses, a transcriptional response that facilitates the survival of DTCs. Transcriptomic reprogramming in response to BORG resulted in robust signaling via survival and viability pathways, as well as decreased activation of cell death pathways. As such, BORG expression acts as a (1) marker capable of predicting which breast cancer patients are predisposed to develop secondary metastatic lesions; and (2) unique therapeutic target to maximize chemosensitivity of DTCs. Here we review the molecular and cellular factors that contribute to the pathophysiological activities of BORG during its regulation of breast cancer metastasis, chemoresistance, and disease recurrence.
Keywords: BORG, chemoresistance, disease recurrence, dormancy, long noncoding RNA, metastasis, triple-negative breast cancer
INTRODUCTION
With over 250,000 newly diagnosed cases in 2017, breast cancer is the most commonly diagnosed malignancy among U.S. women[1]. Although recent advances in treatment have produced a moderate decline in the mortality rate associated with breast cancer[2], this disease nevertheless still plagues women and is the second leading cause of cancer-related death, resulting in over 40,000 deaths each year[1]. The clinical management of breast cancer is complicated by its manifestation as an exceptionally heterogeneous disease. In fact, the degree of molecular and histopathologic variation demonstrated by breast cancers necessitates its classification not as a single, uniform disease, but rather as one that is composed of a diverse collection of diseases that possess varying clinical prognoses and require distinct treatment strategies. Integral to this heterogeneity are the receptors for the ovarian steroid hormones estrogen (ER) and progesterone (PR), and for the membrane-associated tyrosine kinase HER2/ErbB-2. Moreover, the expression or lack thereof of these receptors dictates therapeutic schemes and disease-free progression[3,4]. Likewise, the multiple various permutations in the expression patterns of these receptors exemplify the heterogeneity of the disease and necessitate the tailoring of specific treatments to each individual patient. In fact, clinicopathologic detection for the expression of these receptors remains an advantageous trait, as they represent some of the most consistently predictive and actionable molecular targets in all of oncology[5]. As such, the largest clinical burden associated with breast cancer stems from a subset of patients whose tumors fail to express ER, PR, and HER2, lesions known as triple-negative breast cancer (TNBC). This genetically distinct breast cancer subtype constitutes ~15%-20% of all diagnosed breast cancers[6]and portends the worst overall survival rates of all breast cancer subtypes, an untoward trait that reflects their extreme propensity to relapse within 5 years of initial diagnosis and treatment[7]. The molecular features that underlie the development, metastasis, and relapse of TNBCs remain to be fully elucidated. Recently, a novel intergenic lncRNA known as BMP/OP-Responsive Gene (BORG) was identified as being a prominent driver of these tumorigenic activities in TNBCs. Here we highlight the pathophysiology associated with aberrant BORG expression in TNBCs, as well as discuss clinical implications of BORG and its potential for therapeutic targeting to alleviate metastatic disease.
Breast cancer metastasis
Despite immense efforts undertaken to characterize the molecular complexity of primary breast cancers, the lethality associated with all subtypes of breast cancer is attributed primarily to the dissemination and colonization of distant tissues[8,9], an untoward clinical event that results in dismal median survival rates of ~1 year. Moreover, the finding of metastatic TNBC is essentially a fatal diagnosis regardless of the chemotherapeutic intervention deployed to combat this disease[10]. Despite decades of intense investigation, a complete understanding of the molecular forces vital to metastasis remains incomplete, a knowledge gap that continues to hinder the development of therapeutics capable of specifically targeting and alleviating metastatic lesions. Nonetheless, fundamental steps taken by malignant cells to facilitate their dissemination from the primary tumor site to distant tissues for colonization have been identified and are called the metastatic cascade[11]. The events that comprise this multi-step process include: (1)local invasion and migration of malignant cells into the stroma surrounding the primary lesion; (2)intravasation into the vasculature or lymphatic system to permit transit through these circulatory routes; (3)vascular stasis and subsequent extravasation into parenchymal tissues at distant sites of colonization; (4)survival and micrometastasis formation; and (5)overt growth and metastasis formation[12]. Interestingly, the earliest phases of metastatic progression (i.e.,dissociation and emigration from site of origin viathe vasculature) represent the barriers that are most easily surmounted by primary lesions, as evidenced by the fact that tumors readily liberate thousands upon thousands of cells into the circulation each day[13,14]. However, despite the relative abundance of tumor cells traversing the vascular systems of some patients, the process of metastasis remains a supreme inefficient event. Indeed, animal modeling studies estimate that only ~0.01% of circulating tumor cells are capable of initiating some form of metastatic outgrowth[15], and as such, it is the late stages of metastatic progression that ultimately dictate the competency of disseminated cells in establishing overt metastases within the metastatic niche[12].
Dormancy: a metastatic bottleneck
The acquisition of metastatic phenotypes was originally believed to be an evolutionary consequence of advanced, late-stage disease. However, recent findings indicate that carcinoma cells comprising primary breast tumors do in fact enter the circulation months-to-years prior to the point at which the primary tumor becomes symptomatic and diagnosable[16]. As such, a large proportion of breast cancer patients already harbor disseminated tumor cells (DTCs) at their time of diagnosis[11,17,18]. Indeed, DTCs are readily detected in the bone marrow of 30%-40% of patients diagnosed with early-stage breast cancer, an event that portends a significantly worse prognosis as compared to patients whose bone marrow is free from DTCs at the time of diagnosis[19]. Interestingly, a large fraction of these DTCs initially remain clinically asymptomatic due in part to their acquisition of dormancy-associated phenotypes upon arrival to the micrometastatic niche[12].
Although diverse genetic and epigenetic analyses have begun to reveal the molecular landscape that characterizes metastatic breast cancers, only recently have these investigations been directed at and tailored for DTCs and their reactivation of proliferative programs during metastatic relapse. As such, a unified definition of dormancy remains elusive. At a cellular level, current models suggest that metastatic cells often undergo proliferative arrest upon arriving to a micrometastatic niche, a phenomenon believed to reflect the initial maladaptation of DTCs to foreign stromal environments[20]. Indeed, the stromal characteristics that impact DTC dormancy are multifactorial and encompass a diverse array of immunomodulatory and vascular endothelial cell signals (e.g.,cytokine milieu, rigidity of the microenvironment, presence of active immunosurveillance[17,21,22]) that converge on niche-localized DTCs. In doing so, these unique microenvironment signals may prove to be inhospitable to DTCs, thereby provoking cellular stress and the initiation of apoptosis as these cells struggle to cope and respond to foreign junctional and adhesive signaling networks[23-25]. Consequently, newly established micrometastatic lesions fail to propagate and expand due to: (1) deficiencies to initiate and progress through the cell cycle; and (2)propensity to undergo apoptosis at a pace that equals or exceeds the proliferative rate of DTCs. Despite these barriers, a subset of DTCs remain viable and poised to reactivate proliferative programs that result in metastatic relapse yearsto-decades after implementation of initial treatments, such as chemotherapy and radical primary tumor resection[26,27]. Clinically, extended periods of metastatic dormancy is evidenced by growth modeling studies performed on over 1,000 breast cancer patients. In doing so, two discrete peaks that define the probability of metastatic recurrence were identified: (1)one correlating with a model of continuous, slow growth of metastatic cells; and (2)one corroborating the principle that the majority of delayed relapses are indeed the result of a temporary period of dormancy prior to reactivation of proliferation programs[28]. Indeed, it is this second peak that poses the greatest threat to breast cancer patients, with ~62% of breast cancer deaths occurring 5-20 years after initial diagnosis[27]. Taken together, these findings reveal that dormant DTCs play a pivotal role in the majority of breast cancer-associated mortality, a feature that cements them as one of the most clinically relevant targets in all of oncology.
MODELS OF METASTATIC DORMANCY IN BREAST CANCER
In vitroandin vivomodels of dormancy represent critical tools for investigating the molecular mediators that impact dormant states. However, the establishment of such models pose significant challenges, as the growth and propagation of dormant cell lines is, by definition, inherently impractical. Likewise, the size and sparsity of dormant micrometastases makes their identification highly burdensome. Nonetheless, accepted models of breast cancer dormancy do in fact exist. One particularly powerful model of metastatic dormancy is the murine D2.HAN series, which consists of two cell lines that display distinct metastatic properties, namely the dormant D2.OR cell line and the highly metastatic D2.A1 cell line[29]. These cell lines were derived from a premalignant murine hyperplastic alveolar nodule implanted into the cleared mammary fat pad ofBALB/cmice, resulting in spontaneous tumors that were subsequently classified by alternations in their surface glycoprotein composition as determined by retention to the lectin, peanut agglutinin[29].
Accordingly, dormant D2.OR cells exhibit high affinity for peanut agglutinin and produce slow growing tumors that are incapable of forming disseminated tumors in either spontaneous or experimental metastasis models. In stark contrast, metastatic D2.A1 cells exhibit low affinity for peanut agglutinin and produce fast growing tumors that metastasize aggressively[30]. Importantly,in vivovideomicroscopy reveals that these D2.HAN derivatives show no differences in the mechanism, timing, and proportion of cells capable of extravasating into the lungs of mice, indicating that post-extravasation events underlie their vastly disparate abilities to undergo metastatic outgrowth[31]. Despite their inability to form secondary tumors, a large proportion of disseminated D2.OR cells readily survive the process of extravasation (i.e.,~80% at 3 weeks and ~50% at 11 weeks) and remain viable despite their non-proliferative phenotype (i.e.,Ki-67-negative)[32]. Collectively, these cellular and functional features form the crux of the dormancy-associated phenotypes exhibited by D2.OR cellsin vivo,thus establishing them as a valuable model for studying the molecular mechanisms underlying metastatic dormancy[33-35].
lncRNAs in breast cancer
Elucidating the molecular determinants of breast cancer metastasis, especially those associated with DTC acquisition of and eventual emergence from dormancy, remains a critical undertaking that is essential to the future development of therapies capable of targeting DTCs. Recently, a surprising class of molecules called long noncoding RNAs (lncRNAs) have been identified and function as potent contributors to the malignant properties of breast cancer cells. Broadly, these transcripts are defined as RNA molecules that are > 200 nucleotides in length and lack an open reading frame capable of producing a functional protein[36]. LncRNAs were originally believed to possess negligible function and exist primarily as “transcriptional noise” originating from illegitimate regulatory DNA elements. However, it is now clear that lncRNA expression patterns transpire in a highly specific cell- and tissue-dependent manner[37,38]. Importantly, noncoding RNA molecules do not require protein-coding capacity to act as powerful determinants of cell fate. Indeed, lncRNAs harbor immense intrinsic functionality within the course of cellular homeostasis and disease formation. For instance, the developmental complexity of organisms correlates more closely with the extent and diversity of the noncoding genome rather than with the collective composition of protein-coding genes[39-41].
In light of the dynamic and diverse functions attributed to lncRNAs, it is unsurprising that malignant cells, including those arising from the breast, have hijacked lncRNAs to directly and indirectly alter their proliferative, invasive, and metastatic ability[42-44]. As such, several lncRNAs have been shown to modify critical breast cancer-associated molecular pathways in a manner that transcends hormone receptor status, frequently driving the development and progression of TNBCs[45]. Similarly, ER-associated signaling pathways also regulate the expression of lncRNAs[46], including HOTAIR, whose promoter contains several estrogen response elements[47].
Moreover, induction of HOTAIR promotes the growth and metastasis of breast cancers viawidespread epigenetic reprogramming[48].
BORG
Although numerous lncRNAs have been linked to breast cancer tumorigenesis and metastasis[49], the intergenic lncRNA BORG has recently emerged as a unique and formidable regulator of the metastatic competence and survival of breast cancer cells. Originally discovered in murine C2C12 myoblast cells treated with BMP2 or BMP7, BORG is a spliced and polyadenylated ~2.8 kb transcript that shows no evidence of an open reading frame and carries multiple conserved repeat sequence elements of unclear significance[50]. Collectively, these features implicate BORG as a lncRNA, whose primary sequence has been subject to several functional analyses. For instance, BORG houses several novel pentamer motifs that are essential in facilitating its strict residence in the nucleus, representing the first demonstration of sequencebased determinants operant in dictating the subcellular localization of lncRNAs[51]. Indeed, the nuclear localization of lncRNAs directly impacts their ability to elicit widespread alterations in transcriptional networks by: (1)localizing transcription factors to specific genomic loci[52-54]; and (2)exerting gross changes in the nuclear architecture of cells[55]. Likewise, BORG oversees a host of cellular functions that are readily harnessed by breast cancer cells to enhance their tumorigenic behaviors. As will be discussed in the succeeding sections, these BORG-dependent events play an essential role in promoting breast cancer cell proliferation, chemoresistance, and survival.
Control of proliferation
In undertaking a combination ofin silicoand cell biological analyses, we recently determined that the expression of BORG directly correlates with aggressive breast cancer phenotypes, and with their metastatic competence and recurrence. Specifically, BORG liberates D2.OR cells from a state of dormancy in 3D-organotypic cultures by conferring a proliferative shift in the cell cycle from G0/G1 to G2/S[56]. Importantly, this proliferative stimulus is sufficient to enable BORG-expressing D2.OR cells to form overt metastases in the lungs of BALB/c mice[56]. Interestingly, the mitogenic properties of BORG are highly context-dependent, as they only emerge in D2.OR cells propagated in microenvironments that mimic primary and/or metastatic tumor sites (e.g.,3D-cultures). Such context-dependent activities of BORG imply that this lncRNA confers malleable phenotypes to DTCs, thus compelling them to activate adaptive signaling programs that enable their survival and outgrowth within diverse metastatic niches.
BORG as a manipulator of protein function
In searching for mechanistic insights into how BORG induces DTCs to escape from dormancy, we performed mass spectrometry analyses on proteins captured by the pulldown of biotinylated, anti-sense BORG transcripts. These analyses identified the E3 SUMO ligase TRIM28 (KAP1) as a strong binding partner of BORG[57]. TRIM28 functions as a transcriptional co-repressor and scaffolding protein for histone and DNA modifying enzymes that enhance breast cancer cell proliferation, doing so in part by suppressing the transcription of senescence promoting genes, especiallyp21andGadd45a[58-61]. Interestingly, elevating BORG expression in D2.OR cells to levels that approximate those detected in their metastatic D2.A1 counterparts dramatically downregulated the expressionp21andGadd45a, indicating that BORG may rely upon TRIM28 to confer proliferative states to dormant DTCs. Indeed, CRISPR/Cas9-mediated knockout of TRIM28 restores a dormant phenotype in BORG-expressing D2.OR cells, as does expression of mutant BORG transcripts that can no longer bind TRIM28[57]. Thus, the oncogenic activities of BORG depend upon its physical interaction with TRIM28, an event that serves as a proliferative stimulus to dormant DTCs.
The diverse range of functions elicited by lncRNAs is thought to be promoted by their unique structural diversity. Their inherent length and nucleic acid structure allow the formation of flexible, complex secondary and higher order structures that facilitate their interactions with macromolecular complexes[40]. Indeed, lncRNAs can acquire behaviors analogous to ligands, as their binding to proteins can trigger conformational changes and/or modify protein: protein interactions that dramatically impact protein activation states[62]. Accordingly, BORG enhances the function of TRIM28 to inhibit transcription by regulating the pause and release of RNA Polymerase II (Pol II)[63,64]. For instance, heterologous expression of mutant BORG transcripts that retain their capacity to bind TRIM28 remain competent to elicit Pol II promoter pausing at thep21andGadd45aloci, whereas those BORG mutants incapable of binding TRIM28 fail to impact the pausing index of Pol II at these loci[57]. Furthermore, widespread evidence indicates that lncRNAs can function as molecular scaffolds for proteins, thereby: (1)tethering cooperative proteins together to enhance their functions; or (2)localizing RNA-protein complexes to specific genomic regions through base-pair and tertiary structure interactions with DNA[41,65]. Along these lines, chromatin immunoprecipitation assays reveal that BORG enhances the binding of TRIM28 to specific genomic loci[57]. Thus, BORG promotes the metastatic outgrowth of dormant DTCs in part through its ability to promote the localization and transcriptional repressive activity of TRIM28. Although BORG directly modifies the ability of TRIM28 to suppress the expression ofp21andgadd45ain dormant DTCs, it should be noted that TRIM28 also exerts widespread alterations in the transcriptomes of a multitude of cell types[63,64,66]. To gain additional insight into the repertoire of transcriptional events coupled to TRIM28 in DTCs, we performed RNA-seq and microarray-based transcriptomic analyses on parental and BORG-expressing D2.OR organoids propagated in 3D-cultures. To assess the specific impact of TRIM28 on these dormancy-associated phenotypes (i.e.,parental dormancyvs.BORG-mediated outgrowth), we also rendered these cells deficient in TRIM28 expression. Unsurprisingly, BORG-expressing D2.OR cells harbored a transcriptional signature that deviated significantly from its parental and TRIM28-deficient counterparts. Moreover, cellular network analyses revealed specific BORG- and TRIM28-dependent transcriptional patterns that were significantly enriched for proliferative and pro-metastatic signatures[57]. Collectively, these findings establish BORG as the only known lncRNA that functions in modifying the activity and cellular localization of a transcriptional regulator (i.e., TRIM28) to confer genome-wide transcriptomic alterations that compel the reactivation of proliferative programs in dormant DTCs.
Chemoresistance
The development of therapeutic resistances continues to hamper the prolonged efficacy of standard-of-care treatment regimens. Moreover, these clinical challenges are compounded by the fact that the underlying mechanisms responsible for targeted and chemotherapeutic resistance are immensely diverse. Nonetheless, malignant breast cancer cells regularly rely on the malleable intrinsic state of cancer cells, which enables their adaptation to cytotoxic cellular stresses in order to maintain viability[67]in a manner that most closely follows the paradigm of acquired resistance[68]. Indeed, the plasticity underlying the appearance of chemoresistance is naturally permissive and reflects alterations in the epigenome. Moreover, these events are bolstered by defects in the ability of DTCs to maintain genome integrity that arise in response to aberrant cell cycle checkpoints and DNA repair mechanisms, and to increased rates of proliferation. Additionally, interactions between DTCs and the tumor microenvironment induce uniquede novomechanisms of therapeutic resistance, as cell adhesion networks (e.g.,integrins) activate a specialized survival program known as “cell adhesion-mediated” drug resistance that elicit DTC insensitivity to numerous treatment regimens[69,70]. As such, the stromal composition of the metastatic microenvironment creates a natural sanctuary for DTCs to survive therapeutic insults.
Interestingly, dormant DTCs have long been recognized for their inherent resistance to commonly used chemotherapeutic drugs[11]. These resistant traits naturally stem from the quiescent phenotype of dormant cells, which effectively abolishes the clinical utility of chemotherapeutics and cytotoxic agents that target metabolically active and dividing cells[71]. Moreover, dormant cells preferentially upregulate signaling pathways associated with cell survival, a trait stemming from their allocation of metabolic resources away from cell cycle progression as a means to remain viable in the face of environmental stressors (i.e., chemotherapy)[72]. Accordingly, chemotherapeutic treatment can select for a subset of dormant cells that are enriched for pro-survival pathways and multidrug resistance, implying that cytotoxic insults can select for a population of cells that are exceedingly equipped to instigate post-therapy relapse[73,74]. Importantly, we recently determined that BORG plays a central role in driving the development of chemoresistance in TNBCs (see below).
BORG: a novel inducer of chemoresistance
In addition to possessing enhanced proliferative abilities, BORG-expressing D2.OR cells also exhibit: (1)extensive upregulation of pro-survival and viability pathways; and (2)widespread downregulation of cell death pathways[56]. Moreover, BORG expression is highly responsive to metabolic stresses such as hypoxia and nutrient deprivation, as well as to treatment with a wide panel of chemotherapeutic drugs, including doxorubicin, hydroxyurea, docetaxel, 5-fluorouracil, and 6-thioguanine[56]. These cellular attributes of BORG are consistent with its ability to enhance metastatic outgrowth and disease recurrence. Indeed, comet assays demonstrate that heterologous expression of BORG dramatically mitigates the extent of doublestranded DNA breaks experienced by D2.OR cells in response to doxorubicin exposure. Moreover, these genoprotective features of BORG rely upon its binding to the single-strand DNA-binding protein, RPA1, which functions as an essential molecule in the repair of DNA damage[75,76]. Interestingly, the capacity of BORG to induce resistance to doxorubicin appears to supersede its ability to enhance cell cycle progression, an event that could potentially render BORG-expressing cells more sensitive to the cytotoxic activities of doxorubicin[77,78]. As such, the genoprotective effects of BORG are essential in establishing the foundation operant in mediating deadly relapse in patients with metastatic breast cancers. Moreover, as lncRNAs typically function as molecular scaffolds that facilitate the formation of protein complexes, particularly in the nucleus[79], it is tempting to speculate that BORG promotes the interaction of RPA1 with proteins critical to its repair of DNA, including additional subunits RPA2 and RPA3[80], as well as other DNA-repair associated proteins, such as BRCA2, XPA, and p53[81-83][Table 1].
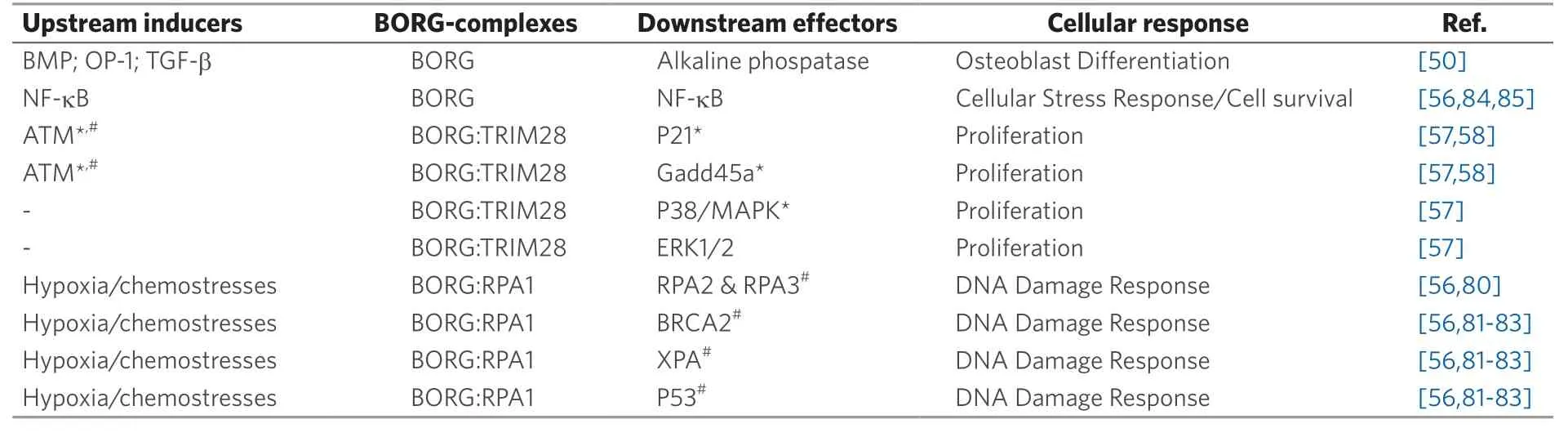
Table 1. BORG-associated signaling pathways
BORG and NF-kB: a feed-forward loop to chemoresistance
Cellular network analyses of parental and BORG-expressing D2.OR derivatives demonstrate significant enrichment of a hallmark gene signature that correlates with the induction of NF-kB activity[56]. Accordingly, NF-kB reporter assays show enhanced activation of NF-kB signaling in BORG-expressing D2.OR cells as compared to their parental counterparts. In light of the longstanding association between NF-kB activation and the initiation of pro-survival and cellular stress responses[84,85], we further explored the connections between NF-kB and BORG in eliciting DTC resistance to chemotherapy. In doing so, we inactivated the NFkB pathway viaseveral complementary strategies, including: (1)stable expression of a dominant-negative form of IkBa; (2) CRISPR/Cas9-mediated knockout of the BORG-responsive gene NEMO/IKKg; or (3)administration of small molecule inhibitors to IKKb. In all cases, inhibiting NF-kB signaling in BORGexpressing D2.OR cells restored their sensitivity to doxorubicin bothin vitroandin vivo, and to apoptosis induced by hypoxia- and nutrient deprivation[56]. It should be noted that while BORG requires ample expression of RPA1 to protect against doxorubicin-induced DNA damage and subsequent apoptosis, this event appears to be dissociated from the ability of BORG to activate NF-kB signaling. Indeed, CRISPR/Cas9-mediated disruption of RPA1 expression had no effect on the ability of BORG to activate NF-kB signaling[56]. Thus, the chemoresistant phenotypes afforded by BORG-RPA1 interactions act independently of the NF-kB signaling axis.
Interestingly, the promoter region of the BORG locus contains multiple stretches of nucleotides that exhibit sequence homology to the consensus DNA-binding sequence for NF-kB. Accordingly, NF-kB readily bound to the BORG promoter in BORG-expressing cells, thereby identifying a novel feed-forward loop whereby the activation of NF-kB (i.e.,through chemotherapeutic and environmental stressors[86-89]) leads to enhanced BORG expression, which further promotes the induction NF-kB responsive genes. Along these lines, expression of a dominant-negative IkBain TNBCs prevented their expression of BORG following exposure to doxorubicin, and to environmental stresses, such as hypoxia and nutrient deprivation[56]. Collectively, these findings implicate BORG as a unique lncRNA that is capable of promoting a NF-kB feed-forward signaling loop that effectively links metastasis-associated cellular stresses to a coordinated signaling program that engenders the survival of disseminated TNBCs.
BORG and breast cancer stem cells
Breast cancer stem cells (BCSCs) are malignant cells capable of tumor initiation, self-renewal, and differentiation into a heterogeneous group of cancer cells that reflect those present in the original primary breast tumor; they are also associated with the acquisition of metastatic and chemoresistant phenotypes[90-93], particularly upon their colonization of foreign tissue microenvironments[94]. BCSCs typically divide asymmetrically to create: (1) a single progenitor cell capable of differentiating into a variety of functionally diverse cancer cell types; and (2)a single BCSC that can expand and undergo continual self-renew. Attempts to characterize BCSCs has been hampered by a relative lack of universal markers for BCSCs[95,96]. Indeed, BCSCs have been linked to the expression of several cell surface proteins, such as CD133 and CD44+/CD24-, and to the intracellular protein, ALDH1[97-99]. Recent findings have associated the expression of lncRNAs with the generation and expansion of BCSCs. For instance, the lncRNAs ROR, HOTAIR, and Hh all induce the expression of transcription factors that regulate “stemness”, such as SOX2 and OCT4; they also impact the initiation of EMT programs[100-103], which elicit the selection, expansion, and self-renewal of BCSCs[104]. The role of BCSCs in regulating metastasis, chemoresistance, and survival signaling are reminiscent of the features attributed to aberrant BORG expression, suggesting that BORG may also regulate the behaviors of BSCSs. Accordingly, BORG expression is elevated significantly in BCSCs as compared to non-BCSCs populations; it also enhances the mammosphere forming activity of human and murine breast cancer cells (Goodinget al.[56,57], unpublished observation). Thus, future studies need to determine precisely how BORG impacts the transcriptomic and epigenetic landscapes of breast cancer cells to impart BCSC characteristics coupled to metastatic progression and disease recurrence.
DISCUSSION AND CLINICAL IMPLICATIONS OF BORG
Metastasis is an exceedingly complicated process, whereby tumor cells must undergo coordinated efforts to successfully disperse from primary tumors, emigrate to distant tissues, and survive and colonize foreign microenvironments. Even for highly transformed cells, traversing the metastatic cascade is immensely challenging, with the vast majority of disseminated cells unable to form overt metastases[15]. The inefficiencies characterizing metastasis are predominantly attributed to the cellular stresses associated with unfamiliar metastatic microenvironments, forces that obstruct both the survival and outgrowth of DTCs. Accordingly, BORG has emerged as a potent and unique lncRNA that is poised to enhance breast cancer metastasis by altering both sides of this equation, i.e.,providing both proliferative and pro-survival stimuli to DTCs.
From a potential clinical perspective, aberrant BORG expression is most frequently associated with TNBC/basal-like breast cancers, as determined by scrutinizing several publicly available RNA-seq datasets. Moreover, we detected significant elevations of BORG in metastatic human patient-derived xenograft tissue samples, and in CNS metastases and their matched primary tumors from which these metastatic foci derived. In all cases, malignant tissues clearly express increased levels of BORG as compared to normal human mammary epithelial cells[56,57]. Thus, these findings support the hypothesis that primary malignancies that house high levels of BORG will disperse aggressive breast cancer cells that are predisposed to establishing clinically-relevant, chemoresistant secondary lesions. Indeed, TNBC patients who succumbed to metastatic relapse within 5 years of initial diagnosis and treatment possessed primary tumors that contained significantly higher levels of BORG compared to primary tumors derived from TNBC patients who remained disease-free for at least 5 years post-treatment[57]. Taken together, these intriguing findings suggest that quantifying BORG expression in primary tumors could offer important insights into predicting the natural and clinical course of breast disease within TNBC patients.
The correlative finding that BORG expression largely aligns with the overall malignant propensity of breast cancer cells fails to address the mechanisms and signaling systems ultimately coupled to its upregulation in developing mammary tumors. Indeed, it has been proposed that lncRNAs evolved as a means to assist in maintaining cellular homeostasis in response to a wide variety of pathophysiologic conditions[105]. Accordingly, we show that BORG expression is similarly influenced by a number of cellular stressors, including chemotherapeutic insult, nutrient deprivation, and hypoxia[56], as well as in response to heat shock (Goodinget al.[56]and Valadkhanet al.[105], unpublished observation). Moreover, NF-kB is critical in linking the responsiveness of BORG to these environmental stressors[56], as NF-kB activation has long been tied to the survival of cells confronted with a host of intrinsic and extrinsic stressors[84,85,106]. It therefore stands to reason that increased flux through the NF-kB pathway stemming from exposure to noxious stimuli may serve as the initial impetus in triggering the aforementioned NF-kB feed-forward loop, wherein NF-kBinduced upregulation of BORG propagates expression of NF-kB-responsive gene expression patterns[56].
Tumor progression and metastatic competence are thought to rely heavily upon the intrinsic plasticity of malignant cells, which facilitates their adaptation to harsh foreign microenvironments in order to maintain viability[67,107]. Because BORG is largely regulated by extracellular paracrine factors (e.g.,TGF-b, BMP-2, and BMP-7[50,56]) and the environmental cues, we surmise that BORG acts as a context-dependent, transcriptional rheostat for disseminated breast cancer cells, thereby dictating their engagement of proliferative and prosurvival pathways. Indeed, when faced with environmental or therapeutic stresses, the induction of BORG in malignant cells orchestrates a transcriptional signature that provokes aggressive tumorigenic states that ensures for their survival. As such, preventing BORG expression or impeding the activation of its downstream targets represents an innovative and potentially impactful strategy to target metastatic breast cancers and drive them into an apoptosis-prone state.
Current mechanistic insights into the regulation of BORG and its downstream effectors reveal that this lncRNA is uniquely poised to promote the metastasis of breast cancer cells. Indeed, BORG clearly exerts a pro-metastatic effect at both the primary and metastatic sites of tumor growth. For example, a subset of breast cancer patients may harbor primary tumor cells that have gained the expression of BORG (BORGhi) as a result of environmental stresses associated with a growing primary tumor (i.e.,hypoxia and nutrient deprivation; Figure 1). Such tumors are prone to shedding BORGhicells into the circulation that disseminate to distant tissues, wherein they exploit the proliferative and pro-survival effects of BORG to overcome the hostile metastatic microenvironment and form overt metastases [Figure 1]. Accordingly, and as noted above, TNBC patients who succumbed to metastatic relapse within 5 years of initial diagnosis and treatment possessed primary tumors that contained significantly higher levels of BORG compared to primary tumors derived from TNBC patients who remained disease-free for at least 5 years post-treatment. Furthermore, breast cancer cells can emigrate from the primary tumor at very early stages of tumor development[18]. As such, these early disseminated breast cancer cells are likely to originate from a lesion experiencing little hypoxic or metabolic stress and are therefore more prone to harboring low levels of BORG (BORGlo). Although still capable of disseminating to distant tissues, these BORGlocells are predicted to struggle within their foreign microenvironments, resulting in their undergoing cell death or retreating into a state of metastatic dormancy. Nonetheless, stromal paracrine signals (e.g.,TGF-b, BMP2, or BMP7), as well as the hypoxic and metabolic stresses associated with the metastatic microenvironment, can lead to the induction of BORG expression within these dormant DTCs, thereby activating proliferative programs and survival signaling to promote their metastatic outgrowth [Figure 1].
Finally, neoadjuvant or adjuvant treatment of primary breast cancers with chemotherapeutic agents (e.g., doxorubicin) will selectively kill BORGlocells that are not inherently resistant to the cytotoxic effects of these agents. The residual, chemoresistant BORGhicells can subsequently contribute to the recurrence of metastatic or primary lesions that are insensitive to standard-of-care therapies [Figure 1]. These diverse cellular outcomes downstream of BORG establish this lncRNA as an essential driver of breast cancer metastasis and highlight the potential utility derived from therapeutically targeting BORG or its effectors as a novel means to alleviate the metastatic outgrowth and recurrence of disseminated TNBCs.
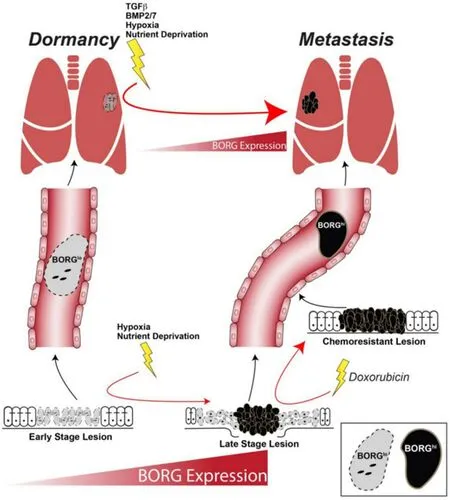
Figure 1. BORG is a potent facilitator of breast cancer metastasis. Breast cancer cells can disseminate at very early stages of development. Due to the benign microenvironment associated with these lesions, these cells are likely to harbor low levels of BORG (BORGlo) expression and are prone to establishing dormant lesions in metastatic tissues. Nonetheless, stromal factors and environmental stressors in the metastatic microenvironment can induce BORG expression, thereby compelling these dormant cells to reinstate proliferative programs. The progression of primary tumors is associated with a hypoxic environment and stark competition for nutrients. Such stresses can enhance BORG expression in a subset of cells (BORGhi). These BORGhi cells can disseminate to distant tissues where they exploit the proliferative and survival advantages afforded by BORG to produce overt metastases. Cytotoxic chemotherapeutic treatment is effective against BORGlo cells, but BORGhi cells show resistance to such therapies and are the foundation for chemoresistant, residual disease that can eventually metastasize to distant tissues. BORG: BMP/OP-Responsive Gene
DECLARATIONS
Acknowledgments
Members of the Schiemann Laboratory are thanked for critical comments and reading of the manuscript.
Author’s contributions
Conception and study design: Gooding AJ, Parker KA, Schiemann WP
Drafted and revised the manuscript: Gooding AJ, Parker KA, Valadkhan S, Schiemann WP
Availability of data and materials
Not applicable.
Financial support and sponsorship
Research support was provided in part by the National Institutes of Health (CA236273) to Schiemann WP and Valadkhan S, and (T32GM007250 and F30CA203233) to Gooding AJ. Additional support was graciously provided by the METAvivor Foundation to Schiemann WP, and by pilot funding from the Case Comprehensive Cancer Center’s Research Innovation Fund, which is supported by the Case Council and Friends of the Case Comprehensive Cancer Center to Schiemann WP.
Conflicts of interest
All authors declare that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2019.
 Journal of Cancer Metastasis and Treatment2019年5期
Journal of Cancer Metastasis and Treatment2019年5期
- Journal of Cancer Metastasis and Treatment的其它文章
- Operative treatment of metastatic breast cancer in the spine with regard to molecular phenotypes
- Chemotherapy-induced immunological breast cancer dormancy: a new function for old drugs?
- Autophagy in breast cancer metastatic dormancy: tumor suppressing or tumor promoting functions?
- Stem cells, immortality, and the evolution of metastatic properties in breast cancer: telomere maintenance mechanisms and metastatic evolution
- Training and evaluation of a knowledge-based model for automated treatment planning of multiple brain metastases
- ESR1 alterations and metastasis in estrogen receptor positive breast cancer