
长白落叶松过氧化氢酶LoCAT1基因克隆及表达分析
2019-07-19 10:30:42白晓明董实伟杨宇宁张含国李淑娟
植物研究 2019年4期
白晓明 董实伟 杨宇宁 宋 跃 张含国 李淑娟
(林木遗传育种国家重点实验室,东北林业大学,哈尔滨 150040)
植物在逆境胁迫下,活性氧(Reactive oxygen species,ROS)的产生和积累会带来的氧化逆境毒害。ROS不仅可以直接对体内的大分子物质(如蛋白质、脂质以及DNA等)造成损伤,同时也作为信号分子参与信号转导、基因调控和氧化还原调控等过程[1]。此外还通过破坏电子传递链来抑制光合、呼吸以及氮代谢等过程,进而影响植物生长和发育[2]。在长期的进化过程中,植物细胞形成了一套有效的抗氧化防御系统以维持细胞内ROS的平衡,降低毒害作用。其中,过氧化氢酶(Catalase,CAT)在解除H2O2毒害的过程中发挥着重要的作用。
过氧化氢酶是抗氧化酶中首先被发现的,Loew将其命名为Catalase[3],广泛存在于动物、植物和微生物体内,在植物体内主要存在于叶绿体、线粒体、内质网中,而在动物体主要存在肝和红细胞中[4]。它能够清除植物细胞内光呼吸、线粒体电子传递及脂肪β-氧化等过程中产生的H2O2,从而保护细胞免于过氧化氢的毒害[5],通过提高植物体内抗氧化酶活性和增强抗氧化代谢水平可以提高植物本身的抗逆性[6]。研究表明,过氧化氢酶基因在病害防御、胁迫应答、延缓衰老等方面起着重要作用[7~9],如转过氧化氢酶基因KatG的棉花(Cotton)在干旱条件下生长优良,显著提高棉花的抗旱能力[10],转盐穗木(Halostachyscaspica)过氧化氢酶基因HcCAT1的大肠杆菌,可明显提高其耐盐性[11],过量表达的水稻(OryzasativaL.)过氧化氢酶基因OsCATb提高大肠杆菌对不同重金属逆境胁迫的耐受性[12]。近年来研究者已陆续从烟草[13](Tobacco)、拟南芥[14](Arabidopsisthaliana)、冬枣[15](Ziziphusjujuba)、银杏[16](GinkgobilobaL.)、丝瓜[17](Luffacylindrical)等多种植物中克隆出相关CAT基因,并对这些基因功能及表达特性进行了研究。
长白落叶松(Larixolgensis),又名黄花松,落叶乔木[17],是北方和山地温带干燥寒冷气候条件下寒湿型针叶林的主要树种之一。长白落叶松以早期速生、适应性和抗逆性强而著称[18],因此,它是进行抗逆基因克隆和抗逆机制研究的理想材料。本研究以长白落叶松为材料,克隆获得LoCAT1基因,运用生物信息学方法对其序列特征及编码蛋白结构等方面进行预测分析,同时对不同组织在逆境胁迫下该基因的表达模式进行了分析,旨在初步探究该基因的功能,为揭示林木逆境响应的分子机制奠定基础。
1 材料与方法
1.1 植物材料及胁迫处理
供试植物材料选自实验室播种育苗的长白落叶松植株。选取6个月苗龄,生长状态良好且长势一致的长白落叶松植株分别进行盐胁迫和渗透胁迫处理。盐胁迫处理方法:将长白落叶松植株转移到含有200 mmol·L-1NaCl的水溶液中,根部避光培养;渗透胁迫处理方法:将长白落叶松植株转移到含有40%(w/v)的PEG6000水溶液中,根部避光培养。以未处理的正常生长植株作为对照,处理12、24、48、96 h后,分别取各处理和对照长白落叶松的根、茎、叶,立即液氮冷冻处理,保存于-80℃备用中用于RNA提取。
1.2 试验方法
1.2.1 长白落叶松总RNA提取与cDNA第一链的合成
采用CTAB法分别提取根、茎、叶及整株的RNA,用超微量紫外分光光度计检测RNA的浓度和纯度,1%琼脂糖凝胶电泳检测RNA完整性。根据cDNA反转录试剂盒操作说明,分别以上述获得的RNA为模板,在PCR仪上进行逆转录,合成cDNA第一链,于-20℃保存。
1.2.2 特异引物设计与克隆
从长白落叶松转录组数据库中获得CAT1基因的全长序列,利用Primer5.0软件设计引物(表1),引物由上海生工生物公司有限公司合成。以整株的RNA反转录所得的cDNA为模板,采用KOD-FX PCR酶进行PCR扩增。PCR反应体系(50 μL):上下游引物各1.5 μL,模板cDNA 1 μL,KOD FX 1 μL,ddH2O 10 μL,2 mmol·L-1dNTPs 10 μL,2x PCR buffer for KOD FX 25 μL。扩增条件:94℃,2 min;98℃,10 s,58℃,30 s,68℃,55 s,35个循环;68℃,10 min。1%琼脂糖凝胶电泳对扩增产物进行检测并回收目的片段,将目的片段与pROK-Ⅱ载体连接,并转化到大肠杆菌DH5α感受态细胞中,卡那霉素(50 mg·L-1)筛选阳性转化子。PCR检测后将阳性结果送上海生工测序。

表1 引物序列
1.2.3LoCAT1基因的生物信息学分析
利用DNAMAN软件对获得的长白落叶松LoCAT1基因进行核苷酸序列的分析,并检测其编码蛋白质氨基酸序列。利用Protparam(http://web.expasy.org/protparam/)在线软件分析LoCAT1蛋白质的基本理化性质。利用ProtScale(https://web.expasy.org/protscale/)在线软件分析LoCAT1蛋白质亲水疏水性。利用NCBI数据库中的Blast对LoCAT1氨基酸序列进行同源序列比对,然后选择不同物种中的同源氨基酸序列。首先使用ClustalX对氨基酸序列进行多序列比对,再结合Mega 7.0软件构建系统进化树。
1.2.4LoCAT1基因的组织特异性表达检测
根据LoCAT1的基因序列设计特异定量引物LoCAT1-FQ及LoCAT1-RQ,以tubulin为内参设计引物tubulin-F及tubulin-R(表1),以1.2.1获得的根、茎、叶cDNA为模板,进行实时定量RT-PCR分析。体系如下:2xTransStart® TOP/Tip Green qPCR Supermix 10 μL,LoCAT1-FQ/RQ混合引物(10.0 μmol·L-1)0.4 μL,cDNA 1.5 μL,Passive Reference Dye(50x)0.4 μL,加ddH2O至20 μL。RT-PCR反应程序如下:94℃ 30 s;94℃ 5 s,60℃ 15 s,72℃ 35 s,40次循环;95℃ 15 s,60℃ 1 min,95℃ 30 s,利用2-△△CT法进行数据处理。
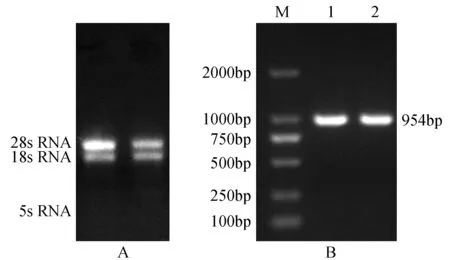
图1 长白落叶松总RNA(A)和扩增产物电泳图(B)Fig.1 The electrophoresis of total RNA in L.olgensis(A) and PCR production of LoCAT1(B)

图2 LoCAT1基因序列及推测的氨基酸序列Fig.2 The nucleotide and predicted amino acid sequences of LoCAT1
2 结果与分析
2.1 长白落叶松总RNA的检测及LoCAT1基因的克隆
提取的RNA经1%琼脂糖电泳检测,有明显的18 s RNA、28 s RNA两条条带和微弱的5 s RNA弥散状条带,且28 s RNA的亮度为18 s RNA的两倍(图1A),结果表明提取的总RNA质量完好,可用于后续试验。
以长白落叶松cDNA为模板,利用特异性引物对LoCAT1基因ORF序列进行扩增,扩增产物经1%琼脂糖凝胶电泳检测,结果显示与预期目的片段954 bp大小基本一致(图1B),条带清晰且无明显拖尾。将菌液送至上海生工生物公司测序,结果表明:LoCAT1基因共含有954个碱基,编码317个氨基酸(图2);LoCAT1基因克隆成功,可以进行后续操作。
2.2 长白落叶松LoCAT1基因生物信息学分析
2.2.1 LoCAT1蛋白质的理化性质分析
通过Protparam分析显示,长白落叶松LoCAT1基因蛋白分子式为C1675H2520N456O477S13。对其所编码氨基酸序列进行生物信息学分析,结果显示:LoCAT1蛋白分子量为56.81 kD,含有带负电的氨基酸(Asp+Glu)和带正电的氨基酸(Arg+Lys)个数分别为44和40个,理论等电点(pI)为6.21,脂溶指数68.26,总平均亲水系数为-0.640,不稳定系数为33.42,说明该蛋白为稳定的脂溶性亲水蛋白。
2.2.2LoCAT1基因的同源性比较及进化树分析
根据Blast同源搜素结果,将长白落叶松LoCAT1氨基酸序列与GenBank登录的北美云杉(Piceasitchensis)、银杏(Ginkgobiloba)、拟南芥(Arabidopsisthaliana)、江南卷柏(Selaginellaemoellendorfii)、麻风树(Jatrophacarcas)、枇杷(Eriobotryajaponica)等12个物种氨基酸序列进行同源性分析。结果表明,在比较的12个物种中,与北美云杉氨基酸序列相似性最高,达到93.7%,与枇杷的相似性最低,仅为68.1%(图3)。用MEGA7.0软件通过近临相接法(Neighbor-Joining)构建蛋白序列的进化树,由系统进化分析表明,科属亲缘关系接近的植物归纳为同一类,预测长白落叶松和北美云杉的距离最为接近,和其他植物均相差较远,结果与植物系统分类一致(图4)。
2.3 LoCAT1基因的组织特异性表达分析
以长白落叶松3个不同组织部位的cDNA为模板,通过RT-qPCR检测分析各组织部位的相对表达量(图5)。以LoCAT1基因在根中的相对表达量为参照,在叶中的表达量最高接近根的10倍,茎的表达量低于根。
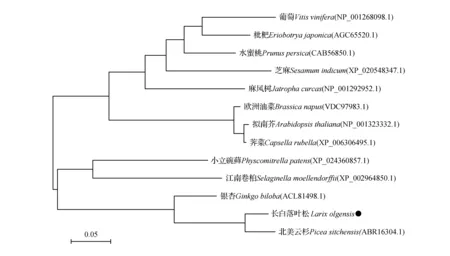
图4 LoCAT1基因系统进化树Fig.4 Phylogenetic tree of LoCAT1
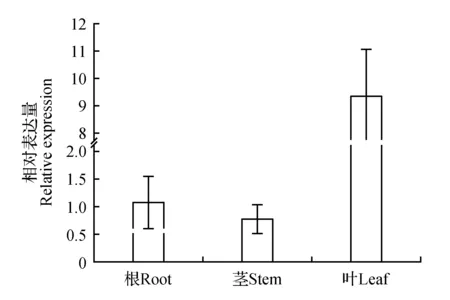
图5 LoCAT1基因组织特异性表达分析Fig.5 Tissue specific expression analysis of LoCAT1
2.4 LoCAT1基因在逆境胁迫下的表达模式
为了初步分析LoCAT1基因的逆境胁迫应答功能,利用qRT-PCR技术分析了200 mmol·L-1NaCl、40%(w/v)PEG6000胁迫处理后长白落叶松根、茎、叶中LoCAT1基因相对表达情况。结果表明:在NaCl处理后,根和茎中LoCAT1基因在所有时间点均表现为下调表达,其中12 h时表达量最低,仅为对照的38%和64%。叶中LoCAT1基因表达在24 h明显受抑制(仅为对照的24%),随后被上调表达,胁迫96 h时表达量最高,为对照的22倍(图6)。PEG6000处理后,根和茎中LoCAT1基因的表达在胁迫早期被明显抑制,随后被上调表达。叶中LoCAT1基因的表达在所有时间点均表现为上调表达,随着胁迫时间的延长LoCAT1基因的表达量逐渐增加,处理96 h时表达量最高,为对照的1.8倍(图7)。这些结果初步显示,高盐和渗透等非生物胁迫能够显著诱导LoCAT1基因的表达,由此推测该基因可能参与长白落叶松的逆境响应过程。
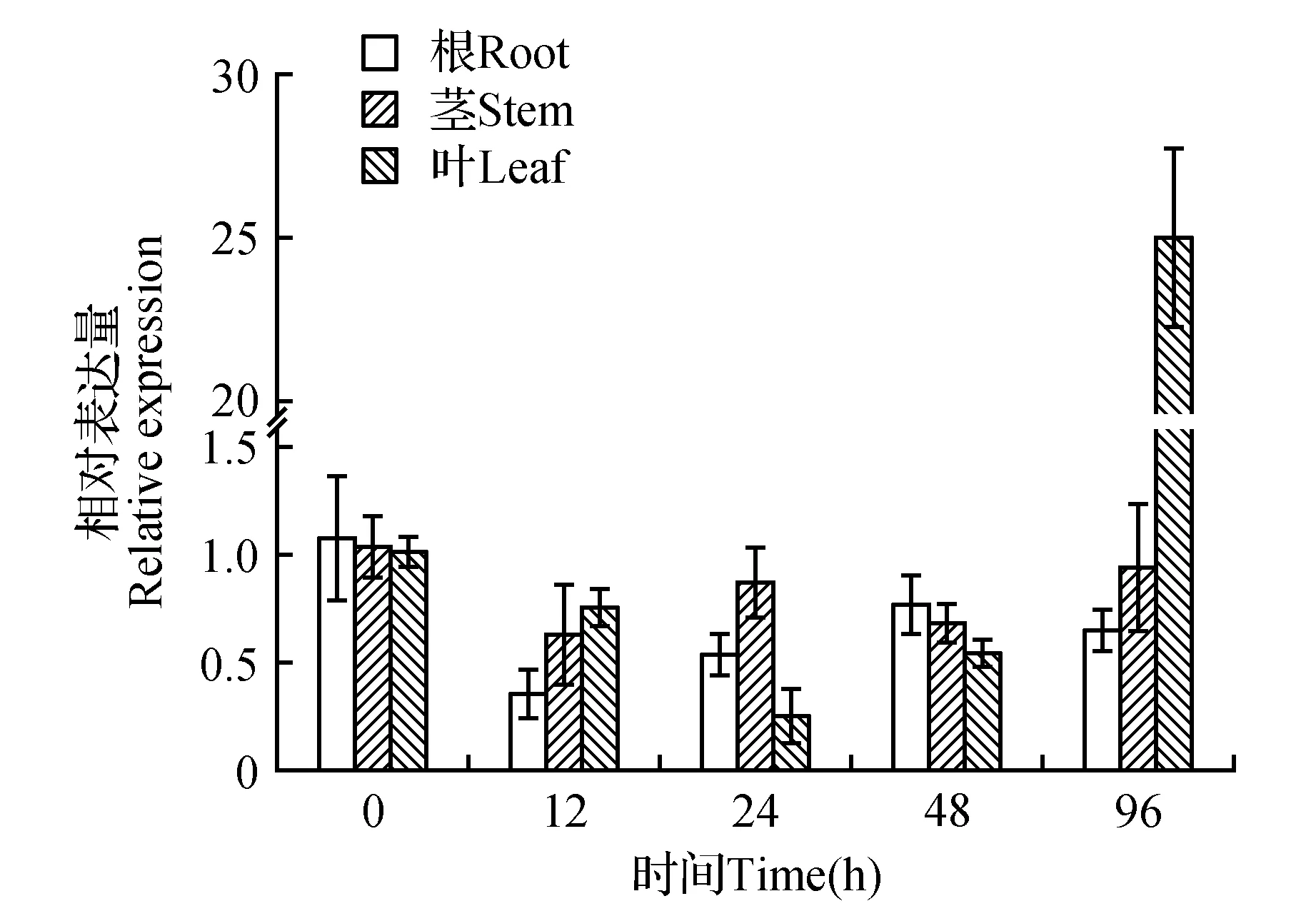
图6 NaCl胁迫处理下LoCAT1在长白落叶松根、茎、叶中的相对表达量Fig.6 Relative expression of LoCAT1 in L.olgensis root, stem and leaf with the treatment of NaCl
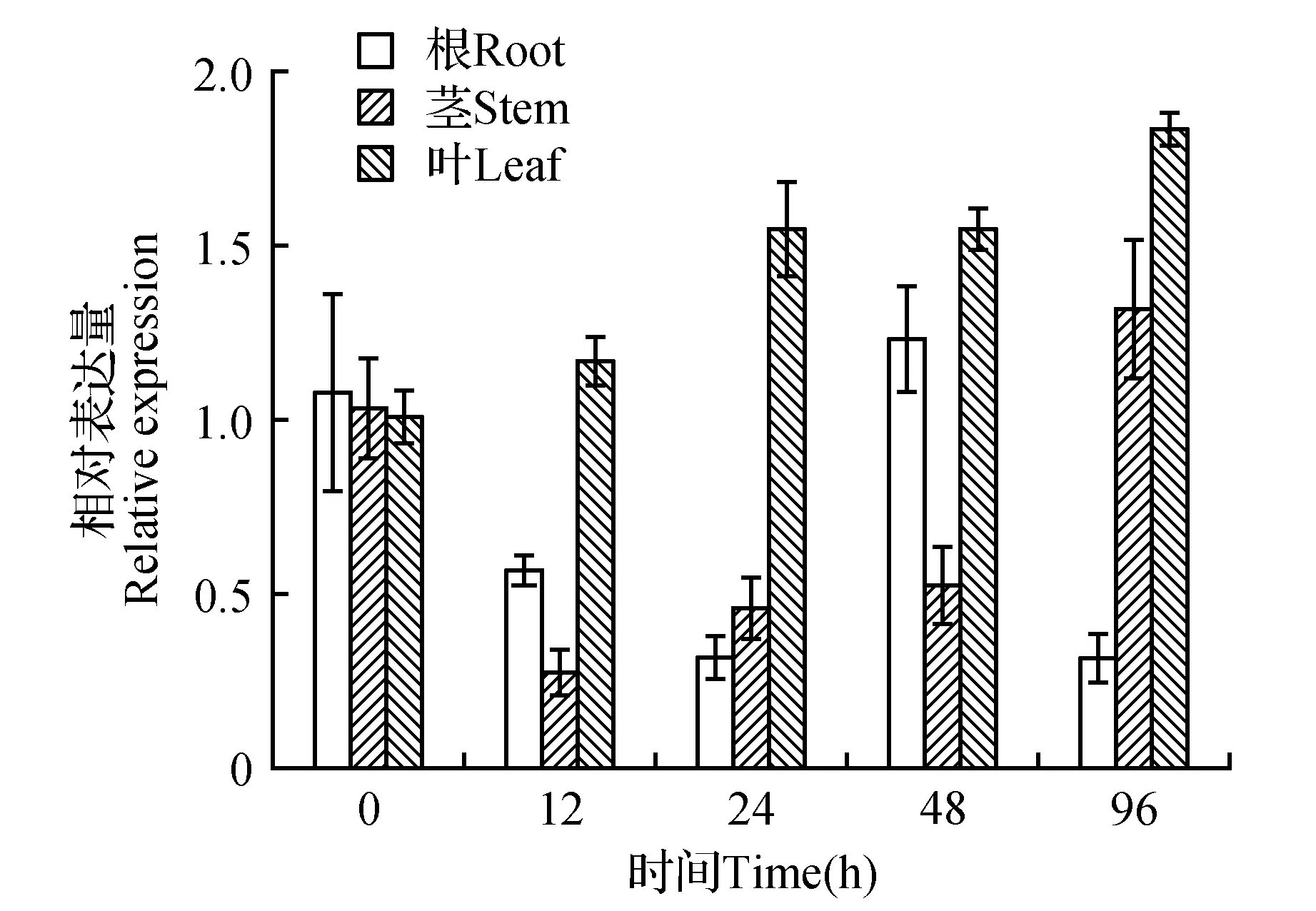
图7 PEG胁迫处理下LoCAT1在长白落叶松根、茎、叶中的相对表达量Fig.7 Relative expression of LoCAT1 in L.olgensis roots,stems and leaves with the treatment of PEG
3 讨论
植物在逆境胁迫下,过氧化氢酶对抵抗活性氧毒害发挥重要作用,该酶通过调节胞内ROS稳态,在植物的生长发育和抗逆反应中发挥重要的作用。本研究从长白落叶松中克隆得到LoCAT1基因,选取北美云杉等12种植物的CAT氨基酸序列进行多序列比对与进化树分析,结果显示,CAT蛋白氨基酸序列保守性较高。
CATs家族基因的分子表达特性与其所调控的植物生理代谢过程关系密切。在烟草中CAT1基因在叶内表达量高,CAT3基因主要在种子内表达,CAT2基因的表达受紫外、臭氧及病原诱导[20];南瓜(Pumpkin)CAT1基因在种子和幼苗发育早期表达,与衰老有关,CAT2基因在上胚轴和成熟叶内表达,而CAT3基因在下胚轴和根中表达[21];拟南芥CAT1基因、CAT2基因和CAT3基因在花序和种子内都表达,且CAT2基因和CAT3基因在叶中表达量高[22];蓖麻(Castor)CAT1基因主要在胚乳和胚轴中表达[23]。本研究通过RT-PCR对LoCAT1基因在长白落叶松的不同组织部位表达特征进行了分析。结果表明,LoCAT1基因在长白落叶松的根、茎、叶中都有表达,LoCAT1基因在叶组织中表达量最高,根次之,茎表达量最低。在叶片中高度表达,揭示LoCAT1可参与催化叶片光呼吸中产生的H2O2的分解作用,使机体免受H2O2的毒害,而相对于地下部的光呼吸产生的H2O2的清除则可能与其他同源基因的表达有关。
植物CAT基因的表达不仅具有组织器官的特异性,而且能够被逆境胁迫如低温、高温、盐害、干旱等诱导表达。例如,烟草和银杏在高盐和干旱的胁迫下,CAT1基因表达量上调,参与植物抵御逆境胁迫[13,16]。高羊茅FaCAT1基因在植株叶片受到高盐处理4 h后时,FaCAT1基因的表达量增加到最大[24],来清除活性氧造成的损害。干旱胁迫可使玉米CAT活性升高,从而减轻膜脂过氧化伤害,使玉米对干旱产生一定的抗性[25]。在本研究中,长白落叶松在非生物逆境胁迫下,LoCAT1基因的表达量都发生了明显改变,由此推测表明长白落叶松LoCAT1基因可能参与了渗透和耐盐胁迫的应答,但参与的调控途径不同。在后续研究中我们将继续对LoCAT1基因在长白落叶松抗逆过程中的具体功能和作用机制进行分析。
猜你喜欢
现代交际(2022年10期)2022-11-01 08:06:02
当代音乐(2022年8期)2022-04-29 16:02:48
今日农业(2021年10期)2021-11-27 09:45:24
当代音乐(2019年11期)2019-11-11 02:23:50
山东林业科技(2019年2期)2019-06-03 10:10:54
现代营销(创富信息版)(2018年2期)2018-08-15 00:45:27
发明与创新(2015年37期)2015-02-27 10:40:35
机电信息(2014年5期)2014-02-27 15:51:48
机电信息(2014年2期)2014-02-27 15:51:39
电源技术(2014年9期)2014-02-27 09:03:30
