
84K杨4CL3/4CL5基因克隆及生物信息学分析
2019-07-19 10:30:42孙晓莎赵曦阳曲冠证
植物研究 2019年4期
孙晓莎 王 遂 赵曦阳 曲冠证
(林木遗传育种国家重点实验室,东北林业大学,哈尔滨 150040)
木质素由三种不同木质素单体通过不同比例组成,分别为香豆醇(p-coumaryl alcohol,H型),松柏醇(Coniferyl alcohol,G型),芥子醇(Sinapyl alcohol,S型)。不同的植物品种中木质素单体的比例大致是相近的,例如在杨树中,S型和G型木质素单体比值大约为2∶1[1~2]。不同单体的化学特性有所不同,例如G型单体C5位置形成C-C键,该键在化学降解作用具有高度稳定性,而S型木质素单体的醚键在化学降解过程中稳定性则弱一些,因此在Kraft制浆工艺中,G型单体比S型单体更难去除[3]。木质素单体的合成由三个代谢途径共同完成。第一个代谢途径:莽草酸代谢途径(Shikimate pathway);第二个代谢途径:苯丙烷类代谢途径(General phenylpropanoid pathway);第三个代谢途径:木质素合成的特异途径(General phenylpropanoid pathway)。在代谢途径中有几种重要的酶如:4-香豆酰-CoA连接酶(4-coumarate:coenzyme A ligase,4CL,EC6.2.1.12)、苯丙氨酸脱氨酶(Phenylalanine Ammonia-Lyase,PAL)及肉桂酸-4羟化酶(Cinnamate 4-Hydroxylase,C4H)等,其中4CL酶主要催化4-香豆酸、咖啡酸、阿魏酸、5-羟基阿魏酸和芥子酸等,生成相应的酰基CoA酯,如香豆酰-、咖啡酰-、阿魏酰-、5-羟基阿魏酰-和芥子酰-CoA等。这些CoA将进入木质素合成特殊途径,经由肉桂酰-CoA还原酶(Cinnamoyl-CoA reductase,CCR)和肉桂醇脱氢酶(Cinnamyl alcohol dehydrogenase,CAD)的作用下,最终形成木质素单体-香豆醇、松柏醇和芥子醇[4~5]。在1999年,首次成功地通过降低白杨(Populustremuloides)4CL基因表达促使羟基肉桂酸酯在木质素合成途径中无法被活化,从而降低了45%的木质素含量,并使纤维素的含量增加了15%。此后,降低4CL表达的策略在其它树种中也得到了验证[6]。吴锦程等人在枇杷中发现低温(4℃)会增加果肉和木质素含量,该过程主要是由于PAL酶升高导致[7]。因此推断降低4CL酶活可能会保持果肉含量不变而降低木质素合成含量。木质素填充于纤维素构架中,可起到增强植物体的机械强度、有利于根部输导组织的水分运输及抵抗不良外界环境的侵袭。另一方面木质素是制浆造纸业的不利因素,除去纤维素中的木质素,需要消耗大量能源,提高了造纸成本。4CL作为重要的木质素合成关键基因之一,研究4CL有助于了解木质素合成,进而提高木材抗压性。此外抑制4CL基因表达可以减少木质素积累,降低造纸成本。
杨树是我国重要的造林和用材树种,也是研究林木生理和基因工程的模式植物。84K杨由韩国育种学家以银白杨(Populusalba)为母本,腺毛杨(P.glandulosa)为父本杂交培育出来的品种,为雄性无性系,1984年引入我国。84K杨抗逆性强,具有较强的抗旱、抗寒、抗病、抗风能力,以及根系发达、造林成活率高等优良特征[8];同时还具有材质好,苗期生长快,树体高大直立等优点。此外,84K杨为雄性,无“飞絮”,是一种不会对环境造成污染的优良品种。84K杨具备分布广、适应性强、生长速度快、遗传转化率高等特点,是杨树基因工程研究的绝佳材料。因此,本研究以84K杨为材料,采用同源克隆的方法获得了Pag4CL3/4CL5基因并进行生物系信息学分析,为杨树木质素合成的遗传改良奠定理论基础。
1 材料与方法
1.1 试验材料
84K杨(Populusalba×P.glandulosa)由东北林业大学林木遗传育种国家重点实验室保存。
1.2 方法
1.2.1Pag4CL3/4CL5基因克隆
将84K杨野生型植株草炭土培养28天,培养室温度为22±2℃,光照强度为200~300 μmol·m-2·s-1,光周期为16 h/8 h。相对湿度为40%。选取长势一致的植株分别取叶、茎、根以及顶芽四个部分用MiniBEST Plant RNA Extraction Kit试剂盒提取RNA,并利用TaKaRa反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser反转录为cDNA。根据已公布的毛果杨(PopulustrichocarpaTorr.&A.Gray)基因组信息,设计Pag4CL3基因和Pag4CL5基因特异性引物见表1,以反转录的cDNA为模板,通过PCR方法,克隆出Pag4CL3/4CL5两个基因。PCR反应程序:94℃ 5 min;94℃ 30 s,58℃ 30 s,72℃ 1 min 45 s,30个循环;最后72℃ 7 min。用Biospin Gel Extraction Kit试剂盒回收PCR产物,回收产物与pEASY连接载体,转化大肠杆菌感受态细胞(Trans1-T1),涂于LB固体培养基(含50 mg·mL-1卡那霉素),37℃培养12 h后,随机挑取单克隆,用液体LB培养基(含卡那霉素)进行37℃振荡培养,6 h后,用全式金的Easy Pure Plasmid MiniPrep Kit试剂盒提取质粒DNA并进行菌液PCR检测,将阳性单克隆菌液送往哈尔滨擎科生物公司测序。
表1Pag4CL3/4CL5基因克隆及定量引物
Table 1Pag4CL3/4CL5 gene cloning and quantitative primers

引物名称Primer name引物序列Primer sequence4CL3-FTCTAGATCTGCATCTTTAGCCCGCAATG4CL3-RGGTACCTGTTCAGTAACGTCTTCAGTTA4CL5-FTCTAGACCCTCTGCAACTTTAGCCCACAATG4CL5-RGGTACCATATGCCCTTTACAATCTTCATTTA4CL3-RT-FGGCAGACGAAAATGAAGCGCCT4CL3-RT-RACCGACTCTCAGCCCGCAGA4CL5-RT-FACACAGGCTGATGATGATGATATGCC4CL5-RT-RGCAGCACCAGCTCTCAACCCAActin-FTACAACGAGCTTCGTGTTGCActin-RACAAGGAAAGGACAGCCTGA
1.2.2Pag4CL3/4CL5基因的生物信息学分析
通过在线软件DNAMAN和BioEdit进行氨基酸序列比对分析;利用在线软件MEME(http://meme.Nbcr.net/meme/cgi-bin/meme.cgi)进行4CL蛋白的保守结构域分析;利用在线网站ProtParam(http://web.expasy.org/protparam/)进行蛋白的理化性质分析;利用SOPM(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.ht ml)预测4CL蛋白的二级结构;利用SWISS-MODEL(https://www.swissmodel.expasy.org/)预测4CL蛋白的三级结构及预测4CL基因编码蛋白的功能;利用ProtScale软件(http://web.Expasy.org/protscale/)进行疏水性进行分析;利用TMpred软件(https://embnet.vital-it.ch/cgi-bin/TMPRED_form_parser)进行4CL蛋白序列跨膜区的预测。利用MEGA5软件构建系统进化树。
1.2.3Pag4CL3/4CL5基因在84K杨不同组织中的表达模式分析
利用Primer5.0软件设计定量引物,由哈尔滨市擎科生物公司合成。qRT-PCR通过SYBR Premix Ex TaqTMⅡ荧光定量试剂盒,以不同组织部位cDNA为模板,并重复3次,进行定量检测。反应体系为:SYBR Mixture 10 μL、模板2 μL、水6.4 μL、上游引物0.8 μL、下游引物0.8 μL;反应程序为:95℃ 30 s;95℃ 5 s,60℃ 35 s,40个循环;95℃ 15 s;60℃ 1 min;95℃ 15 s。数据分析采用ΔΔCt法,得到Pag4CL3/4CL5基因在不同组织部位的相对表达量。
2 结果与分析
2.1 Pag4CL3/4CL5基因全长克隆及其编码的氨基酸序列分析
根据毛果杨4CL基因序列设计引物,PCR扩增,克隆得到两个84K杨4CL基因(Pag4CL3和Pag4CL5)(图1),Pag4CL3基因cDS全长1 623 bp,编码一个由540个氨基酸残基组成的蛋白序列(图2);Pag4CL5基因cDS全长1 632 bp,编码一个由543个氨基酸残基组成的蛋白序列(图3)。GenBank登录号分别为MK183033(Pag4CL3)和MK183034(Pag4CL5)。对84K杨4CL基因编码的蛋白的理化性质进行预测分析,结果显示Pag4CL3蛋白的氨基酸中共含有70个带负电荷氨基酸残基,58个带正电荷氨基酸残基,蛋白的分子质量为58 987.28 Da,等电点为5.52,不稳定系数为36.50,该值小于40,为稳定蛋白;脂肪族指数为98.44,蛋白疏水性平均值为-0.027,为亲水性蛋白。Pag4CL5蛋白的氨基酸中共含有67个带负电荷氨基酸残基,59个带正电荷氨基酸残基,蛋白的分子质量为59 268.57 Da,等电点为5.72,不稳定系数为35.66,为稳定蛋白;脂肪族指数为96.10,蛋白疏水性平均值为-0.060,为亲水性蛋白。

图1 Pag4CL3/4CL5基因PCR产物 M.DL5000 marker;1.Pag4CL3基因的PCR产物;2.Pag4CL5基因的PCR产物Fig.1 PCR product of Pag4CL3/4CL5 gene M.DL5000 marker;1.PCR product of Pag4CL3 gene;2.PCR product of Pag4CL5 gene
2.2 84K杨4CL蛋白结构域及二、三级结构分析

图2 Pag4CL3基因的cDNA序列及其编码的氨基酸序列Fig.2 cDNA sequence and its deduced amino acid sequence of Pag4CL3
我们与毛果杨、青杨及毛白杨三种杨树的4CL蛋白进行序列比对,结果发现Pag4CL3与Pt4CL3蛋白相似性高达97%;Pag4CL5与Pt4CL5蛋白相似性也达到97%。此外,这四种杨树4CL蛋白均包含SSGTTGLPKGV和GEICIRG两个基序(图4),这两个基序构成了4CL家族标志性的保守结构域。SSGTTGLPKGV被认为是4CL催化反应中结合AMP保守积序[9];GEICIRG催化活动中心[10]。

图4 Pag4CL3、Pag4CL5与Pt4CL3、Pt4CL5、PtC4CL、Pc4CL蛋白结构域分析Fig.4 Structural domain analysis of Pag4CL3,Pag4CL5 with Pt4CL3,Pt4CL5,PtC4CL and Pc4CL
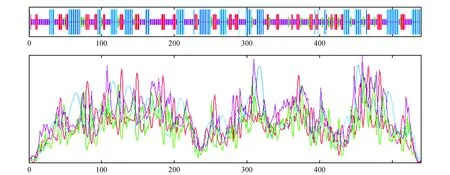
图5 Pag4CL3蛋白二级结构预测与分析Fig.5 Secondary structure prediction and analysis of Pag4CL3 protein
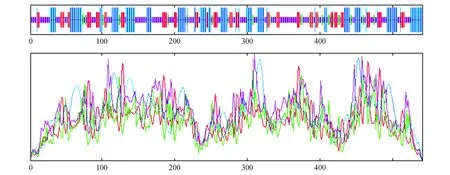
图6 Pag4CL5蛋白二级结构预测与分析Fig.6 Secondary structure prediction and analysis of Pag4CL5 protein
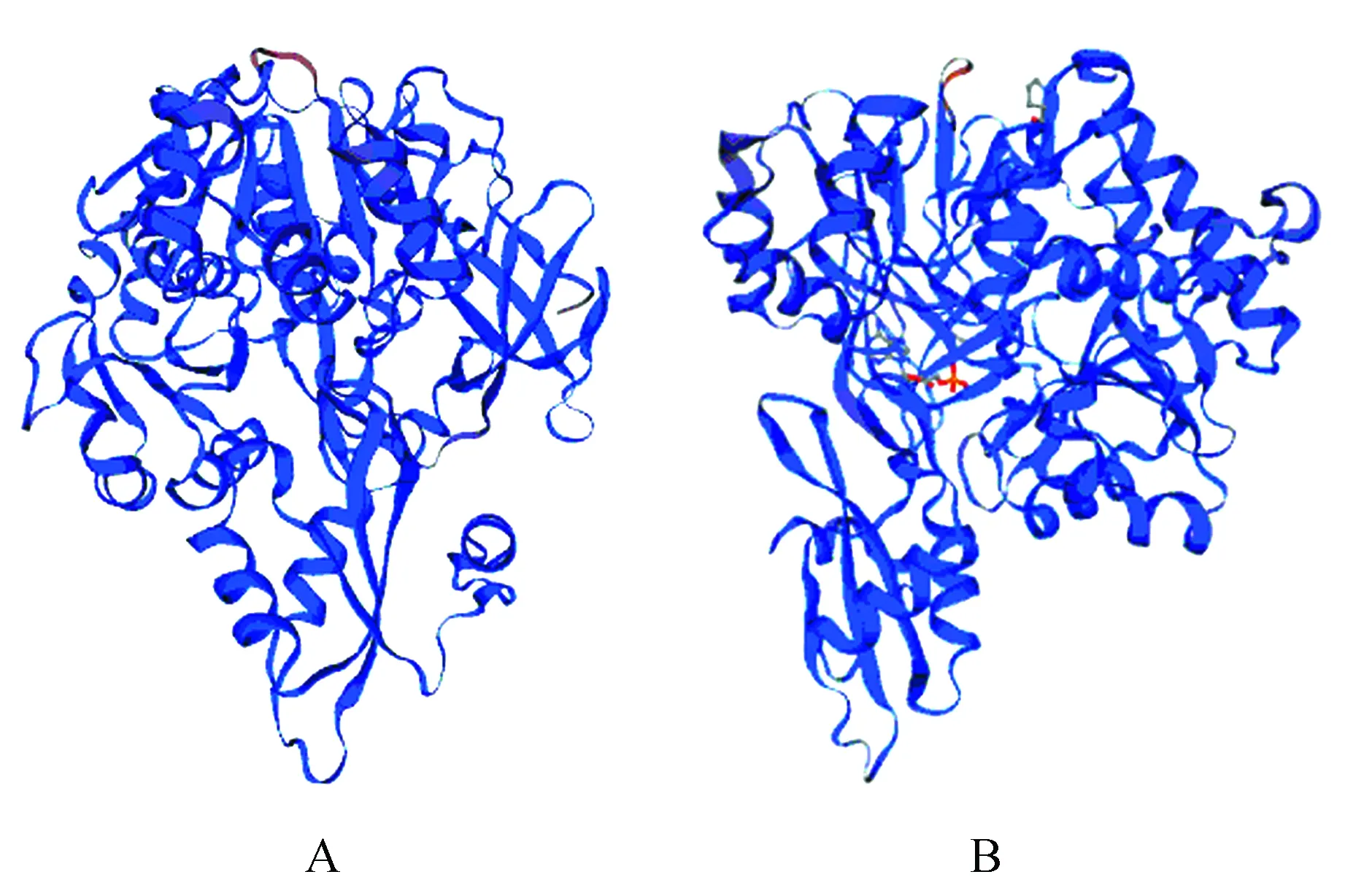
图7 4CL蛋白三级结构预测 A.Pag4CL3编码蛋白三级结构;B.Pag4CL5编码蛋白三级结构Fig.7 The encoding protein tertiary structure prediction of 4CL A.Pag4CL3 encoded protein tertiary structure; B.Pag4CL5 encoded protein tertiary structure
应用SOPMA在线软件对蛋白的二级结构进行预测和分析。84K杨4CL3蛋白的二级结构由32.04%的α螺旋(α helix)、20.0%的延伸链(extended strand)、7.41%的β转角(β turn)和40.56%无规则卷曲(random coil)组成(图5);84K杨4CL5蛋白的二级结构由32.60%的α螺旋(α helix)、18.60%的延伸链(extended strand)、7.00%的β转角(β turn)和41.80%无规则卷曲(random coil)组成(图6)。
运用在线软件SWISS-MODEL得到蛋白质三级结构的预测结果:4CL蛋白质的多肽链在各种二级结构的基础上再进一步盘曲或折叠形成具有一定规律的三维空间结构。Pag4CL3蛋白三维结构由一个PDB号为3a9u.1.A的结构为模板建立,3a9u.1.A属于4-coumarate—CoA ligase,序列的相似性为0.6,范围为9~536 aa,平均值为0.99(图7A);Pag4CL5蛋白三维结构由一个PDB号为3a9v.1.A的结构为模板建立,3a9v.1.A属于4-coumarate—CoA ligase,序列的相似性为0.57,范围为9~536 aa,平均值为0.98(图7B)。
2.3 4CL基因系统发育树的构建
在NCBI网站下载4CL蛋白序列包括:毛果杨4CL3(Populustrichocarpa,XP_002297699)、毛果杨4CL5(Populustrichocarpa,XP_002304825)毛白杨4CL(Populustomentosa,AAL56850)、青杨4CL(Populuscathayana,AJA91785)、白花泡桐4CL(Paulowniafortunei,ACL31667)、赤桉4CL(Eucalyptuscamaldulensis,ACX68559)、拟南芥4CL1(Arabidopsisthaliana,AAQ86588)、拟南芥4CL2(Arabidopsisthaliana,AAQ86587)、拟南芥4CL3(Arabidopsisthaliana,AAQ86589)、拟南芥4CL5(Arabidopsisthaliana,AAQ86591)、葡萄4CL(Wine grape,XP_002274994)、白桑4CL(Morusalbavar.multicaulis,AHL83551)、陆地棉4CL(Gossypiumhirsutum,ADU15550)、山葡萄4CL(Vitisamurensis,AWY10751)、珊瑚菜4CL(Glehnialittoralis,ATG32161)、枣4CL(Ziziphusjujuba,ALM23512)、石榴4CL(Punicagranatum,AFU90743)、苹果4CL(Malushybridcultivar,AFG30056)、大麻槿4CL(Hibiscuscannabinus,ADK24217)、辣椒4CL(Capsicum annuum,AAG43823)以及芝麻4CL(Sesamumindicum,ALA55541),并构建进化树(图8)。依照进化树的推断得知Pag4CL3/4CL5与毛果杨4CL、毛白杨4CL及青杨4CL被分为同一个分支上,说明它们在进化的过程中亲缘关系最为密切。
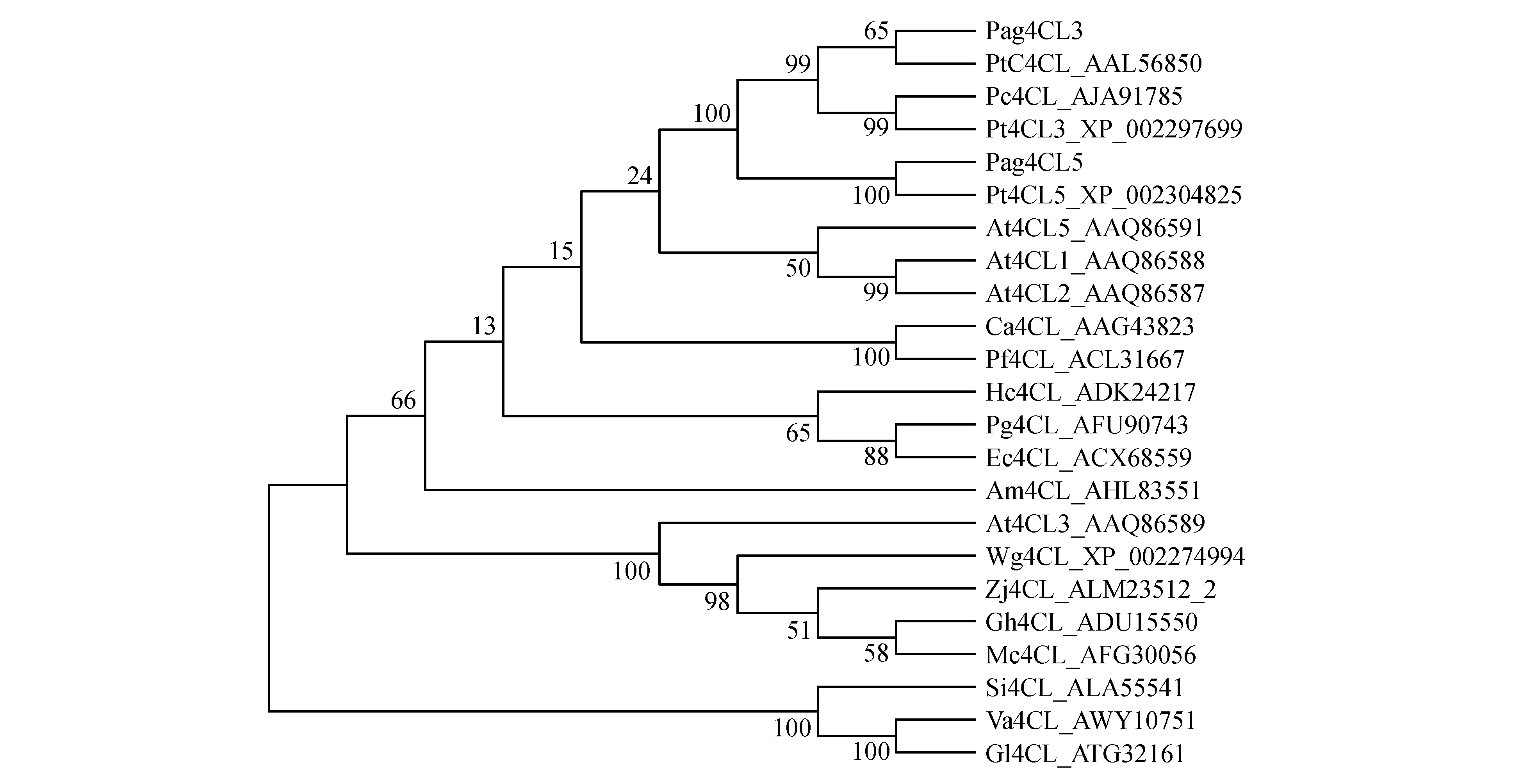
图8 4CL蛋白家族系统进化树Fig.8 The clustering analysis of 4CL protein family

图9 4CL保守结构域分析Fig.9 The conservative structure domain analysis of 4CL
利用在线软件MEME对Pag4CL3/4CL5进行蛋白序列分析,结果发现Pag4CL3/4CL5蛋白与其它杨树中4CL蛋白一致,基序种类、排序方式及基序位置都非常保守。这一结果与系统进化树相吻合(图9)。
2.4 84K杨Pag4CL3/4CL5功能预测
利用GenScript在线预测Pag4CL3/4CL5蛋白序列亚细胞定位,结果发现(表2),Pag4CL3/4CL5蛋白可能是一种膜蛋白,主要在质膜上行使功能。
表2 Pag4CL3/4CL5蛋白亚细胞定位预测
Table 2 Subcellular localization prediction of Pag4CL3/4CL5 protein
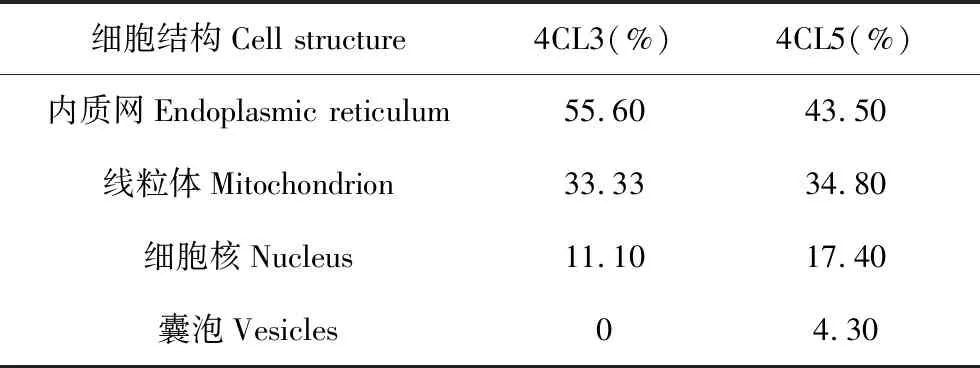
细胞结构Cell structure4CL3(%)4CL5(%)内质网Endoplasmic reticulum55.6043.50线粒体Mitochondrion33.3334.80细胞核Nucleus11.1017.40囊泡Vesicles04.30
利用ProtScale软件对蛋白疏水性进行分析,结果表明:在Pag4CL3蛋白的81~85区域、92~96区域、230~238区域、243~246区域、248~258区域、266~268区域、278~286区域、342~348区域、454~457区域、480~483区域以及515~518区域具有很强的疏水性,其中疏水性最强的在233,高达2.9(图10);在Pag4CL5蛋白的81~85区域、92~96区域、231~238区域、243~246区域、248~260区域、266~268区域、278~289区域、342~348区域、465~467区域、480~483区域以及517~518区域具有很强的疏水性,其中疏水性最强的在233,高达2.9(图11)。
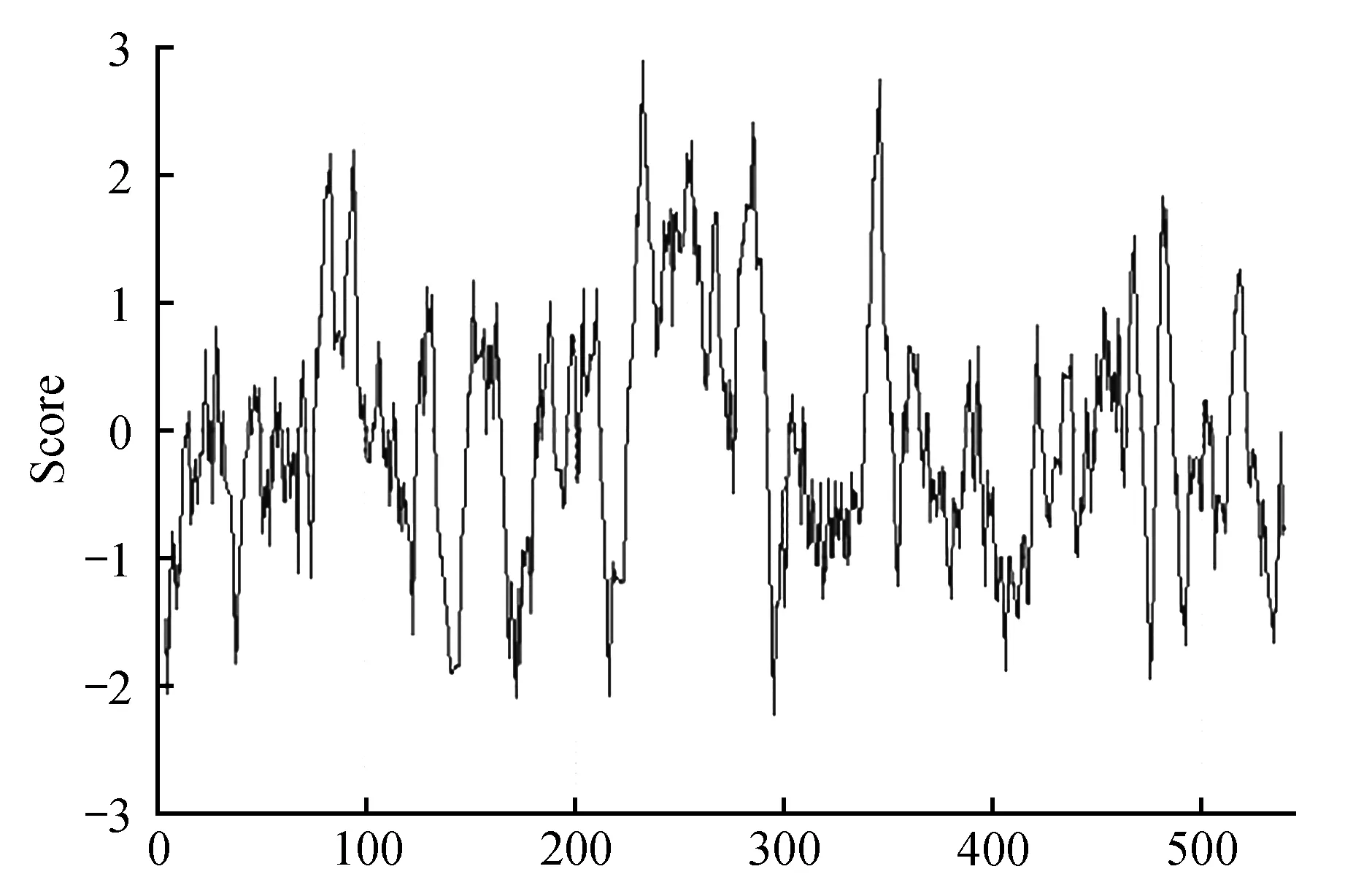
图10 Pag4CL3疏水性分析Fig.10 Hydrophobicity analysis of Pag4CL3
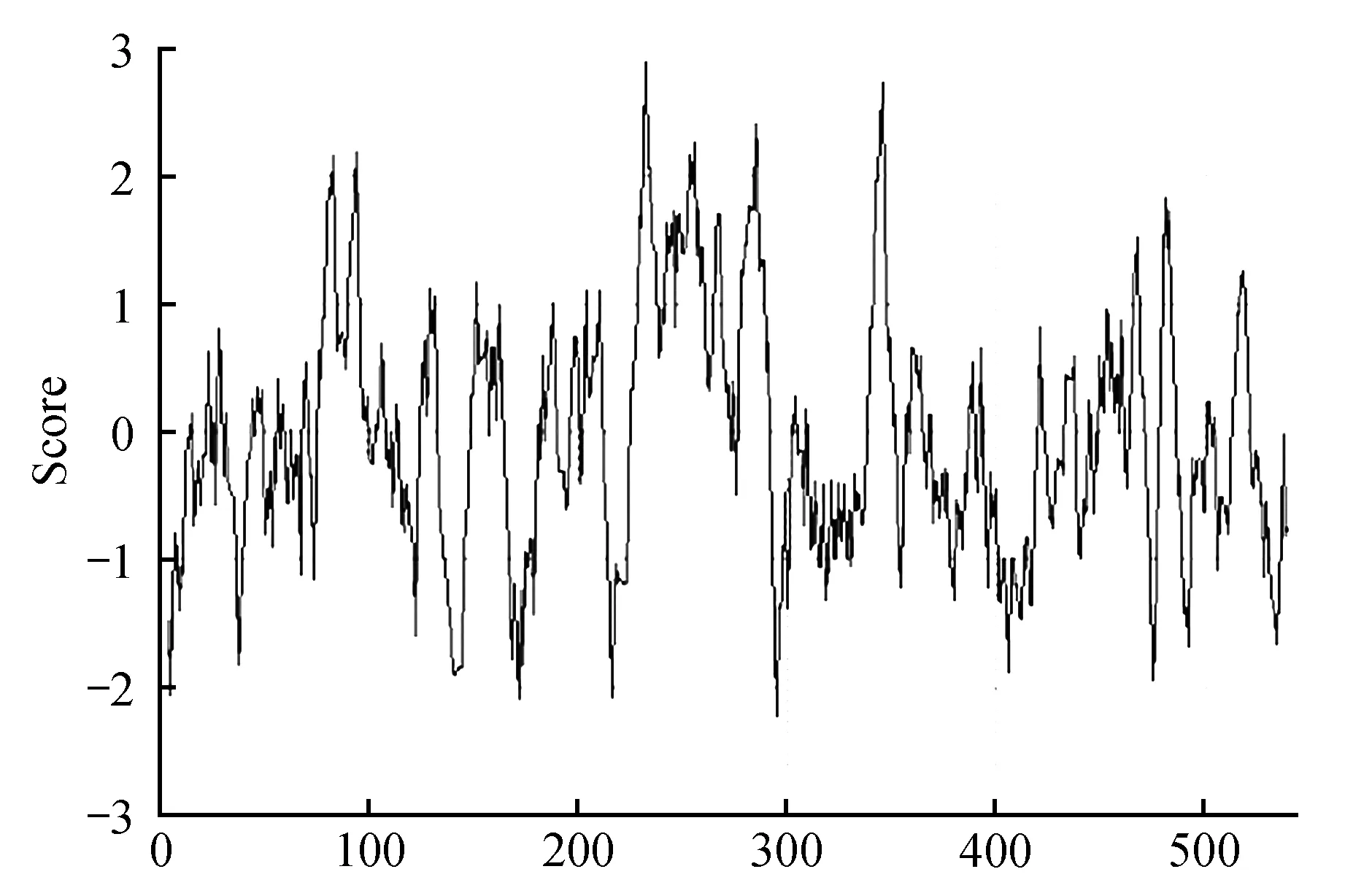
图11 Pag4CL5疏水性分析Fig.11 Hydrophobicity analysis of Pag4CL5

图12 Pag4CL3蛋白跨膜结构域预测Fig.12 Transmembrane domain prediction of Pag4CL3 protein
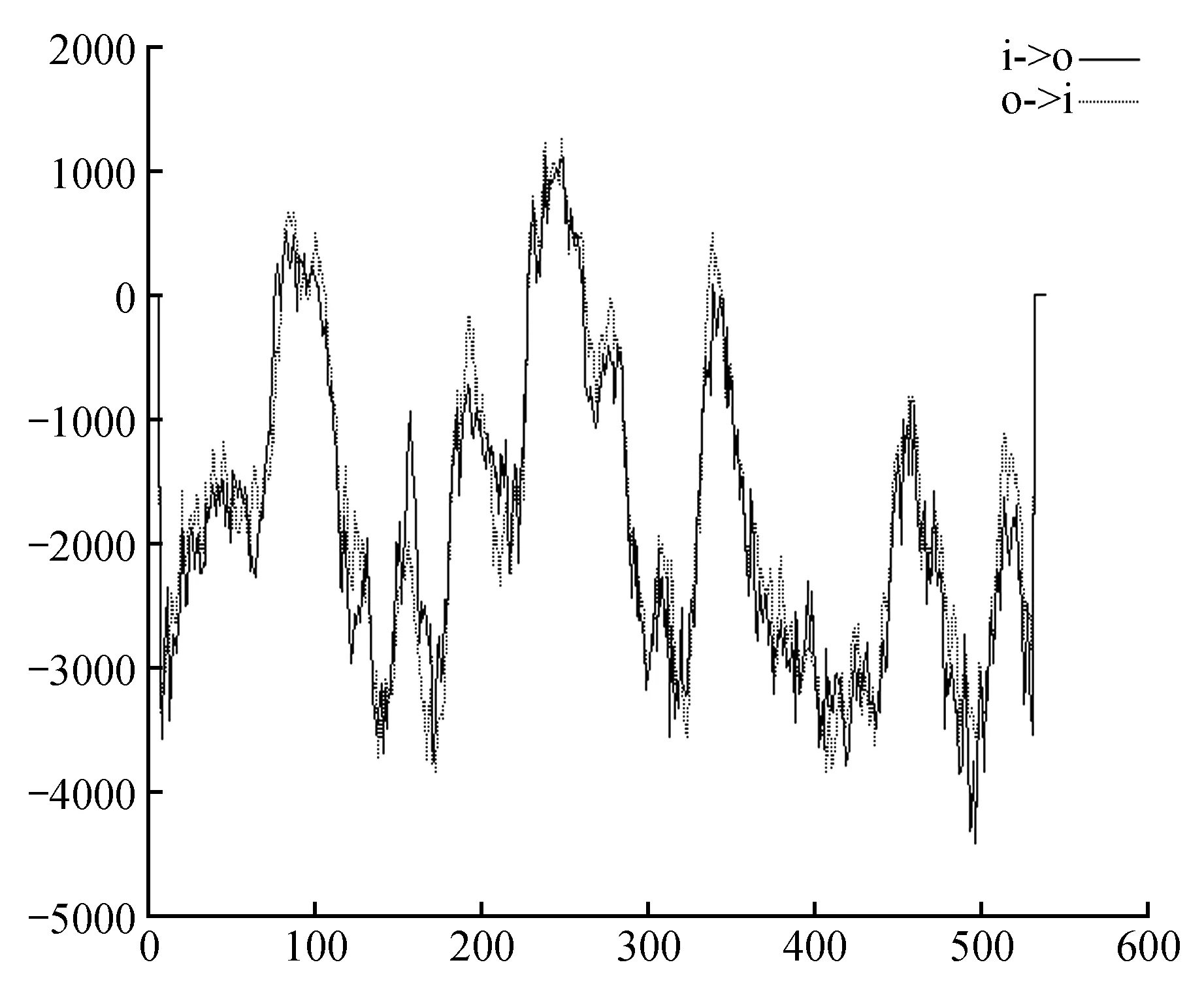
图13 Pag4CL5蛋白跨膜结构域预测Fig.13 Transmembrane domain prediction of Pag4CL5 protein

图14 Pag4CL3基因在不同组织中的表达分析Fig.14 Relative expression levels of Pag4CL3 in different organs
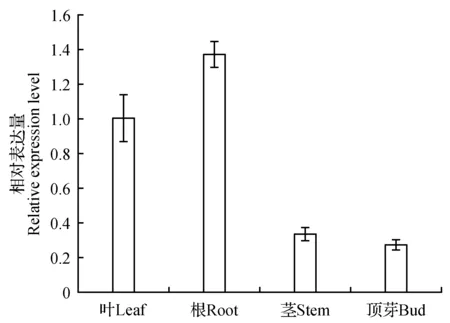
图15 Pag4CL5基因在不同组织中的表达分析Fig.15 Relative expression levels of Pag4CL5 in different organs
利用TMpred软件对蛋白跨膜结构域进行预测,预测结果为Pag4CL3蛋白从内到外共有1个跨膜区域在229~254 aa。从外到内有2个跨膜区域,在73~96 aa以及330~350 aa(图12);Pag4CL5蛋白从内到外共有1个跨膜区域,241~259 aa,从外到内有2个跨膜区域,在73~96 aa以及330~350 aa(图13)。
2.5 Pag4CL3/4CL5基因组织表达特异性分析
为了研究Pag4CL3/4CL5基因在不同组织部位中的表达特异性,本实验采用实时荧光定量PCR方法,分析其在不同部位的表达差异。如图14~15所示:Pag4CL3主要在叶及茎中表达,根和顶芽中表达量较低;与叶组织相比,顶芽中4CL3转录本低10倍左右。Pag4CL5主要在叶及根中表达,茎和顶芽叶中表达量较低;与根组织相比,顶芽中4CL5转录本低6.5倍左右。综上所述,两种同源基因的表达具有一定的组织部位差异性。
3 讨论
4CL是苯丙烷类代谢途径中的关键酶,也是木质素合成中重要的限速酶之一[11]。已被证实,大多数维管植中存在多种4CL同工酶。在模式植物拟南芥基因组中发现了14个4CL基因或类似基因[12]。其中只有4CL1、4CL2、4CL3及4CL5编码的蛋白具有催化活性,剩余的4CL基因及蛋白功能仍不清楚[13]。此外,拟南芥中4CL1、4CL2及4CL5与木质素合成相关,4CL3与黄酮素合成有关[14]。在白洋麻中也克隆出一个4CL基因,该基因被认为与木质素合成有关[15]。山杨中已被鉴定出两个结构和功能不同的4CL基因Pt4CL1和Pt4CL2,其中Pt4CL1被认为与茎中木质素生物合成有关,Pt4CL2被认为参与黄酮类物质的生物合成[16]。本实验首次克隆出84K杨中的Pag4CL3基因和Pag4CL5基因,通过对Pag4CL3/4CL5编码的蛋白与其他杨树4CL蛋白比较发现,均含有SSGTTGLPKGV保守结构域及GEICIRG催化活动中心。序列比对和系统进化树分析发现Pag4CL3/4CL5蛋白与毛果杨4CL、毛白杨4CL及青杨4CL蛋白分为同一个分支,亲缘关系较近。已有报道称毛白杨4CL蛋白能催化咖啡酸(caffeic acid)和阿魏酸(ferulic acid)生成相应的大CoA硫酯,不能催化芥子酸[17]。拟南芥中只有4CL5能催化芥子酸形成相应的CoA硫酯,以用于合成木质素单体。推测Pag4CL3/4CL5同毛白杨4CL蛋白一样不具有催化芥子酸的能力。
4CL基因在植物不同组织中的表达存在差异。美洲山杨Pt4CL1在木质化的组织中特异性表达;Pt4CL2的mRNA丰度在茎及叶的表皮层中得到积累。欧芹花茎中4CL基因的表达量远高于非生殖茎中4CL基因的表达量[18]。在拟南芥中At4CL1主要在幼根及抽薹茎中表达;At4CL2主要在幼根中表达;At4CL3在花和果荚中表达,在花中表达量最高[19]。本实验克隆的Pag4CL3/4CL5在序列上具有极高的相似性,可能执行近似的功能,但组织部位的表达差异性又导致了其在空间上的分工差异。Pag4CL3在叶及茎中表达较高,根和顶芽叶中表达量较低;Pag4CL5在叶及根中表达较高,茎和顶芽中表达量较低。推测Pag4CL3可能主要在叶和茎中行使功能,Pag4CL5主要在叶和根中行使功能。杨树作为重要的经济树种,是造纸工业的重要原材料。上述结果为今后研究杨树4CL基因的功能及改良木质素生物合成奠定了理论基础。
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
今日农业(2020年24期)2020-12-15 16:16:00
上海包装(2019年8期)2019-11-11 12:16:14
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
天津造纸(2016年1期)2017-01-15 14:03:29
兽医导刊(2016年12期)2016-05-17 03:51:50
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
中国造纸学报(2015年1期)2015-12-16 19:35:25
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
