
长链非编码RNA uc002ktr.3在肺鳞癌顺铂耐药中的作用研究
2019-07-16 09:46:20王保龙
安徽医科大学学报 2019年6期
王 琪,罗 朋,王保龙
肺癌是全球发生率和死亡率很高的常见恶性肿瘤之一,居各种癌症死亡之首[1]。肺鳞癌是非小细胞肺癌(non small cell lung cancer, NSCLC)常见的病理类型。与肺腺癌相比,其发病机制的研究和治疗进展明显滞后[2]。目前铂类为基础的化疗仍然是肺鳞癌主要的治疗手段之一,但目前化疗失败的主要原因是其耐药性的产生[3]。长链非编码RNA(long non-coding RNA,lncRNA)是一类转录本长度超过200 nt的RNA,由于缺乏明显的开放阅读框,无蛋白质编码功能,曾一度被认为是基因组在进化过程中转录所产生的“噪音”,不具备任何生物学功能[4]。现已证实,lncRNAs与细胞周期、信号转导以及细胞凋亡等途径密切相关进而介导肿瘤的发生、发展、转移和药物耐受等[5]。前期本课题组通过肺鳞癌患者来源的肿瘤移植瘤(patient derived tumor xenograft, PDTX)模型筛选出肺鳞癌顺铂相对耐药和敏感的组织[6],为鉴定lncRNAs在肺鳞癌顺铂耐药中的调控作用,该研究拟检测肺鳞癌顺铂相对耐药和敏感的组织中lncRNAs表达谱,采用小干扰RNA技术(siRNA)下调长链非编码RNA uc002ktr.3的表达,研究uc002ktr.3对肺鳞癌顺铂耐受性的影响及机制,为肺鳞癌诊断和治疗提供新的靶标。
1 材料与方法
1.1 材料肺鳞癌SK-MES-1细胞株(中国科学院上海生命科学研究院);胎牛血清(美国Gibco公司);DMEM高糖培养基、0.25%胰酶(南京Wisent公司);TRIzol试剂、Lipofectamine 2000(美国Invitrogen公司);uc002ktr.3-siRNA及其阴性对照siRNA-NC(上海吉玛科技有限公司合成);荧光定量PCR检测试剂盒(日本Takara公司);Quick Amp Labeling Kit 荧光标记RNA试剂盒(美国Agilent公司);总RNA提取纯化试剂盒RNeasy Mini Kit(美国Qiagen公司);长链非编码RNA引物(安徽通用生物公司合成);顺铂、CCK8检测试剂盒(美国Sigma公司);β-actin、GAPDH、Zeb1和E-cadherin抗体(美国Proteintech公司);Annexin V/PI法细胞凋亡检测试剂盒、细胞总蛋白提取试剂盒(南京Vazyme Biotech公司)。
1.2 方法
1.2.1细胞培养 用10%胎牛血清的DMEM高糖培养基培养细胞SK-MES-1细胞,培养箱环境:37 ℃、5% CO2。待培养瓶内细胞生长达90% 融合度时进行传代,取对数生长期的细胞进行后续实验,整个培养过程中不加抗生素。
1.2.2LncRNA基因芯片检测及数据分析 抽提样品的总RNA,质检合格的RNA进行纯化。RNA样本通过Quick Amp Labeling Kit进行标记,杂交后清洗、甩干。通过RNeasy Mini Kit纯化标记的cRNA。用芯片扫描仪采集杂交后的图像,使用Agilent Feature Extraction软件(v11.0.1.1)处理相关数据。设定Fold Change≥2为数据筛选的标准,获取lncRNAs的差异表达谱。将表达基因或者差异基因数据做聚类分析,通过聚类热图(heatmap)直观地展现基因在不同样本中的表达情况。
1.2.3细胞瞬时转染 用胰蛋白酶消化对数生长期的SK-MES-1细胞,按照40%~60%密度将细胞接种于6孔板,培养24 h后进行转染。按照Lipofectamine 2 000转染试剂说明进行操作,将3条uc002ktr.3-siRNA基因序列和siRNA-NC序列分别转染于SK-MES-1 细胞中。转染24 h后,用TRIzol试剂收集细胞并提取细胞总RNA,通过qRT-PCR验证uc002ktr.3转染后的干扰效果。
1.2.4CCK-8检测体外顺铂药物敏感性变化 将uc002ktr.3-siRNA和siRNA-NC分别转染SK-MES-1细胞24 h后,胰蛋白酶消化,以 3×104/孔密度接种细胞于96孔板中。24 h后更换含不同终浓度的顺铂培养基,分别以 0、2、4、6、8、16 μmol/L的浓度进行加药,以不加药的一组为阴性对照(NC)组,平行设置5个复孔。37 ℃孵育48 h后, 每孔加入10 μl的CCK-8,温箱继续培养4 h后,自动酶标仪测定490 nm处的吸光度(A)值。通过细胞的生长抑制率(%)公式,最终由软件计算出半数抑制浓度(half inhibition concentration,IC50)值。
1.2.5Annexin V FITC/PI法检测细胞凋亡率 以3×105/孔接种细胞于6孔板中,将uc002ktr.3 siRNA和NC分别转染 SK-MES-1细胞,转染24 h后,加入终浓度为20 μmol/L的顺铂。12 h后收集细胞,PBS洗涤细胞3次,按Vazyme凋亡检测试剂盒操作说明采用Annexin V FITC/PI法双染,在1 h内上流式细胞仪检测,检测试验组和对照组细胞的凋亡率变化。通过公式计算细胞的细胞凋亡率(%)=(细胞早期凋亡数+细胞晚期凋亡数)/总细胞数×100%。
1.2.6Western blot检测转染后细胞Zeb1及E-cadherin蛋白表达变化 将uc002ktr.3-siRNA和siRNA-NC分别转染SK-MES-1细胞,作用72 h后加入细胞裂解液裂解细胞,提取总蛋白,二辛可酸法(BCA 法)进行蛋白定量。以同等质量的蛋白上样并进行SDS-PAGE电泳,电转移分离后的蛋白质至硝酸纤维素(NC)膜上,电转90 min后用10%脱脂牛奶室温封闭3 h,再分别加入Zeb1(1 ∶1 000)、E-cadherin(1 ∶1 000)和GAPDH(1 ∶6 000)抗体,4 ℃孵育过夜。TBST洗膜5次,每次6 min。加入二抗(1 ∶6 000稀释),室温孵育1 h,TBST洗膜3次,每次10 min。进行ECL增强化学发光检测,以GAPDH作为内参,分析Zeb1和E-cadherin蛋白表达情况。
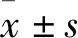
2 结果
2.1 相对耐药和敏感肺鳞癌组织中lncRNA的差异表达前期通过构建PDTXs,课题组筛选出顺铂相对耐药和敏感的肺鳞癌组织。本研究分别选取耐药和敏感的肺鳞癌组织各2例,根据lncRNAs 在不同样本中的差异表达水平进行聚类分析发现,相对于顺铂敏感组,耐药组中差异表达上调2倍的lncRNAs有644个,表达下调2倍的lncRNAs有576个。系统树状热图显示样本差异上调变化最大的前20个lncRNAs和下调变化最大的前20个lncRNAs(图1)。其中每列代表一个样本,R组代表顺铂耐受,S组代表顺铂敏感。每行代表一个lncRNAs,红色代表高表达,绿色代表低表达。最终选取lncRNAuc002ktr.3作为研究对象,芯片结果提示其在耐受组中表达显著上调。

图1 顺铂敏感和耐药肺鳞癌组织中lncRNA的差异表达
2.2 干扰uc002ktr.3后肺鳞癌细胞顺铂敏感性变化为进一步验证uc002ktr.3在肺鳞癌顺铂药物耐受性中所发挥的作用,首先通过siRNA干扰肺鳞癌SK-MES-1细胞中uc002ktr.3的表达,并由qRT-PCR检测基因干扰效率。3条siRNA(siRNA-1、siRNA-2和siRNA-3基因序列)分别转染细胞后,与siRNA-NC组相比,uc002ktr.3表达水平分别下降至(0.59±0.07)、(0.51±0.05)和(0.64±0.08),差异有统计学意义(F=61.691,P<0.01),见图2A。选取干扰效果最好的siRNA-2序列用于后续实验。通过转染siRNA降低SK-MES-1中的uc002ktr.3表达水平,利用CCK-8法检测uc002ktr.3-siRNA组与siRNA-NC组对顺铂药物敏感性变化。uc002ktr.3-siRNA组与siRNA-NC组对顺铂药物IC50分别为(7.4±1.6)μmol/L和(16.0±3.5)μmol/L,差异有统计学意义(t=-3.982,P<0.01),见图2B,结果提示降低uc002ktr.3可增加肺鳞癌细胞对顺铂的敏感性。
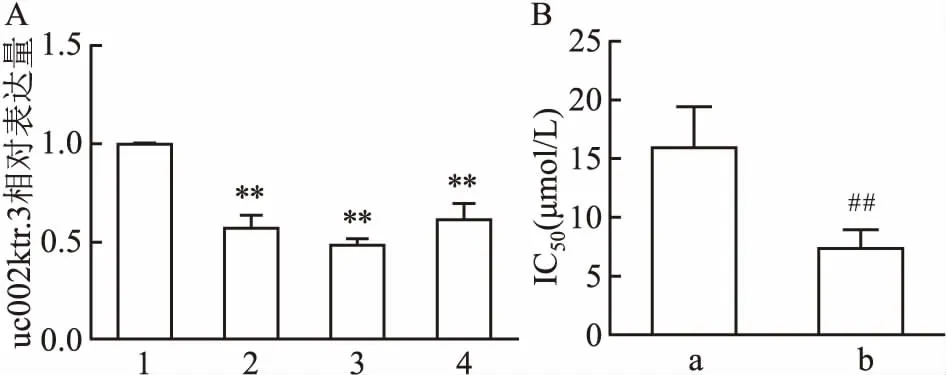
图2 干扰uc002ktr.3对SK-MES-1细胞顺铂敏感性的影响
A:3条siRNA干扰SK-MES-1细胞后uc002ktr.3的相对表达水平;1:siRNA-NC组;2:siRNA-1序列组;3:siRNA-2序列组;4:siRNA-3序列组;与siRNA-NC组比较:**P<0.01;B:uc002ktr.3-siRNA-2序列对SK-MES-1细胞顺铂IC50的影响;a:siRNA-NC组;b:uc002ktr.3-siRNA组;与siRNA-NC组比较:##P<0.01
2.3 干扰uc002ktr.3后顺铂诱导肺鳞癌细胞凋亡率的变化流式细胞术(FCM法)检测发现,在相同剂量顺铂药物(20 μmol/L)的诱导下,干扰了uc002ktr.3的SK-MES-1细胞凋亡率(39.93±2.03)%显著高于siRNA-NC组(20.63±1.90)%,差异有统计学意义(t=18.24,P<0.001),见图3。这表明干扰uc002ktr.3表达可增加顺铂诱导的细胞凋亡,从而增加肺鳞癌细胞对顺铂药物的化疗敏感性。
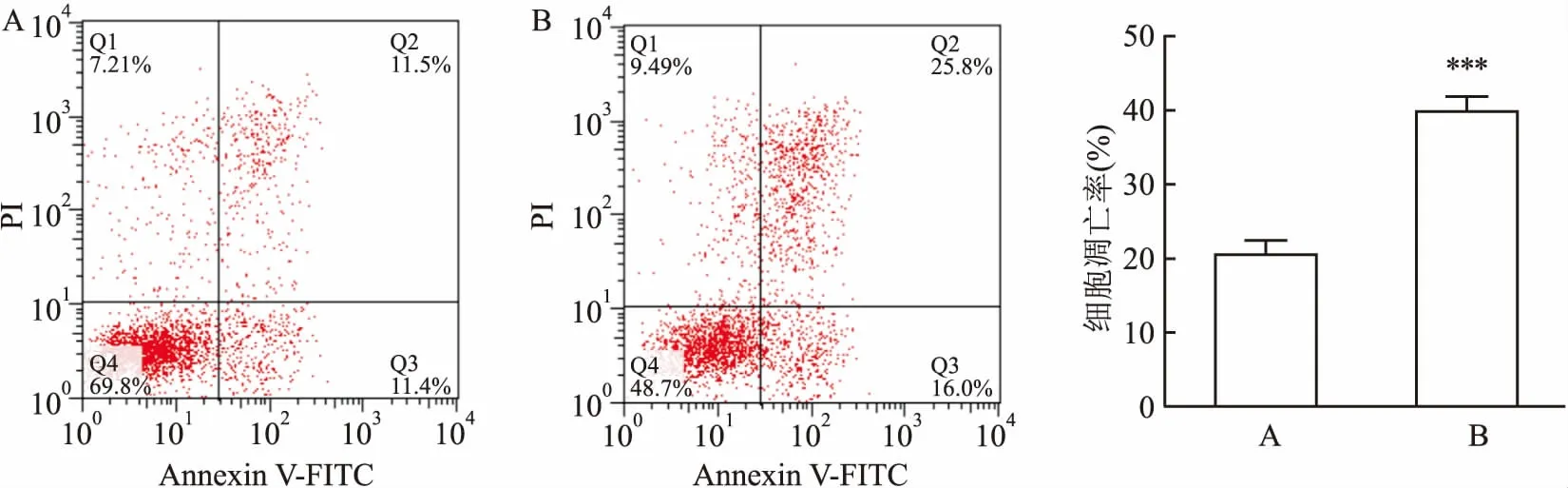
图3 干扰uc002ktr.3后流式细胞仪检测顺铂对细胞凋亡的影响
2.4 干扰uc002ktr.3后Zeb1和E-cadherin蛋白表达的变化Western blot结果证实,通过siRNA降低SK-MES-1细胞uc002ktr.3的表达水平后,相对于siRNA-NC组,Zeb1蛋白表达显著降低而E-cadherin蛋白表达显著增加(t=-12.84,P<0.01;t=7.20,P<0.01),见图4,说明高表达的uc002ktr.3可能通过上皮间质转化(Epithelial-mesenchymal transition, EMT)进而促进肺鳞癌细胞耐药表型产生。
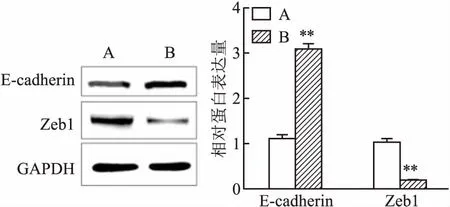
图4 Western blot法检测SK-MES-1细胞中Zeb1
A:siRNA-NC组;B:uc002ktr.3-siRNA组;与siRNA-NC组比较:**P<0.01
3 讨论
目前,肺腺癌的靶向治疗已取得显著进展,但由于肺鳞癌缺乏有效的分子靶标,以铂类药物为基础的化疗仍然是其治疗的主要手段,然而耐药却大大阻碍了肺鳞癌的治疗疗效[7]。顺铂抗肿瘤作用主要是通过作用于细胞的DNA链,造成 DNA不可逆性的损伤[8]。在顺铂诱导肿瘤细胞发生凋亡受到抑制时,细胞对顺铂的敏感性下降,继而发生肿瘤细胞顺铂耐药。近来研究[9-10]表明,大量化疗药物顺铂、卡铂、紫杉醇和长春瑞滨等耐药都与EMT密切相关, EMT是上皮来源恶性肿瘤获得侵袭迁移能力的重要生物学过程,目前已经成为肿瘤侵袭转移及治疗抵抗研究领域的热点。在EMT进程中,最主要的生物标记分子E-cadherin表达会减少,另外还涉及其他分子标记物的表达变化,如波形蛋白(Vimentin)、神经钙粘素(N-cadherin,N-cad)、β-联蛋白(β-catenin)及Zeb1蛋白等[11]。通过EMT,上皮细胞失去与基底膜的连接等上皮表型,获得了较高的侵袭迁移和抗凋亡等间质表型。Luo et al[12]发现,lncRNA H19能激活Wnt/β-catenin信号通路,影响EMT进程,从而促进膀胱癌细胞的侵袭转移。
随着高通量测序、基因芯片等技术的兴起,研究发现lncRNAs的差异表达水平与恶性肿瘤耐药密切相关。高表达的人母系表达基因(maternally expressed gene 3,MEG3)与肿瘤耐药密切相关,过表达的MEG3能通过活化p53,抑制Wnt/β-catenin信号通路中相关蛋白表达水平,从而诱导细胞凋亡,改善NSCLC患者的化疗敏感性[13]。长链非编码RNA GAS5(growth arrest-specifc transcript 5,GAS5)具有肿瘤抑制功能,高水平的GAS5能通过调节抑制细胞的自我吞噬,诱导细胞凋亡,来充分发挥抑癌基因的作用[14]。但是,目前相关的lncRNAs的研究尚在探索阶段,亟需进一步了解lncRNAs 在肺鳞癌的功能和机制,为临床诊治肺鳞癌患者提供更多的帮助。
为了探究lncRNA在肺鳞癌顺铂耐药中的分子机制,课题组前期利用已经构建好的肺鳞癌PDTX模型,成功筛选出了顺铂敏感和耐受的肺鳞癌移植瘤组织。本研究进一步通过对顺铂敏感和耐受的肺鳞癌组织中lncRNAs的表达谱进行分析,共有1 220个lncRNAs存在差异性表达(差异倍数>2.0),相对于顺铂敏感组,耐药组中差异表达上调2倍的lncRNAs有644个,选取了lncRNA uc002ktr.3作为研究对象。为进一明确步uc002ktr.3在肺鳞癌顺铂耐药中的作用,采用siRNA技术干扰uc002ktr.3表达,通过CCK-8实验证实了降低uc002ktr.3表达增加 SK-MES-1对顺铂的敏感性。在相同剂量顺铂药物的诱导下,流式细胞术结果表明这种耐药性与化疗药物诱导的凋亡相关,降低uc002ktr.3表达后,细胞凋亡比率显著增高。此外,降低uc002ktr.3表达,Western blot结果发现EMT相关蛋白Zeb1表达下降,E-cadherin表达显著增高,表明uc002ktr.3可能通过诱导EMT形成,进而影响肺鳞癌细胞顺铂耐药性。然而,uc002ktr.3诱导EMT、促进肺鳞癌耐药的具体机制,尚有待进一步深入探索研究。
猜你喜欢
云南医药(2021年3期)2021-07-21 05:41:06
四川文理学院学报(2020年5期)2020-02-12 02:05:58
中国有色金属学报(2018年2期)2018-03-26 07:58:26
焊接(2016年1期)2016-02-27 12:55:37
法医学杂志(2015年4期)2016-01-06 12:36:36
法医学杂志(2015年4期)2016-01-06 12:36:36
新闻传播(2015年8期)2015-07-18 11:08:24
癌变·畸变·突变(2015年3期)2015-02-27 06:15:13
肿瘤预防与治疗(2014年2期)2014-11-24 08:56:50
当代畜禽养殖业(2014年6期)2014-02-27 07:59:03
