
过表达 GhMYB4基因对棉花茎秆木质素合成的影响
2019-07-18 07:08:46夏成林于月华陈全家倪志勇
西北农业学报 2019年8期
夏成林,于月华,左 林,陈全家,倪志勇
(新疆农业大学 农学院,乌鲁木齐 830052)
MYB转录因子是植物中功能最多样化的家族之一,已被证实在多种植物中参与调控不同的代谢途径[1]。MYB转录因子具有1~4个由 50~53个氨基酸残基构成的不完全重复的高度保守的DNA结构域,即MYB结构域[2],根据其结构域位置和数目的不同,可将其分为4类(R1/2-MYB 或R3-MYB、R2R3-MYB、3R-MYB和4R-MYB)[3]。自从在玉米中克隆首个MYB转录因子基因 C1[4]后,在多种生物中也相继发现含有R2R3-MYB的同源基因,研究发现MYB 转录因子参与调节多种次生代谢途径,如苯丙烷类代谢途径、细胞壁组分合成和硫代葡萄糖苷的生物合成等,进而对植物次生壁的形成发挥调控作用[5]。
木质素(Lignin)是一种苯环结构的天然高分子物质,是构成植物细胞壁骨架的重要组成部分[6]。在植物维管组织的机械强度、抵抗病菌侵害和运输水分等方面发挥着重要作用[7]。近年来的研究表明,MYB转录因子与多种顺式作用元件结合,调控下游基因的表达。MYB转录因子与木质素代谢相关基因启动子序列中的一些顺式作用元件结合,如AC-Ⅰ、AC-Ⅱ、AC-Ⅲ等顺式作用元件,进而对木质素的合成产生促进或抑制作用[8]。木质素含量降低会造成作物株系易倒伏,易被病菌侵染等不良状况的发生[9]。为了克服这一现象,一些经济作物的改良需要适当增加木质素在茎秆中的合成积累,适当提高单位面积下细胞的数目,降低细胞排列分布的间隙,使改良的木质部导管壁韧性加强,从而可以在极大程度上降低病菌的感染入侵,对减少水分流失也有一定抑制作用[10]。
陆地棉(Gossypiumhirsutum)是世界上重要的经济作物之一。棉花的抗倒伏性状依赖于茎秆组织中木质素的机械支撑作用。因此,研究调控棉花茎秆木质素代谢的MYB转录因子的功能具有重要的意义。本研究旨在分析过表达 GhMYB4基因对陆地棉茎秆中木质素合成的影响。为深入研究 GhMYB4基因在陆地棉中的生物学功能奠定基础。
1 材料与方法
1.1 材 料
1.1.1 植物材料 对照受体棉花株系为‘cqj-5’,转基因过表达 GhMYB4棉花株系为T4代植株,编号为‘lrp1’~‘lrp29’。试验材料栽种于新疆农业大学棉花光照培养室培育。
1.1.2 试剂 甲醇、巯基乙酸、浓盐酸、氢氧化钠、木质素标准品、间苯三酚、超纯水和双蒸水。
1.2 方 法
1.2.1 棉花茎秆木质素化学组织染色 对35~55 d棉花茎秆材料进行化学组织染色。分别取对照受体和转基因型株系距离下胚轴0~1 cm、 2~7 cm、10~12 cm部位徒手制作临时切片。参考Wiesner法[11]在切片表面滴加w=15%间苯三酚和φ=10%盐酸溶液,漂洗后于光学显微镜下观察并拍照。
1.2.2 棉花茎秆木质素总量测定 样品取自 45 d棉花茎秆距离下胚轴2~7 cm处的对照受体株系和转基因株系茎秆,做3次生物学重复试验。参考Klason法[12],获得木质素-巯基乙酸(LTGA)沉淀物。用2 mL的0.5 mol/L氢氧化钠溶液溶解棕色沉淀。在紫外分光光度计下的280 nm处测定木质素溶液的吸光度。
1.2.3 棉花木质素标准品标准曲线测定 称取100 mg木质素标准品于100 mL的0.5 mol/L氢氧化钠溶液溶中,终质量浓度为1 mg/mL。在25 mL容量瓶中将木质素的终质量浓度再稀释成不同的6份,用紫外分光光度计在280 nm处测定溶液的吸光值。

表1 棉花木质素标准曲线质量浓度的测定Table 1 Determination of cotton lignin standard curve
1.2.4 棉花木质素质量分数测定 通过标准曲线的回归方程计算出棉花茎秆中木质素的质量浓度 ,再利用下面公式计算其质量分数。
木质素质量分数=C(μg/mL)×2(mL)× 0.001/0.03(g)
2 结果与分析
2.1 棉花茎秆木质素的化学组织染色
用Wiesner法对棉花茎秆进行染色,根据染色程度的深浅和范围可以初步反映出木质素的积累情况,以及所在的组织位置和分布情况。以下试验株系均是以转基因‘lrp17’株系为例与对照受体‘cqj-5’株系进行的结果比对分析。
2.1.1 初生木质部木质素染色范围距离分析 在电子显微镜10×物镜下观察木质素组织染色区域发现(图1),在距离下胚轴0~1 cm、2~ 7 cm、10~12 cm处进行对比,依据细胞排列的方向进行直线测量,发现大部分的‘lrp17’转基因株系相比对照受体‘cqj-5’株系在初生木质部区域的染色范围距离更长。在初生木质部染色范围距离的定量分析发现(图2),在距离下胚轴0~1 cm和2~7 cm中,转基因株系和对照受体株系在染色的范围距离上具有显著性的增加。
2.1.2 初生木质部木质素染色细胞层数分析 在电子显微镜40×物镜下观察初生木质部染色区域发现(图3),大部分转基因‘lrp17’株系初生木质部的细胞排列层数相比对照受体株系‘cqj-5’有一定增多。通过对细胞层数的定量分析发现(图4),在距离下胚轴2~7 cm处,转基因‘lrp17’株系相比对照受体‘cqj-5’株系具有显著性的增加。
2.1.3 初生韧皮部木质素染色深浅程度分析 在电子显微镜40×物镜下观察初生韧皮部染色区域发现(图5),在距离下胚轴0~1 cm、2~ 7 cm、10~12 cm处进行对比,‘lrp17’转基因株系相比对照受体‘cqj-5’株系的染色程度大部分都有显著性的加深。
2.2 棉花茎秆木质素总量测定分析
通过木质素标准品参与得到标准曲线测定的结果,用Excel计算得回归方程:y=312.941x- 9.565(R2=0.999 175,y为待测液质量浓度,x为待测液吸光度值)。将对照受体株系‘cqj-5’和‘lrp1’~‘lrp29’系列的每一个棉花株系的茎秆进行木质素总量测定,数值带入回归方程中。结果表明(图6),转基因株系中的‘lrp13’‘lrp15’‘lrp16’‘lrp18’和‘lrp29’株系相比对照受体‘cqj-5’株系的木质素质量分数没有出现显著性的增减差异,‘lrp19’株系的木质素质量分数出现显著性的减少现象,其他23个株系均与对照受体株系‘cqj-5’的木质素质量分数有显著性增加,其中‘lrp17’株系木质素的表达量最高。

a.初生木质部 Primary xylem;b.初生韧皮部 Primary phloem;刻度尺为10 μm The scale is 10 μm;下图3和图5同此 Fig.3 and Fig.5 are the same
图1 电子显微镜10×物镜下的35~55 d转基因‘lrp17’株系与受体株系‘cqj-5’(CK)棉花材料茎秆的横切木质素组织化学染色分析
Fig.1 Cross-cut lignin histochemical staining analysis of cotton stalks of 35-55 d transgenic ‘lrp17’ strain and recipient strain ‘cqj-5’(CK) under electron microscope 10×objective

数值是3次生物学重复的“平均值±标准差”,星号表示明显不同于对照受体cqj-5(P<0.05),下同 Values are “mean ± standard” deviation of three biological replicates,and asterisks indicate significantly different from control receptor cqj-5(P<0.05).The same bellow
图2 在电子显微镜40×物镜下的35~55 d转基因‘lrp17’株系与受体株系‘cqj-5’(CK)棉花材料茎秆的横切木质素组织化学染色距离分析
Fig.2 The cross-cut lignin histochemical staining distance analysis of the 35-55 d transgenic ‘lrp17’ strain and the recipient strain ‘cqj-5’(CK) cotton material stem under the electron microscope 40×objective lens
3 讨论与结论
之前的研究表明,在大多数植物生长发育进程中MYB转录因子对次生壁的表达调控存在促进或抑制作用[13-14]。本研究通过对棉花茎秆木质素染色分析发现,过表达 GhMYB4株系相比对照受体株系初生木质部的染色范围扩大、细胞层数增多,且初生韧皮部染色加深。大部分过表达 GhMYB4基因株系相比对照受体株系茎秆木质素质量分数增加,因此推测 GhMYB4转录因子可能正调控棉花茎秆的木质素代谢。

图3 电子显微镜40×物镜下的35~55 d转基因‘lrp17’株系与受体株系‘cqj-5’(CK)棉花材料茎秆的横切木质素组织化学染色分析Fig.3 Cross-cut lignin histochemical staining analysis of cotton stalks of 35-55 d transgenic ‘lrp17’ strain and recipient strain ‘cqj-5’(CK) under electron microscope 40×objective
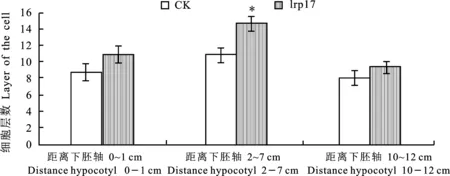
图4 在35~55 d转基因‘lrp17’株系与受体株系‘cqj-5’(CK)棉花材料茎秆的横切木质素组织化学染色分析下的细胞层数测量Fig.4 The number of cell layers under the cross-cut lignin histochemical staining analysis of the 35-55 d transgenic ‘lrp17’ strain and the recipient strain ‘cqj-5’(CK) cotton material stem
在许多植物中陆续发现多种MYB转录因子对次生壁中的木质素合成起到一定促进或抑制作用。如在拟南芥过表达菊花 CmMYB1基因能够在一定程度上抑制次生壁中木质素的合成积累[15]。拟南芥 AtMYB58和 AtMYB63[16],小麦 TaMYB4[17],以及玉米 ZmMYB31和 ZmMYB46[18-19]等基因都在次生壁发育过程中调控木质素的合成。火炬松 PtMYB4[20]与 Ptmyb1[21]和桉树 EgMYB2[22]转录因子通过与启动子区域中的AC顺式作用元件结合,从而对木质素的生物合成起到正调控作用;水稻 OsMYB46基因可以促进表皮细胞木质素和木聚糖纤维素的合成[23];拟南芥 AtMYB46基因受NST3的调控,通过同时对 AtMYB85和 KNAT7基因的多级转录调控,进而对次生壁的木质素含量产生抑制或促进作用[24]。本研究发现 GhMYB4基因在陆地棉茎秆中参与次生壁木质素的合成,本研究为进一步探究 GhMYB4基因的生物学功能奠定理论基础。
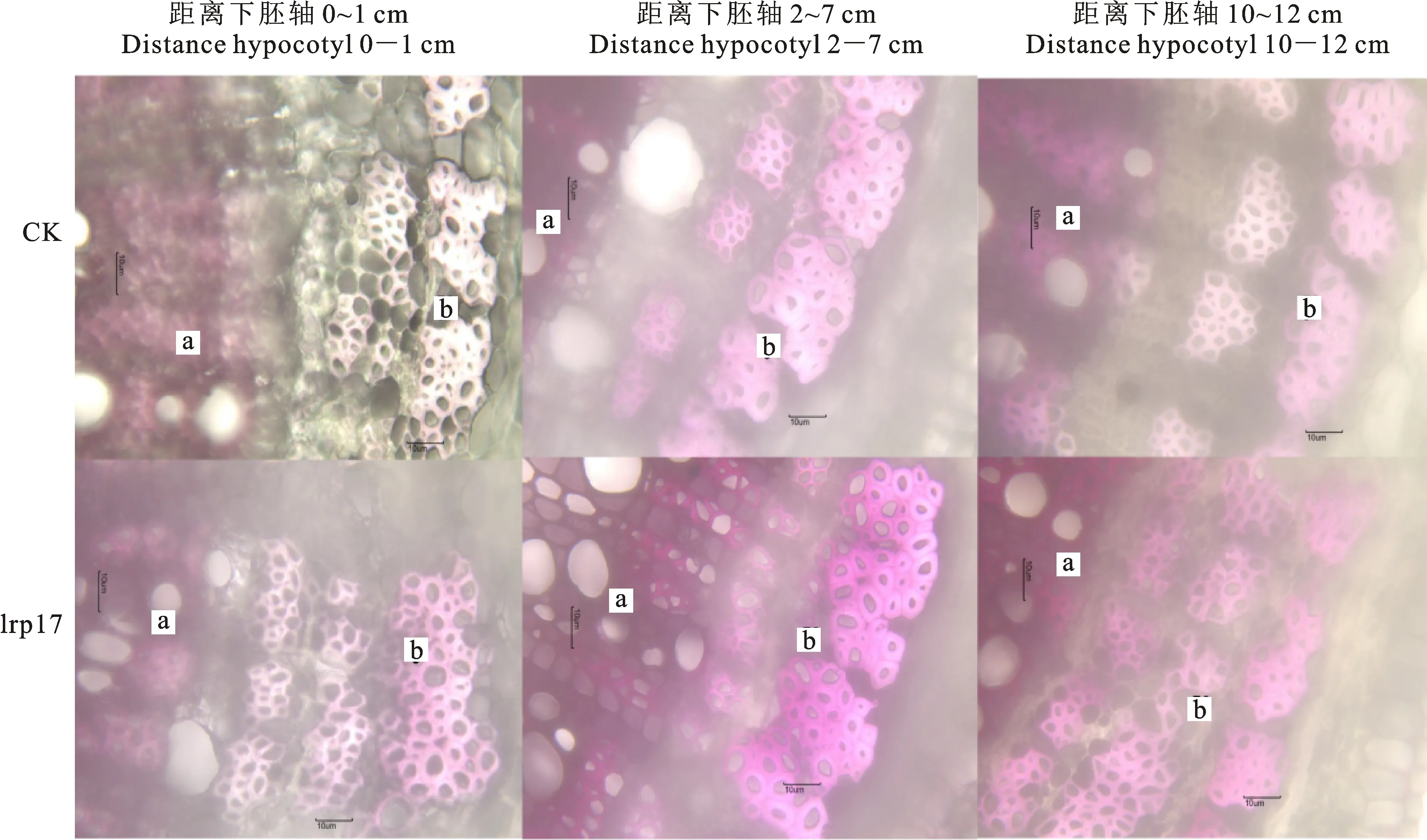
图5 电子显微镜40×物镜下的35~55 d转基因‘lrp17’株系与受体株系‘cqj-5’(CK)棉花材料茎秆的横切木质素组织化学染色分析Fig.5 Cross-cut lignin histochemical staining analysis of the 35-55 d transgenic ‘lrp17’ strain and the recipient strain ‘cqj-5’(CK) cotton material stem under electron microscope 40×objective
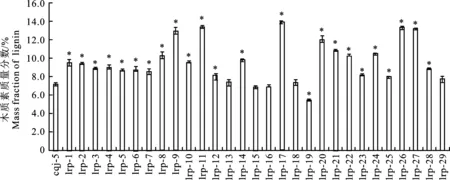
星号表示明显不同于对照受体‘cqj-5’(P<0.01) The asterisk indicates significantly different from the control receptor ‘cqj-5’ (P<0.01)
图6 受体‘cqj-5’(CK)和转基因‘lrp1’~‘lrp29’棉花株系茎秆中木质素质量分数
Fig.6 Mass fraction of lignin in the stems of the receptor ‘cqj-5’(CK) and the transgenic ‘lrp1’-‘lrp29’ cotton
猜你喜欢
江西农业大学学报(2022年3期)2022-07-07 04:00:52
亚热带植物科学(2022年1期)2022-05-17 12:39:32
植物研究(2021年2期)2021-02-26 08:40:10
四川农业科技(2019年5期)2019-07-01 09:46:46
农业机械学报(2019年4期)2019-04-29 02:20:48
畜牧与饲料科学(2018年5期)2018-06-13 12:03:48
浙江柑橘(2016年1期)2016-03-11 20:12:31
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
安徽农学通报(2014年7期)2014-04-29 03:00:24
