
焦酚对共培养铜绿微囊藻和雨生红球藻影响的初步研究
2019-07-19 07:46高云霓付琴琴张方李晨露武静董静
生态环境学报 2019年6期
高云霓,付琴琴,张方,李晨露,武静,董静
河南师范大学水产学院,河南 新乡 453007
藻类是水生态系统中重要的初级生产者,是水生态毒理学研究中最为常用的受试生物之一(Fu et al.,2017),也被广泛用于化感抑藻活性物质筛选和抑藻机理研究(Gao et al.,2017)。通过测试待测组分或物质对特定藻株的影响程度,评价其毒性或活性强弱。水华鱼腥藻(Anabaena flos-aquae)、水华微囊藻(Microcystis flos-aquae)和铜绿微囊藻(Microcystis aeruginosa)等蓝藻和羊角月牙藻(Selenastrum capricornutun)、四尾栅藻(Scenedesmus quadricauda)、斜生栅藻(Scenedesmus obliqnus)、普通小球藻(Chlorella vulgaris)、蛋白核小球藻(Chlorella pyrenoidosa)等绿藻被用来研究有机锡和拟除虫菊酯农药的生物毒性(Ma,2005)。产毒和非产毒铜绿微囊藻、四尾栅藻和湖泊微拟球藻(Nanno-chloropsis limnetica)被用来比较沉水植物水剑叶(Stratiotes aloides)分泌物化感抑藻活性(Mulderij et al.,2005)。这种经典抑藻测试方法的主要特点之一就是以单株藻为目标,构建纯培养体系研究目标藻株在个体、细胞、分子或代谢层面上的响应水平,方法简单、成熟、易标准化,因此在活性组分或物质的筛选研究中具有明显的优势。
然而,自然水体中藻类常常相伴而生,不同藻类之间相互作用。铜绿微囊藻通过释放亚油酸抑制普通小球藻生长(Song et al.,2017),却可促进水华束丝藻(Aphanizomenon flos-aquae)、水华鱼腥藻的生长(张婷等,2006)。普通小球藻能明显抑制羊角月牙藻(Pseudokirchneriella subcapitata)生长(Fergola et al.,2007);可控条件下产毒铜绿微囊藻和非产毒片状微囊藻与单针藻(Monoraphidium convolutum)、尖状栅藻(Scenedesmus acuminatus)之间可相互抑制对方生长(Bittencourt-Oliveira et al.,2014)。铜绿微囊藻和阿氏浮丝藻(Planktothrixa gardhii)共培养导致后者生长减慢,丝状体尺寸变小(Briand et al.,2018)。大量研究显示共存藻类之间存在着明显的相互作用,而这种相互作用是否会影响目标藻株对植物化感物质的响应水平,目前还不得而知。
在沉水植物对藻类的化感作用研究中,小二仙草科、金鱼藻科、眼子菜科、水鳖科和茨藻科的多种沉水植均有不同程度的化感抑藻作用,大量野外和室内实验证明穗花狐尾藻(Myriophyllum spicatum)的化感抑藻活性最强。而焦酚是从穗花狐尾藻甲醇提取物和培养液中分离、检测到的活性最高、研究最深入的典型化感物质之一(Nakai et al.,1996;Nakai et al.,1999)。已有大量研究报道焦酚对铜绿微囊藻等蓝藻的抑制机理,如高浓度焦酚对铜绿微囊藻氧化损伤、转录水平和毒素合成的影响(Wang et al.,2016);焦酚可诱导铜绿微囊藻产生细胞凋亡现象等(Lu et al.,2017)。焦酚等化感物质的持续低剂量释放,可能是自然水体水生植物化感抑藻的重要模式之一(Lu et al.,2014)。然而,这些研究都是基于纯培养抑藻测试体系,共存藻类的相互作用是否对焦酚的化感抑藻活性产生影响还不清楚。
混合共培养是最能模拟自然水体中藻类共存、相互作用的可控实验体系,现已被广泛用于研究藻类对营养等资源的竞争,不同光照、温度等环境条件下藻类的演替过程(Van De Waal et al.,2011)。混合共培养也在水生植物化感作用研究中得到初步运用。张庭廷等(2016)调查了水杨酸、肉桂酸与焦酚对铜绿微囊藻与蛋白核小球藻复合体系的影响,比较单一和复合酚酸对混合藻类的抑制作用。但此项研究没有关注藻类在纯培养和共培养体系中对化感物质的响应差异,不能提供直接的证据证明目标藻株对化感物质的响应是否受到共存藻类的影响以及影响程度。
本研究选择有害蓝藻铜绿微囊藻和经济绿藻雨生红球藻(Haematococcus pluvialis)为受试藻,比较混合共培养和纯培养体系中焦酚对两株藻生长和代谢的影响。其中,铜绿微囊藻是富营养化水体最为常见的水华藻种之一(Xiao et al.,2018),雨生红球藻是一种广泛分布的单细胞淡水绿藻,具有相对复杂的生活周期(Shah et al.,2016)。当受到环境胁迫时(例如高温、高光照、营养盐缺乏时),会形成较大的厚壁孢子,并失去鞭毛成为不动细胞,逐渐形成红色的虾青素(Kobayashi,1997)。本研究在焦酚暴露实验中同时设置纯培养和共培养体系,选取细胞密度等生长指标,结合微囊藻毒素释放量和雨生红球藻胞内虾青素质量浓度等代谢指标,比较分析共培养和纯培养条件下焦酚对两种藻的影响,为深入揭示水生植物化感抑藻作用生态机制,明确化感作用和化感物质的生态价值提供方法学参考。
1 材料与方法
1.1 材料和试剂
实验藻种蓝藻铜绿微囊藻(FACHB 905)和绿藻雨生红球藻(FACHB 797)均购自中国科学院水生生物研究所淡水藻种库。保种和实验期间均用无菌 BG11培养基在 2000 lx,12 h∶12 h光暗比,(25±1) ℃的条件下培养。每天早晚各手工振摇 1次。抑藻测试采用指数生长期藻细胞进行测试。
实验所选化感物质焦酚购自 Sigma-Aldrich公司(美国),Folin-Ciocalteu试剂购自北京索莱宝科技有限公司(中国),实验所用到的丙酮和二甲基亚砜均为分析纯。
1.2 实验设计
本实验设置3个培养体系,分别为铜绿微囊藻纯培养体系,雨生红球藻纯培养体系和铜绿微囊藻与雨生红球藻混合共培养体系,每个培养体系均设置空白对照组和焦酚处理组,每组3个平行。实验在250 mL锥形瓶中进行,每瓶藻液100 mL,各培养体系中藻细胞起始光密度值OD680均为0.03,其中铜绿微囊藻在纯培养和共培养体系中的起始细胞密度分别为 5.20×105cells·mL-1和 3.70×105cells·mL-1,雨生红球藻在纯培养和共培养体系中的起 始 细 胞 密 度 分 别 为 6.20×103cells·mL-1和2.91×103cells·mL-1。处理组添加焦酚到各培养体系后的最终浓度均为5.0 mg·L-1。实验进行3 d,分别在实验起始和结束时取样,测定藻细胞数、OD680、铜绿微囊藻胞外藻毒素质量浓度、雨生红球藻虾青素细胞配额(单个细胞内虾青素产量)和包括焦酚在内的总酚质量浓度。
1.3 测试指标及方法
细胞光密度测定及细胞计数:根据紫外-可见光谱扫描结果,发现680 nm处铜绿微囊藻和雨生红球藻均有最大吸收峰,因此所有样品均测定OD680。取0.1 mL 1%鲁哥氏液固定样品,于藻细胞计数框中,沉淀一定时间,光学显微镜下计数细胞数。由于雨生红球藻细胞直径是铜绿微囊藻的 10倍,共培养体系中两种藻在显微镜下容易区分,可分别计数。根据比生长速率计算公式计算基于细胞数和OD680的日生长速率(Xiao et al.,2017)。
胞外铜绿微囊藻毒素测定:采用中国科学院水生生物研究所微囊藻毒素 ELISA检测试剂盒测定胞外微囊藻毒素质量浓度(常孟阳等,2019)。5 mL藻液于10000 r·min-1下离心10 min,上清液直接用于胞外毒素测定。
胞内虾青素质量浓度测定:参照陈晓飞等(2007)的方法,修订如下:通过抽滤将样品中的藻细胞截留在0.45 µm纤维膜上,然后,将滤膜浸入二甲基亚砜(DMSO)中溶解,然后加入3 g石英砂,45-50 ℃水浴30 min,每隔10 min涡旋振荡15 s(共计3次)。3000 r·min-1下离心5 min使细胞物质沉淀,将上清液转入容量瓶中,再往离心管中加入丙酮,涡旋振荡30 s,继续离心,合并上清液,丙酮抽提至少3次,直至上清液基本无色。用丙酮定容至25 mL,混匀,取5-7 mL,再次离心后,上清液在474 nm波长下测定光密度。
总酚质量浓度测定:取 3 mL藻液于 8000 r·min-1下离心10 min,取2 mL离心后的上清液于试管中,添加1 mL Folin-Ciocalteu试剂,混匀,室温下放置5 min,然后添加1 mL 10%碳酸钠,混匀。25 ℃培养2 h后,测定765 nm波长下的光密度,结果以没食子酸的质量浓度表示(Singleton et al.,1999;Hilt et al.,2006)。
1.4 数据处理
运用Microsoft Excel 2016分析实验数据并作图,运用SPSS 22.0软件中的独立样本t检验和单因素方差分析检验不同处理之间的差异显著性(P<0.05)。
2 结果和分析
2.1 共培养条件下铜绿微囊藻和雨生红球藻的生长
通过观察纯培养和共培养体系中铜绿微囊藻和雨生红球藻3 d内的细胞数变化,发现两种藻在共培养与纯培养体系中的生长速率无显著差异(图1,P>0.05),初步说明两种藻在本实验条件下可以在同一环境中共同生长。但两种藻自身的生长速率存在显著差异(P<0.05),纯培养和共培养条件下,铜绿微囊藻的平均日生长速率分别为0.28和0.27,雨生红球藻的则为0.43和0.47。
2.2 共培养条件下焦酚对铜绿微囊藻和雨生红球藻生长的影响
通过对两种藻分别计数,计算生长速率和抑制率,发现焦酚作用下,铜绿微囊藻在纯培养和共培养体系中的平均日生长速率分别降至0.01和0.02,以无焦酚添加处理为对照,计算得到两种培养体系下焦酚对铜绿微囊藻生长的抑制率分别为 96.82%和 93.18%(图 2),说明两种培养方式下,焦酚几乎完全抑制铜绿微囊藻的生长,但共培养组焦酚对铜绿微囊藻生长的抑制率显著低于纯培养组(P<0.05),初步说明铜绿微囊藻在与雨生红球藻共存时,焦酚对铜绿微囊藻的抑制作用受到一定程度限制。
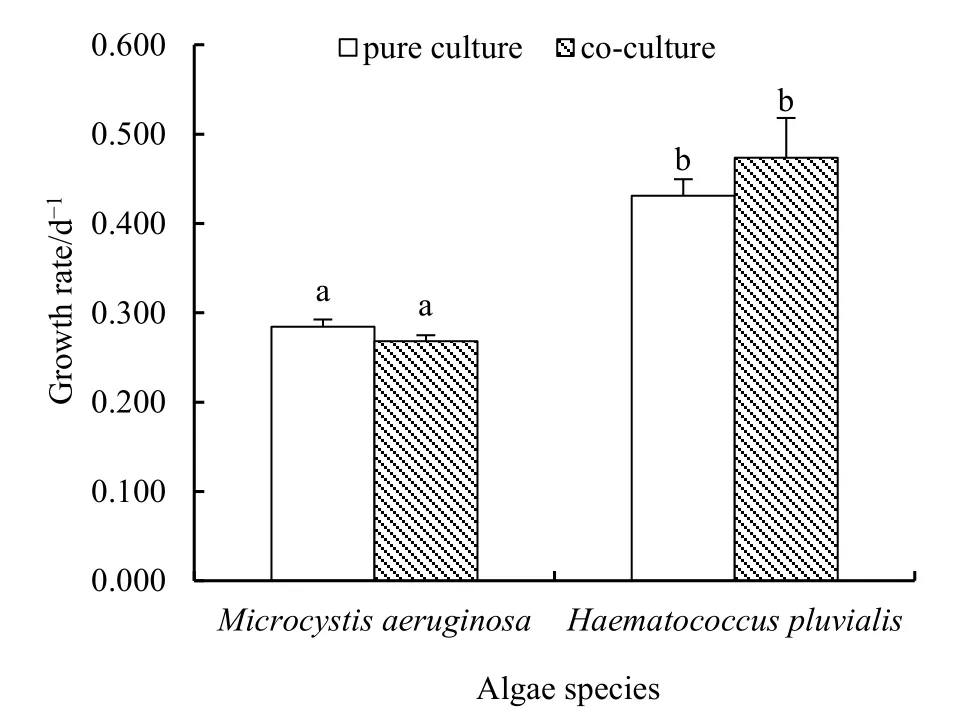
图1 铜绿微囊藻和雨生红球藻纯培养和共培养3 d的生长速率Fig. 1 Growth rate of 3 d pure culture and co-culture of Microcystisaeruginosa and Haematococcus pluvialis

图2 纯培养和共培养体系中焦酚对铜绿微囊藻和雨生红球藻的生长抑制率Fig. 2 Inhibition ratio of pyrogallol on growth of Microcystis aeruginosa and Haematococcus pluvialis in pure and co-culture
焦酚作用下,纯培养和共培养体系中雨生红球藻的平均生长速率分别为0.30和0.26,以各培养体系不加焦酚的空白组为对照,计算得到的抑制率分别为29.39%和45.40%(图2),说明两种培养方式下,焦酚均能显著抑制雨生红球藻的生长,且共培养体系中焦酚抑制率显著高于纯培养体系(P<0.05)。从生长指标来看,共存铜绿微囊藻增强了焦酚对雨生红球藻生长的抑制。
通过测定实验开始和结束时各处理中藻液OD680,计算得到铜绿微囊藻纯培养体系、雨生红球藻纯培养体系和共培养体系的生长速率依次为0.03、0.17和0.08。以各培养体系不加焦酚的空白组为对照,计算得到的抑制率分别为 92.20%、30.50%和73.77%(图3)。结果表明,3种培养体系中焦酚对藻类抑制率最高的是铜绿微囊藻纯培养体系,其次是共培养体系,最后是雨生红球藻纯培养体系。藻细胞计数和光密度测定结果均显示,无论是纯培养还是共培养条件下,焦酚对铜绿微囊藻的抑制均强于其对雨生红球藻的影响。
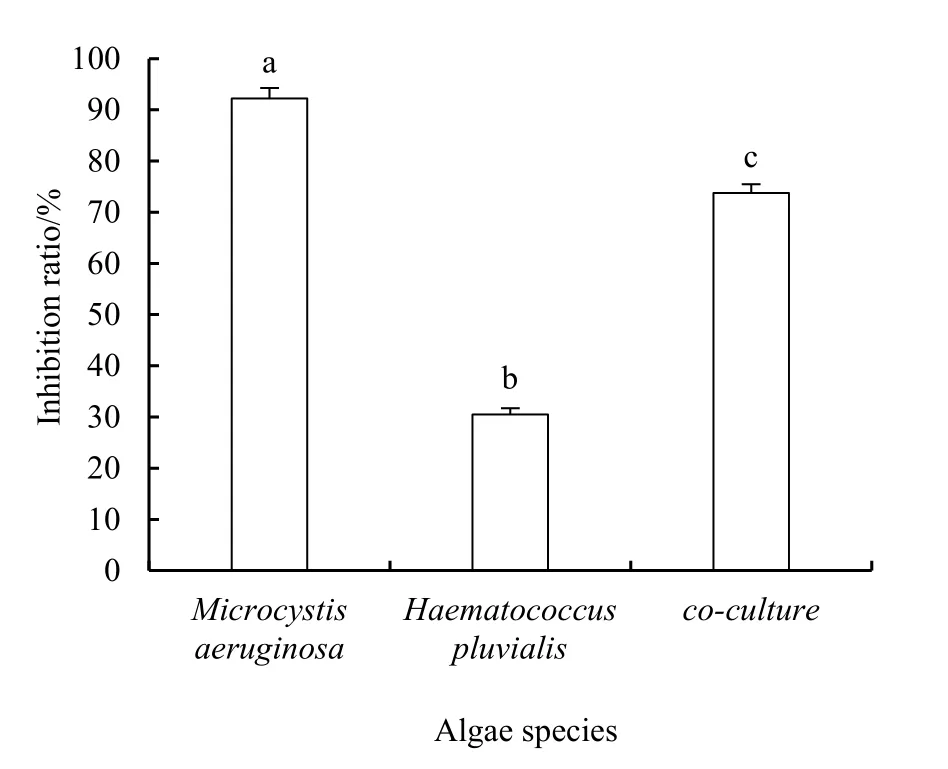
图3 基于藻液OD680计算的3个培养体系中焦酚的抑藻率Fig. 3 Algal inhibition ratio of pyrogallol calculated based on OD680 in the three culture systems
2.3 共培养条件下铜绿微囊藻和雨生红球藻在代谢水平上对焦酚的响应
图4 显示焦酚对纯培养和共培养体系中铜绿微囊藻毒素释放的影响。由图可知,共培养体系中铜绿微囊藻胞外毒素质量浓度均显著低于纯培养体系(P<0.05),这与各培养体系中接种的铜绿微囊藻初始细胞密度有关。实验开始和结束时,共培养体系中铜绿微囊藻细胞密度分别为纯培养体系中的71.02%和67.68%。添加焦酚后,纯培养和共培养体系中铜绿微囊藻释放到水中的藻毒素质量浓度均显著增加(P<0.05)。相比对应的空白组,胞外藻毒素质量浓度增加的幅度分别为105.60%和58.65%,共培养体系中胞外毒素增加幅度显著低于纯培养组(P<0.05),初步说明共存雨生红球藻可能减弱了焦酚对铜绿微囊藻的影响。

图4 焦酚处理下,纯培养和共培养体系中铜绿微囊藻胞外藻毒素浓度Fig. 4 Extracellular microcystin concentration in pure and co-culture solutions of Microcystis aeruginosa under pyrogallol treatment
不同处理下雨生红球藻单个细胞内虾青素产量,即虾青素细胞配额如图5所示。空白组和焦酚处理组,共培养体系中雨生红球藻虾青素细胞配额均显著高于纯培养体系(P<0.05)。无焦酚添加的空白组,共培养体系中胞内虾青素质量浓度比纯培养体系高63.60%,说明铜绿微囊藻的存在诱导雨生红球藻细胞内产生更多的虾青素。添加焦酚后两培养体系中雨生红球藻胞内虾青素质量浓度均显著增加(P<0.05)。与对应的空白组相比,添加焦酚的纯培养和共培养体系中胞内虾青素质量浓度分别增加63.61%和46.78%,共培养体系中胞内虾青素增加幅度显著低于纯培养组(P<0.05)。由于共存铜绿微囊藻和焦酚的复合作用,共培养体系中雨生红球藻产生的虾青素细胞配额显著高于对应的纯培养体系。
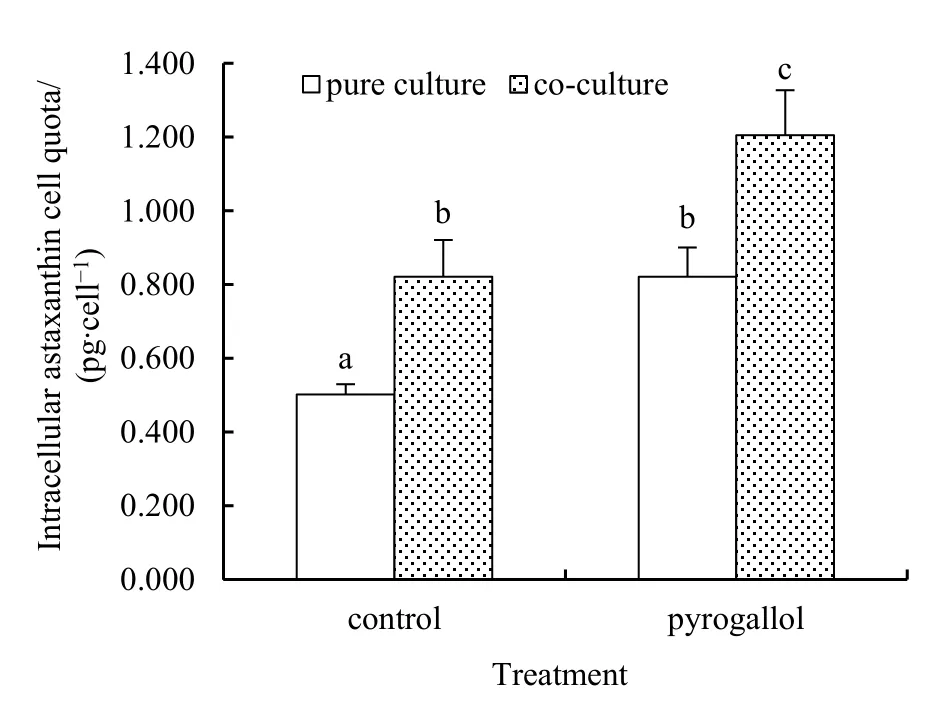
图5 焦酚处理下,纯培养和共培养体系中雨生红球藻胞内虾青素细胞配额Fig. 5 Intracellular astaxanthin cell quota in pure and co- culture of Haematococcus pluvialis under pyrogallol exposure
2.4 培养体系中总酚质量浓度变化

图6 实验开始和结束时纯培养和共培养体系中总酚质量浓度Fig. 6 Total phenolic mass concentration in pure and co-culture solutionsat the beginning and the end of the experiment
焦酚加入到纯培养和共培养体系3 d后,包括焦酚及其转化产物在内的总酚质量浓度均有显著降低(P<0.05)。实验开始时,铜绿微囊藻纯培养、雨生红球藻纯培养和两藻株共培养体系中的总酚质量浓度均值分别为 4.56、4.69、4.85 mg·L-1,组间没有显著性差异(P>0.05)。培养3 d后,3个实验组的总酚质量浓度分别为 1.24、0.84、1.10 mg·L-1,仅为起始浓度的27.28%、17.90%和22.68%(图6),雨生红球藻纯培养体系中总酚质量浓度显著低于微囊藻纯培养和共培养体系(P<0.05)。
3 讨论
光照、营养、温度、pH等非生物环境因子不可避免地影响自然水体中植物对藻类的化感作用(吴振斌等,2016)。相对于高温和强光照,穗花狐尾藻释放的8种化感物质对铜绿微囊藻生长和光合活性的抑制作用在低温低光照条件下更强(Nakai et al.,2014)。当pH为7.0时,氢醌暴露的4 d时间里均可完全抑制具刺链带藻(Desmodesmus armatus)和铜绿微囊藻的生长,而当pH为11.0时,氢醌的毒性在48 h内即完全消失(Bährs et al.,2014)。同时,处于同一水环境的其他藻类与目标藻类之间可能会为争夺有限的光照、营养、空间等资源而相互竞争(Liu et al.,2016),也有可能通过化学通讯和化感作用而相互促进或抑制(Song et al.,2017)。但共存藻类与目标藻类间的相互作用是否会影响它们对植物化感物质的响应目前还不清楚。本研究从生长和代谢水平证实,焦酚对铜绿微囊藻和雨生红球藻的抑制效果在纯培养和共培养体系中显著不同。铜绿微囊藻与雨生红球藻共存时,焦酚对铜绿微囊藻生长的抑制作用有一定程度的减弱,而对雨生红球藻生长的抑制程度显著增强。
微囊藻毒素是由淡水蓝藻(如铜绿微囊藻)产生的一类具有生物活性的环状七肽物质。微囊藻毒素性质稳定,煮沸后不失活,易溶于水,危害水生态系统,威胁人类健康(Roegner et al.,2014)。微囊藻毒素在蓝藻的正常生长阶段,释放到水体中的量很少,在藻细胞膜破损尤其是藻细胞死亡时,才会有大量的微囊藻毒素释放到细胞外进入水体(朱枭强,2016)。本研究两种培养体系中,焦酚处理均显著提高了胞外微囊藻毒素的质量浓度,但相较而言,纯培养体系中胞外毒素增加程度更大。由此可推测,焦酚处理下,共培养体系中破损或死亡细胞所占比例低于纯培养体系,这和细胞计数的结果一致。
大量研究显示,多种胁迫条件如营养缺乏、高盐度、高光、极端温度等都可以诱导雨生红球藻胞内虾青素的积累(Pelah et al.,2004;Giannelli et al.,2015)。尚敏敏等(2017)研究发现添加黄腐酸能促进雨生红球藻细胞内虾青素的积累。但关于化感物质诱导虾青素积累的研究还少有报道。本研究结果显示,共存铜绿微囊藻会促进雨生红球藻胞内产生更多的虾青素,在添加焦酚后,来自铜绿微囊藻和焦酚的共同胁迫导致雨生红球藻胞内虾青素产量显著增加,这与雨生红球藻单独存在时焦酚的胁迫效应明显不同。细胞计数结果也显示,共培养条件下焦酚对雨生红球藻生长的抑制作用显著增强。
Granéli et al.(2003)和 Fistarol et al.(2003)等的研究发现在氮、磷缺乏条件下,小定鞭金藻(Prymnesium parvum)滤液对威氏海链藻(Thalassiosira weissf l ogii)、微小原甲藻(Prorocentrum minimum)和波罗的海红胞藻(Rhodomonas cf. baltica)的生长有抑制作用,而在营养充足的条件下小定鞭金藻的滤液对上述藻不产生抑制作用。本研究中,尽管雨生红球藻的日生长速率显著高于铜绿微囊藻,每株藻在共培养和纯培养体系中的生长速率却没有显著差异,说明在本实验设定的共培养条件下,短时间内两株藻均能在BG 11培养液中正常生长。但焦酚暴露后,两株藻受到的抑制效应却与纯培养时显著不同。
刘碧云等(2008)通过纯培养抑藻测试体系研究发现焦酚对4种蓝藻和绿藻生长的抑制效果存在差异,本研究进一步证实,无论是纯培养还是共培养体系,焦酚对铜绿微囊藻的抑制效果均显著高于对雨生红球藻的影响,这可能与藻细胞结构不同有关。雨生红球藻厚壁孢子具有厚且致密的孢粉质细胞壁,当焦酚作用于细胞时,这层孢粉质细胞壁阻碍了化感物质进入细胞内部,从而降低了焦酚的抑藻效果(任晓丽等,2018)。
大多数酚类物质在pH>7时都易自动氧化,同时伴有自由基的产生(Bährs et al.,2014)。焦酚会快速自氧化为红培酚,随后会进一步转化为其他多酚,多酚氧化时毒性显著增强,因为氧化产生的自由基可以使DNA断裂,使低密度的脂蛋白和脂过氧化,导致藻细胞生理功能的失调,从而影响藻细胞的生长(Lu et al.,2016)。本研究检测了代表焦酚及其各级氧化产物总量的总酚质量浓度,发现3 d后总酚质量浓度为起始量的17.90%-27.28%。3个培养体系中,雨生红球藻纯培养体系中总酚质量浓度最低,其次是共培养体系。这与光密度所显示的焦酚对各培养体系的抑制率一致,即总酚质量浓度最低的雨生红球藻纯培养体系焦酚的抑制率也最低。由此推测,焦酚氧化产物质量浓度与焦酚的抑藻效果之间呈现明显的正相关关系。
本研究结果对植物化感抑藻作用机制研究和植物源控藻物质的应用均具有重要的指导意义。一方面,相对于以前以单株藻为目标的纯培养测试体系,构建两株或多株藻共培养体系,尽可能模拟自然水体真实情况,所揭示的规律和机制才具有较为广泛的生态学价值。另一方面,对于有望用于藻华控制和藻相调控的植物源化感抑藻物质而言,必须考虑到共存藻类的相互作用对抑藻效果潜在的影响,才能较为准确地确定化感物质控藻剂量和控藻方式,以取得接近预期的实践效果。因此,后续相关研究有必要更好地模拟自然水体中的实际情况,优化实验设置,延长实验时间,考虑共存藻株,结合化感物质的释放动态,设置暴露模式,结合室内和原位实验,更充分地揭示生物-非生物环境因子复合作用下植物化感抑藻作用机制,探讨植物化感作用在有害蓝藻控制和藻相调控优化中的应用方式。
4 结论
本研究设置了铜绿微囊藻纯培养、雨生红球藻纯培养和两者共培养体系,比较分析焦酚对两种藻生长和代谢的影响。结果显示,焦酚对铜绿微囊藻的抑制作用均显著强于其对雨生红球藻的影响。与纯培养相比,共培养体系中焦酚对铜绿微囊藻生长的抑制率减弱,微囊藻毒素释放量增加的幅度显著降低(P<0.05),而对雨生红球藻生长的抑制率增强(P<0.05),单个细胞内虾青素产量显著增加(P<0.05),表明两者共存减弱了焦酚对铜绿微囊藻的抑制效应,却增强了焦酚对雨生红球藻的影响。
猜你喜欢
中南药学(2022年8期)2022-11-19
可再生能源(2022年8期)2022-08-17
天然产物研究与开发(2021年10期)2021-11-03
河北地质大学学报(2020年2期)2020-06-04
江苏农业学报(2019年1期)2019-09-10
大自然探索(2019年2期)2019-03-01
现代园艺(2017年23期)2018-01-18
中南大学学报(自然科学版)(2016年2期)2017-01-19
杂草学报(2015年2期)2016-01-04
通化师范学院学报(2012年8期)2012-08-15
