
海拔对藏北高寒草地物种多样性和生物量的影响
2019-07-19 07:46吴红宝水宏伟胡国铮王学霞干珠扎布严俊何世丞谢文栋高清竹
生态环境学报 2019年6期
吴红宝,水宏伟, ,胡国铮,王学霞,干珠扎布*,严俊,何世丞,谢文栋,高清竹
1. 中国农业科学院农业环境与可持续发展研究所,北京 100081;
2. 安徽师范大学地理与旅游学院,安徽 芜湖 241000;3. 西藏自治区那曲市草原站,西藏 那曲 852000
草地生态系统作为陆地生态系统的重要组成部分(王长庭等,2004),具有维持生物多样性稳定、调节气候等生态服务功能和稳定生产力水平、保障畜牧业可持续发展等社会生产功能(郑晓翾等,2007;贾文雄等,2015)。研究表明,人类活动和气候变化可能导致生物多样性和生产力及其关系发生变化,也有可能导致生态系统结构与功能发生改变(Klein et al.,2004;Bellard et al.,2012;Ganjurjav et al.,2016;张中华等,2018)。物种多样性作为生物多样性最直接、最易观察和最主要的研究层次,是研究草地生态系统多样性、结构及功能的重要方面和重要指标,有助于维持草地生态系统的稳定和良性运转(Tilman,2000;白可喻等,2013;干珠扎布等,2015)。生物量是草地生态系统结构与功能的重要表现形式,是维持草地生态系统稳定发展的基础,对草地维护与管理及牧区畜牧业的可持续发展起到重要作用(Tilman et al.,1996;郑晓翾等,2007)。因此,草地生态系统物种多样性和生物量逐渐成为生态学最为关注的热点问题之一。
物种多样性沿海拔的变化规律是生物多样性研究的重要议题,海拔梯度涵盖了温度、湿度及太阳辐射等环境因素的综合影响,对物种多样性的垂直分布格局具有重要作用(Gaston,2000;Oommen et al.,2005)。就海拔梯度而言,国内外研究主要围绕草地物种多样性、生物量及物种多样性与生物量关系等方面展开(Wilson et al.,1990;段敏杰等,2011;刘哲等,2015)。贺金生等(1997)指出,物种多样性随海拔梯度的变化主要有 5种表现形式,即随海拔上升而降低、升高、先升高后降低、先降低后升高及无规律性变化。栗文瀚等(2017)在综述前人研究基础上指出,草地地上生物量与海拔间主要表现为单峰分布、正相关和负相关3种关系。在物种多样性与生产力相互关系中,Waide et al.(1999)则指出30%呈单峰关系,26%呈线性正相关,12%呈线性负相关及32%呈无规律变化。在目前已有的研究中,不同海拔梯度物种多样性、生物量及其二者关系以单峰分布、正相关和负相关最为普遍(冯建孟等,2009;刘海丰等,2012;王国宏,2002;刘哲等,2015),且这些分布关系在青藏高原尤为突出。在青藏高原,王长庭等(2004)发现地上生物量随海拔的升高而逐渐降低,物种多样性在中等海拔梯度时最高,物种多样性与地上生物量表现出单峰分布格局;段敏杰等(2011)也认为该地区物种多样性在海拔上呈单峰分布,但由于受到人类居住环境和放牧等人类活动的干扰,植物群落盖度和地上生物量随海拔增加则呈现出先增加后降低的趋势;牛钰杰等(2017)认为由于水分和热量的差异,即使是同一山体,阳坡和阴坡物种丰富度也明显不同,即阳坡物种丰富度随海拔的增加而增加,阴坡在海拔上呈单峰分布格局。因研究尺度及特定生态环境因子的差异(Davidson et al.,2007;杨瑞红等,2016;牛钰杰等,2017),物种多样性及其生物量与海拔梯度的关系至今还没有统一的定论。因此,如何全面地认识物种多样性及其生物量与海拔梯度的关系需要科研工作者进一步根据研究尺度结合各自区域尺度下海拔梯度土壤因子、气候因子及人为干扰等多重因素进行深入探究。
藏北高原位于青藏高原的腹地,平均海拔4500 m 以上,被称为“世界屋脊”和地球“第三极”。其独特的地理位置和复杂的自然环境,对研究高海拔草地植被群落物种多样性和生物量分布格局具有重要意义。目前,针对研究区物种多样性和生物量沿海拔梯度变化的研究较多(曲广鹏等,2015;牛钰杰等,2017;栗文瀚等,2017),但物种组成沿海拔梯度的变化及物种变化对物种多样性的响应机制还比较缺乏。研究区素以低温、干燥及大风等特点著称,微小的海拔变化将会引起上述环境因子的改变,进而导致草地生态系统产生较大差异。然而,前人的研究选取的山地海拔高度多集中于3840-4635 m(王长庭等,2004;栗文瀚等,2017),忽略了高寒草地植被分布在更高海拔条件下微气候及土壤水热条件等差异对物种组成、物种多样性及生物量的影响。因此,本研究以藏北高原罗玛镇境内4600-4800 m高寒草地为研究对象,通过样方调查与取样的方法对研究区植被物种组成、盖度、高度及地上生物量等进行调查,探讨分析高寒草地植物物种组成、物种多样性及生物量对海拔变化的响应,揭示不同海拔梯度上高寒草地资源现状,为研究区高寒草地生态系统植物群落物种演替研究与保护、草地资源调查、多样性研究及生态系统管理与评价提供科学依据,对明确高寒草地生态系统功能与结构和维持生态系统稳定性具有重要的科学意义。
1 材料与方法
1.1 研究区概况
本试验样地位于藏北高原罗玛镇境内(31°17′N,91°50′E)。藏北高原(那曲市)位于“世界屋脊”、“亚洲水塔”——青藏高原的腹地,平均海拔4500 m以上,辖区面积为4.46×105km2,其中草地面积为 4.21×105km2,约占 94.4%(高清竹等,2006),是西藏自治区主要畜牧业生产基地和中国重要的生态屏障区。罗玛镇位于唐古拉山和念青唐古拉山之间,境内贡曲河、俄曲河、那曲河及江曲河等河流纵横交错,辖区面积4146 km2,其中草地面积1290 km2。研究区属高原亚寒带季风半湿润气候区,年平均气温为0.3 ℃,生长季(5-9月)平均气温为8 ℃,年降水量为461.8 mm,生长季降水量为403.2 mm(2005-2014年)(干珠扎布,2017)。采样期间,研究区空气平均温度为18.0 ℃,土壤平均温度和湿度分别为13.8 ℃和34.7%。空气温度和土壤温度随海拔上升呈降低的趋势,土壤湿度则随海拔的增加呈上升的趋势(图1)。

图1 试验区不同海拔空气温度、土壤温度及湿度Fig. 1 Air temperature, soil temperature and soil moisture of different elevation in experimental area
研究区植物主要为高山嵩草(Kobresia pygmaea)、矮生嵩草(Kobresia humilis)、青藏苔草(Carex moorcroftii)、草地早熟禾(Poa pratensis)、紫花针茅(Stipa purpurea)、紫花棘豆(Oxytropissubfalcata)及委陵菜(Potentilla chinensis)等。土壤类型主要为高寒草地土。
1.2 试验设计
于2018年8月,对罗玛镇境内高山草地不同海拔植物物种多样性及生物量空间分布格局进行调查分析。沿一条垂直样线由山脚至山顶,每隔50 m设置1个海拔梯度,共设5个海拔梯度,分别为4600、4650、4700、4750、4800 m。在每一海拔梯度上布设10 m×10 m采样区,随机选取3个0.5 m×0.5 m的试验样方。对每个样方进行植物群落调查,按植物种类将样方内植物划分为三类群,分别为禾本科、莎草科和杂类草。记录样方内所有植物物种数、高度及盖度等。将样方内植物分种用剪刀齐地面剪下装进信封袋,然后将鲜草置于105 ℃烘箱中杀青0.5 h,以70 ℃恒温烘干至恒质量,称其干质量,计算地上生物量。采用便携式土壤多参数速测仪测定(TDR150,美国,Spectrum)各海拔梯度各样方内5 cm土层土壤温度及湿度,同时手持该仪器测定各海拔梯度1 m高空空气温度,每个海拔梯度3个重复。
1.3 数据计算
物种多样性采用物种丰富度(S)、Shannon-Wiener指数(H)及E. Pielou均匀度指数(E)表示,计算公式如下:
重要值:

物种丰富度:

Shannon-Wiener指数:

E. Pielou均匀度指数:

式中,IV为重要值;C为相对盖度;h为相对高度;Pi为相对重要值;H为Shannon-Wiener多样性指数;E为E. Pielou均匀度指数;S为物种丰富度;N为样方内出现的植物物种数。
1.4 数据处理
运用统计软件Excel 2013对各海拔采样数据进行均值和标准误差处理和绘图。运用 IBM SPSS Statistics 17.0单因素方差分析(One Way ANOVA)及最小显著差异法(LSD)对各海拔采集的数据进行差异显著性检验,利用曲线估计(Curve estimation)对地上生物量与物种丰富度、Shannon-Wiener指数、E. Pielou均匀度指数进行曲线拟合。
2 结果与分析
2.1 物种组成及其重要值沿海拔梯度的变化
海拔对藏北高寒草地植物物种及组成类群的重要值具有重要影响(表1和图2)。随着海拔的增加,植物物种组成差异较大,优势种由高山嵩草(Kobresia pygmaea)+青藏苔草(Carex moorcroftii)逐渐转变为矮生嵩草(Kobresia humilis)+珠芽蓼(Polygonum viviparum),其中旱生的禾本科植物逐渐被独一味(Lamiophlomis rotata)、垫状点地梅(Androsace tapete)和珠芽蓼(Polygonum viviparum)等喜湿植物取代。
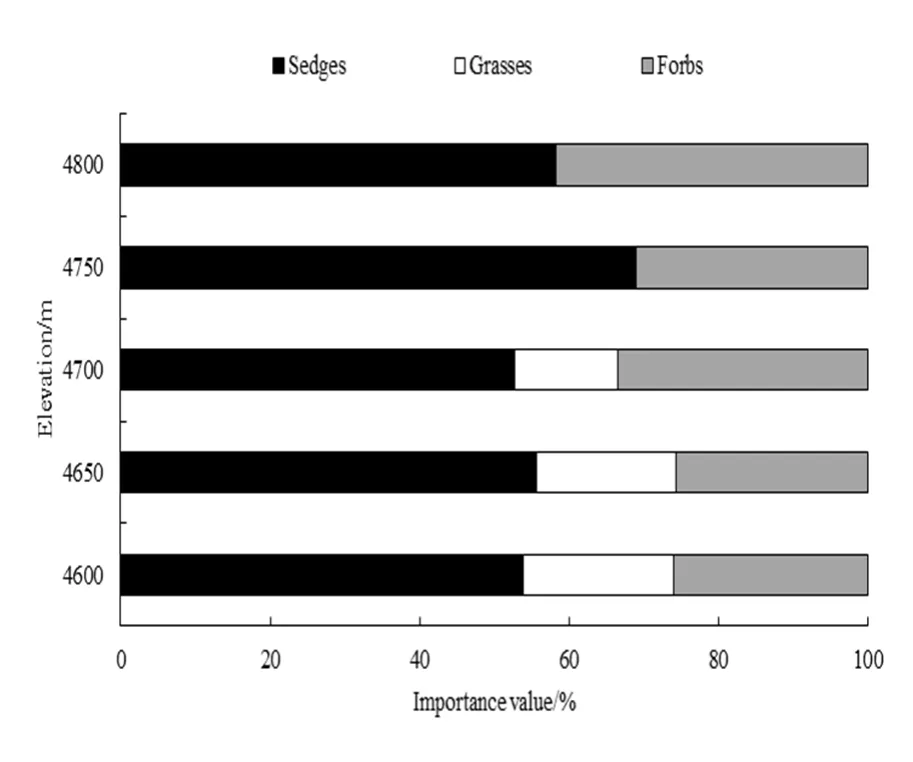
图2 不同植物类群重要值沿海拔变化Fig. 2 Important values of different plant groups vary along the elevation
随着海升高,各海拔物种数依次为15、17、13、10、11种。4600 m和4650 m莎草类、禾草类及杂草类物种比例均较为接近,分别为53.8%、20.2%、26.0%和 55.6%、18.7%、25.7%。不同海拔梯度上物种组成存在一定的差异,如在 4650 m出现了4600 m未出现的线叶嵩草(Kobresia capillifolia)、高山唐松草(Thalictrum alpinum)、山莓草(Sibbaldia procumbens)、蒲公英(Taraxacum mongolicum)和藏黄芪(Astragalus tibetanus)等,而垂穗披碱草(Elymus nutans)、多裂委陵菜(Potentilla multifida)、肉果草(Lancea tibetica)、星状雪兔子(Saussurea stella)和藜(Chenopodium album)等物种消失。至海拔 4700 m,物种重要值主要表现为禾草类物种比例减少,杂草类物种比例增加。具体表现为禾草类如仅羊茅(Festuca ovina)这单一物种存在,而杂草类则增加了独一味和高山紫菀(Aster alpinus),杂草类物种比例较4650 m增加了7.8%。至海拔4750 m和4800 m,禾本科植物物种消失,莎草科植物物种间变化差异明显,如矮生嵩草由低海拔的6.8%显著增加到30.8%。
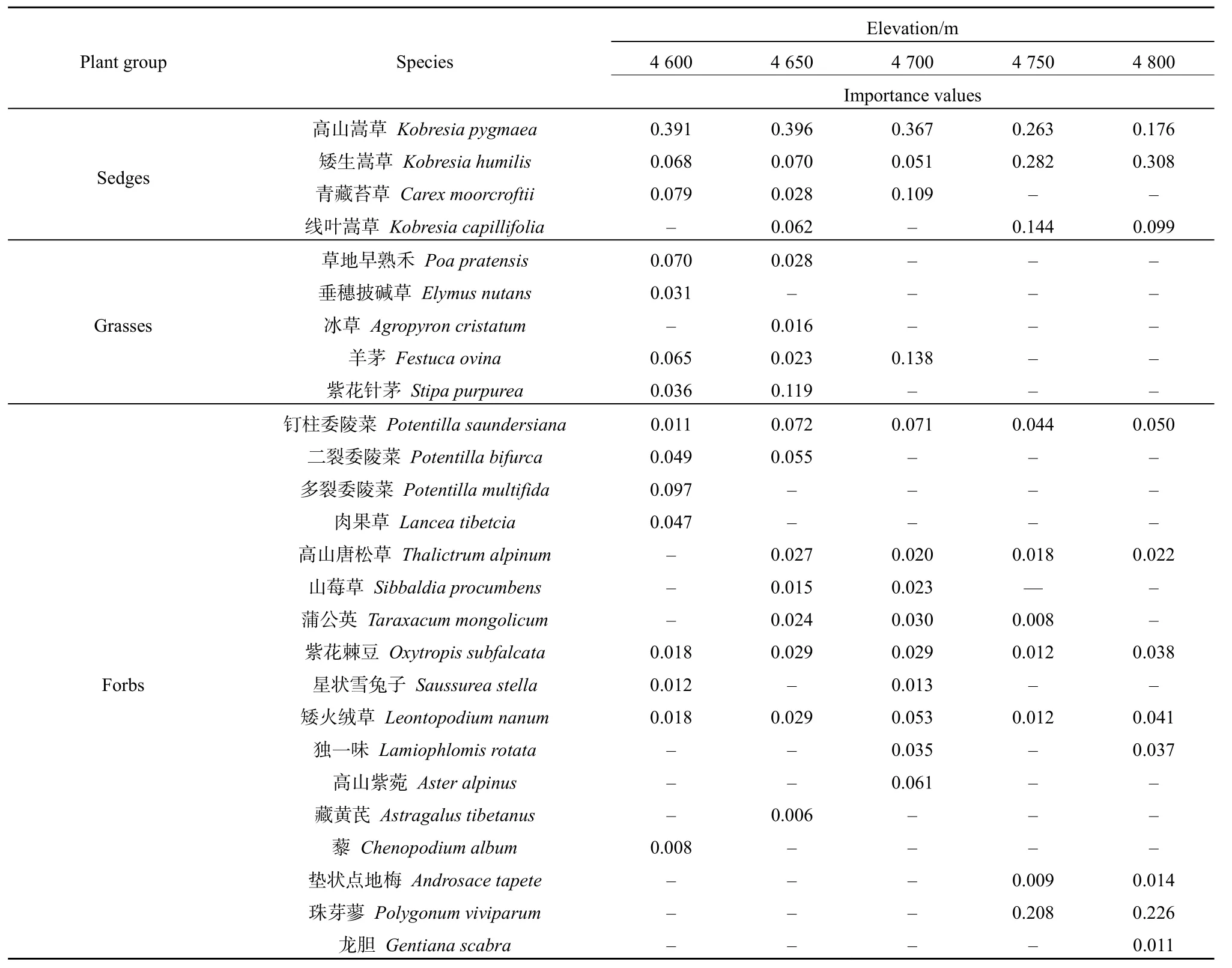
表1 不同海拔高寒草地植物物种及组成类群的重要值Table 1 Plant species and importance values of group composition in alpine grassland at different elevations
2.2 物种多样性沿海拔梯度的变化
高海拔段物种丰富度和 Shannon-Wiener指数显著低于中海拔段(P<0.05),而各海拔梯度间 E.Pielou均匀度指数无显著差异(P>0.05)(图 3)。由图 3可知,物种丰富度的变化趋势基本上与Shannon-Wiener多样性指数相吻合。海拔 4600-4650 m,物种丰富度随海拔增加而增加;至4750 m,则随海拔的升高而显著降低(P<0.05),至4800 m 则再次增加,但差异不显著(P>0.05)。Shannon-Wiener指数在海拔4600-4700 m间随海拔的增加而升高,至4750 m则显著降低(P<0.05)。E. Pielou均匀度指数与物种丰富度和Shannon-Wiener指数具有相反的变化趋势,即当海拔从4600 m升至4650 m时E. Pielou均匀度指数降低,而海拔升高至4750 m时,E. Pielou均匀度指数不断增加,至4800 m时再次下降,各海拔间差异不显著(P>0.05)。
物种丰富度和 Shannon-Wiener多样性指数与海拔间趋势模拟均呈负二次函数关系,E. Pielou均匀度指数则呈正二次函数关系,可以看出,物种丰富度和Shannon-Wiener指数随海拔的变化呈现“单峰”分布格局,即海拔高度增加,物种丰富度和Shannon-Wiener指数呈先增加后降低的趋势;而E.Pielou均匀度指数则呈U型曲线分布格局,即先降低后增加。
2.3 地上生物量与物种多样性的关系
植物盖度和地上生物量在海拔上表现出相同的变化趋势,即随海拔的增加而降低(图4)。4600-4800 m,植物盖度和地上生物量的变化范围分别在63.7%-89.0%和41.0-64.7 g·m-2之间。其中在4600-4700 m,植物盖度均在 80%以上,海拔间差异不显著(P>0.05)。至4750 m,植物总盖度快速降低,降幅达 25.5%,与低海拔间差异显著(P<0.05)。与之不同的是,地上生物量在 4700 m显著降低(P<0.05),较4600 m和4650 m分别降低29.1%和26.9%。4700-4800 m,地上生物量趋于稳定,较4700 m差异不显著(P>0.05)。

图3 物种多样性沿海拔变化Fig. 3 Variation of the species diversity at different elevationsThe lowercase letters in the picture represent significant differences(P<0.05); n=5; Mean±SE. The same below

图4 不同海拔梯度植被盖度及生物量特征Fig. 4 Characteristics of the vegetation coverage and biomass atdifferent elevations
在海拔梯度上,藏北高寒草地地上生物量与土壤温度呈指数增长,与土壤湿度呈指数降低。物种丰富度和 Shannon-Wiener指数与地上生物量间的趋势模拟均呈负二次函数关系,E. Pielou均匀度指数与地上生物量关系不显著(图5)。分析得出,物种丰富度和 Shannon-Wiener指数随地上生物量的变化呈单峰分布,即随地上生物量的增加,物种丰富度和Shannon-Wiener指数先增加后降低。当地上生物量分别为 54.0 g·m-2和 54.7 g·m-2时,物种丰富度和Shannon-Wiener指数分别达到最大值,说明当地上生物量处于中等水平时,物种丰富度和Shannon-Wiener指数最大。
3 讨论
植物物种多样性及生物量的空间分布格局是各种生态因子作用的综合反映,海拔作为众多因子中最为关键的因素之一,其变化通常会影响温度、湿度及太阳辐射等的再分配,进而影响山地植物的分布格局(Gaston,2000;Li et al.,2011;牛钰杰等,2017;张树斌等,2018)。

图5 地上生物量与物种多样性及土壤温度的关系Fig. 5 Relationship between above-ground biomass and species diversity and soil temperature at different elevations
在青藏高原,物种多样性沿海拔变化通常以先增加后降低的单峰分布最为常见(王长庭等,2004;Sa et al.,2012;曲广鹏等,2015),本研究结果也符合这一分布特征,即物种丰富度和Shannon-Wiener指数随海拔变化呈单峰分布。不同海拔梯度土壤水热间的组合关系控制着植物的生理活动,低海拔地区土壤温度高而湿度低(图1),占据39.1%生态位的高山嵩草加强了对土壤水分的竞争,抑制了喜湿植物的生长与繁殖;高海拔地区土壤湿度高而温度低(图1),高湿度的土壤环境不利于中旱生禾本科植物的生长。同时高海拔段风速急、土壤蒸发强烈及昼夜温差大(唐志尧等,2004;冯建孟等,2006)等恶劣条件只能维持适应力极强的物种生存;在中等海拔段,土壤温度和水分适宜,适宜的水热配比允许更多的物种共存,物种多样性达到峰值。因此,在低海拔段,物种多样性主要受土壤湿度的限制,而高海拔区由于土壤水分充足,主要受土壤温度限制,形成了中等海拔梯度物种多样性较高的分布格局(牛钰杰等,2017;苏闯等,2018)。
众多研究表明,植物生物量沿海拔的空间分布格局以单峰、V字型及单调线性降低为主(王长庭等,2004;段敏杰等,2011;刘哲等,2015)。造成以上分布的差异,部分研究结果支持人为干扰是其重要因素(栗文瀚等,2017),尤其是牲畜的采食与践踏(干珠扎布等,2013);但也有研究证实是山地水热组合条件及土壤养分差异导致的(贾文雄等,2015)。在本研究中,高寒草地植物地上生物量随海拔的增加而降低,支持单调线性降低的分布格局,说明温度是决定地上生物量沿海拔分布最关键的因子。通常情况下,在低海拔段,植物生长所需的热量充足而水分短缺,高海拔段则水分充足,热量可能成为其生长的限制因素(王国宏,2002),温度和水分的变化会引起物种组成的改变(Wu et al.,2011;Ganjurjav et al.,2016;Hopping et al.,2018)。本研究发现,不同海拔梯度间山地植物物种组成存在较大差异,在土壤温度高、湿度低的低海拔区,适宜高山嵩草和中旱生的禾本科植物生长。马维玲等(2010)认为海拔升高引起的温度降低导致了禾本科植物获取养分资源的能力下降,因此,随着海拔的升高,禾本科植物物种比例也随之降低(栗文瀚等,2017)。而温度低、湿度大的高海拔段适合矮生嵩草、垫状点地梅及珠芽蓼等耐寒喜湿植物的生长,抑制了中旱生禾本科植物的生长,中旱生禾本科植物逐渐被这些耐寒喜湿植物所取代。干珠扎布等(2015)研究表明灌溉引起土壤水分的增加导致了高寒草地禾草科植物物种比例降低和垫状点地梅等喜湿杂草类的增加。王长庭等(2007)也证实冷湿的土壤环境更有利于矮生嵩草的生长发育。
植物物种多样性与地上生物量的关系是生态系统发生与发展的基础,一直是生态学研究的热点。目前,众多研究表明,二者间的相互关系主要表现为5种关系,即线性正相关、线性负相关、单峰曲线分布(中间高度膨胀)、U型曲线分布及不相关(Waide et al.,1999;Tilman et al.,2001;Adler et al.,2011;干珠扎布等,2015)。造成以上物种多样性和地上生物量关系复杂多样的原因可能是空间尺度、资源利用生态位补偿效应及人为干扰等因素的结果(Davidson et al.,2007;Weigelt et al.,2008;段敏杰等,2011)。尽管有研究支持高物种多样性引起高地上生物量的假设(Tilman et al.,2001),但在青藏高原,物种多样性与地上生物量的关系却以单峰分布格局为主(王长庭等,2004;Sa et al.,2012)。本研究中物种丰富度和Shannon-Wiener指数与地上生物量呈负二次函数关系,即单峰曲线分布格局,这也说明当地上生物量处于中等水平时,物种丰富度和Shannon-Wiener指数最大。
4 结论
通过对不同海拔高寒草甸物种多样性及地上生物量的测定,发现研究区植物物种丰富度和Shannon-Wiener指数随海拔变化呈单峰分布,地上生物量随海拔升高而单调递减,物种丰富度和Shannon-Wiener指数与地上生物量呈单峰曲线分布格局。对不同海拔高寒草地物种组成、物种多样性及地上生物量的研究,有助于明确海拔梯度上草地的空间结构与资源现状,对高寒草地生态系统的管理与保护及生态系统功能与结构的稳定具有重要的科学意义。
猜你喜欢
散文诗(2021年22期)2022-01-12
河南畜牧兽医(2020年21期)2020-01-10
环球时报(2019-05-23)2019-05-23
质谱学报(2018年5期)2018-10-11
北京航空航天大学学报(2017年3期)2017-11-23
中国药业(2014年17期)2014-05-26
郑州大学学报(工学版)(2014年6期)2014-03-01
中成药(2014年10期)2014-02-28
小小说月刊·下半月(2012年12期)2012-05-14
