
饲料脂肪源对大菱鲆生长、体组成及血清生化指标的影响
2019-07-12 07:59赵曼茜张斐然吴立新李思萌李笑迎
中国饲料 2019年13期
赵曼茜, 张斐然, 吴立新*, ,李思萌 ,2, 李笑迎
(1.大连海洋大学辽宁省水生生物学重点实验室,辽宁大连116023;2.聊城市开发区实验中学,山东聊城 252000)
饲料中的脂肪是鱼类必需脂肪酸的来源,具有多种生理功能,是鱼类重要的营养成分之一。有研究表明,不同脂肪源对鱼类生长、体组成和血清生化指标影响不同。冯健和覃志彪(2006)的试验结果表明,太平洋鲑(Oncorhynchus spp.)日粮中脂肪以添加大豆磷脂的生长性能最好;陈家林等(2011)研究发现,豆油、椰子油和菜籽油是异育银鲫 (Carassius auratus gibelio)饲料中良好的脂肪源;吉红等 (2009)建议生产中使用豆油作为鲫(Carassius auratus)饲料的脂肪源等。因此,研究不同鱼类适宜的脂肪源意义重大。
大菱鲆(Scophthalmus maximus)又名多宝鱼,是我国北方最重要的海水养殖鱼类之一 (马爱军等,2001)。本试验以鱼油(FO)、豆油(SO)、菜籽油(RO)、花生油(PO)、猪油(L)、鱼油-豆油(FSO)(1∶1)和混合油(MIX)(鱼油∶豆油∶猪油=3∶4∶3)为脂肪源的7种饲料,研究不同脂肪源对大菱鲆生长、体组成以及血清生化指标的影响,为筛选合适的大菱鲆饲料脂肪源提供一定的理论依据。
1 材料与方法
1.1 试验饲料的配制 本试验共配制了7种饲料,除脂肪源不同外,其余饲料成分相同,试验饲料组成及营养水平见表1。饲料蛋白源主要由鱼粉、豆粕提供,分别添加 6%的鱼油(FO)、豆油(SO)、菜籽油(RO)、花生油(PO)、猪油(L)、鱼油-豆油 (1:1)(FSO) 和鱼油-豆油-猪油(3:4:3)(MIX)。饲料原料粉碎后过60目筛,按试验配比充分混匀,用制粒机制作成粒径为2 mm的颗粒饲料,自然风干后存放于-20℃冰箱备用。
1.2 试验鱼与饲养管理 试验用大菱鲆购自大连天正实业有限公司。以基础饲料(FO)驯养两周。试验开始前,将鱼饥饿24 h,选取体质健康、规格均匀的鱼[(27.31±0.10)g]252 尾。试验共设 7组,每组设3个重复,共21个水族箱(60 cm×45 cm×40 cm,实际水量90 L),每个水族箱放入12尾鱼,饲养40 d。试验期间,以砂滤后的海水为水源,日换水一次,换水量为1/3~1/2。水温控制在14.0~ 19.0℃,溶解氧>6.0 mg/L,氨氮<0.1 mg/L,pH 8.0~8.1,光照采用日光灯控制,周期为12 L/12 D。每日饱食投喂两次(07:30和17:30),投喂后30 min左右收集残饵。
1.3 样品采集 试验结束时将鱼饥饿24 h,将每箱鱼分别称重(精确至0.01 g)并随机取3尾装于自封袋,置于-20℃冰箱中保存,用于常规体成分测定。每箱随机另取2尾于冰盘解剖,取其背肌、内脏和空壳,装入对应自封袋中置于-20℃冰箱待测。每箱随机另取2尾尾静脉取血,经3500 r/min离心15 min制得血清,置于-20℃冰箱待测。
1.4 样品分析测定 饲料和鱼体常规成分测定参照AOAC(1995)。水分通过在105℃烘箱烘至恒定质量测定;粗蛋白质含量用凯氏定氮仪测定(总氮×6.25);粗脂肪含量采用索氏抽提法,以乙醚为抽提液测定;粗灰分含量通过在马福炉550℃灼烧(8 h)测定;血清生化指标由大连市体检中心测定。每个样品重复测定2次,如果相对偏差超过2%,则增加重复测定次数,采用两个相对偏差在2%以下的测定值平均数作为测定结果。
1.5 计算公式
特定生长率/(%/d)=(lnWt-lnW0)/t×100;
蛋白质效率/%=(Wt-W0)/(Wd×Cp)×100;
肥满度/%=Wt/L3×100;
饲料系数=Wd/(Wt-W0)。
式中:W0和Wt分别为试验开始和结束时鱼的湿重,g;Wd为摄入的饲料干重,g;Cp为饲料蛋白质含量,%;L 为鱼体长,cm;t为试验天数,d。

表1 试验饲料组成及营养水平(风干基础)
1.6 统计分析 采用SPSS 17.0统计软件对所得数据进行单因素方差分析(one-way ANOVA),若差异显著(P<0.05),则进行Duncan's多重比较检验组间的差异,统计结果以“平均值±标准误”表示。
2 结果
2.1 生长性能 如表2所示,鱼油-豆油组的特定生长率显著高于其他各组,较鱼油组(基础饲料组)提高了10.42%(P<0.05),猪油组特定生长率最低,较鱼油组降低了19.79%(P<0.05);从蛋白质效率来看,除了鱼油-豆油组显著高于混合油组外 (P<0.05),其他各组间无显著差异 (P>0.05);饲料系数以鱼油-豆油组最低,但与鱼油、豆油、菜籽油组之间无显著差异(P>0.05),猪油、混合油、花生油组饲料系数分别较鱼油组提高了36.36%、33.33%、25.25%(P < 0.05)。
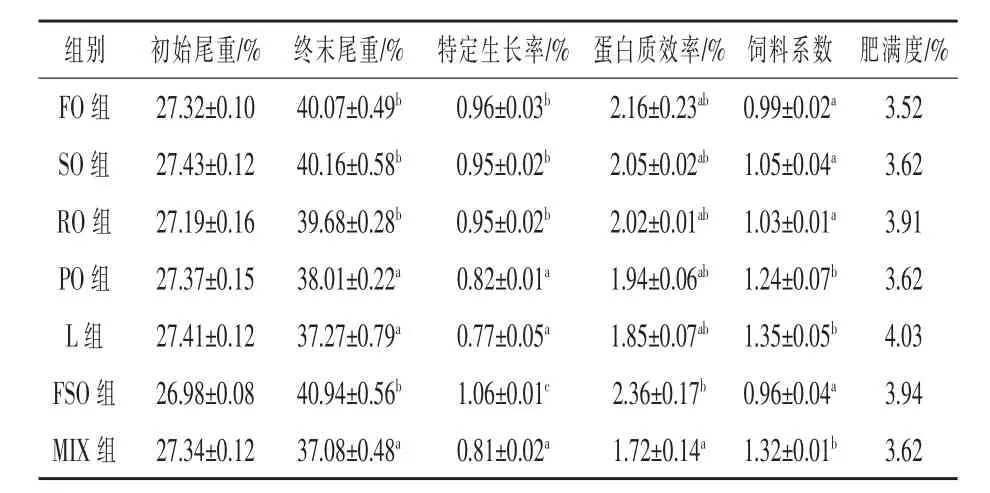
表2 饲料脂肪源对大菱鲆生长和饲料利用的影响
2.2 体组成和体组织水分、脂肪含量 如表3所示,饲料脂肪源对鱼体水分、粗脂肪和粗灰分含量均没有显著影响(P>0.05)。猪油组粗蛋白质含量显著低于菜籽油组、花生油组和鱼油组,其中较鱼油组降低了4.47%(P<0.05),其余各组间无显著差异(P>0.05)。从表4可以看出,除猪油组鱼体空壳的水分显著高于菜籽油组和混合油组 (P<0.05)外,饲料脂肪源对大菱鲆空壳、内脏、背肌的水分和脂肪含量没有显著影响(P>0.05)。

表3 饲料脂肪源对大菱鲆鱼体生化组成的影响%湿重
2.3 血清生化指标 如表5所示,各组间总胆固醇、甘油三酯、高密度脂蛋白胆固醇和碱性磷酸酶无显著差异(P>0.05)。猪油组和混合油组谷丙转氨酶含量显著高于其余各组,较鱼油组分别升高了124.89%和119.94%(P<0.05),其余各组间没有显著差异(P>0.05);混合油组谷草转氨酶含量显著高于其余各组,较鱼油组上升了352.51%(P<0.05),其余各组间无显著差异(P>0.05)。

表4 饲料脂肪源对大菱鲆各组织水分和脂肪含量的影响

表5 饲料脂肪源对大菱鲆血清生化指标的影响U/L
3 讨论
本试验结果显示,各组间特定生长率存在显著差异(P<0.05),以鱼油-豆油组最高,显著高于鱼油、豆油、菜籽油组和花生油、混合油、猪油组(P<0.05),这与张媛媛等(2012)用不同脂肪源饲料饲喂异育银鲫的研究结果相似。蛋白质效率除混合油组显著低于鱼油-豆油组(P<0.05)外,其他各组间饲料营养价值与鱼油无显著差异 (P>0.05)。从饲料系数来看,大菱鲆对鱼油-豆油组、鱼油组、菜籽油组和豆油组饲料利用率高。不同的饲料脂肪源对鱼体的肥满度没有显著影响,与於叶兵等(2012)用不同脂肪源饲料饲喂异育银鲫、杜震宇等(2002)对鲈鱼(Lateolabrax japonicus)的研究结果一致。
本试验中鱼体水分、粗脂肪含量在各组间没有显著差异 (P>0.05),这与真鲷(Pagrosomus major)(Glencross等,2003)、 异育银鲫 (陈家林等,2011)等的研究结果一致。通常不同脂肪源对鱼体组织的脂肪含量有一定的影响(Sargent等,2002),如卫晓怡和陈舜胜(2003)研究表明,大豆油添加组团头鲂(Megalobrama amblycephala)鱼体背部肌肉脂质积累较好,但在本试验中,各试验组空壳、内脏、背肌的脂肪含量均无显著差异(P > 0.05)。
鱼类血清生化指标能够反映鱼类对疾病的生理应答(Hrubec等,2008)。血脂含量占全身脂肪含量的很小一部分,但其代谢非常活跃,鱼类肠道吸收的外源性食物脂类,肝脏合成的内源性脂类和脂肪组织的利用都要经过血液再到其他组织,脂肪的运输也要经过血清,因此,血脂水平可以反映全身脂类代谢情况 (Hiraoka等,1979;Nakagawa,1978)。血清中总胆固醇和甘油三酯含量过高可能会导致心血管疾病,而高密度脂蛋白胆固醇可以将胆固醇从动脉壁运送到肝脏,通过肝脏转化代谢。因此高密度脂蛋白胆固醇有减轻血液中过量的胆固醇,减慢血管炎症,降低心血管疾病发生率的功能(孙明堂等,1982)。本试验中,各组间的总胆固醇、甘油三酯和高密度脂蛋白胆固醇含量没有显著差异(P>0.05),说明这7种脂肪源不会对大菱鲆幼鱼的脂肪代谢产生不利影响。谷丙转氨酶和谷草转氨酶存在于肝组织中,鱼类血液中的谷丙转氨酶和谷草转氨酶活性变化可以反映肝细胞受损情况 (康格菲,1989)。在正常情况下,鱼类血清中转氨酶的活性较低,血液中只含有少量的由鱼体组织细胞释放的转氨酶,当受损组织范围较大或组织病变引起细胞膜通透性增加时,使细胞主动转运体系受到损伤,导致物质主动转运机制和渗透压调节能力削弱从而消失,甚至细胞解体,血液中渗入由组织细胞内释放的大量谷丙转氨酶和谷草转氨酶,导致血液中谷丙转氨酶和谷草转氨酶活性升高(高露姣等,2005;惠天朝等,2000)。本试验中猪油组和混合油组的谷丙转氨酶含量显著高于其他各组(P<0.05),混合油组的谷草转氨酶含量显著高于其他各组 (P<0.05),这表明猪油组和混合油组鱼体肝脏受到一定程度的损伤,猪油和鱼油-豆油-猪油(3:4:3)不适宜作为大菱鲆的饲料脂肪源。这与高露姣等(2005)在饲料中添加猪油饲喂施氏鲟的研究结果一致。
4 结论
本试验结果表明,鱼油-豆油(1:1)、豆油和菜籽油可作为大菱鲆饲料合适的脂肪源,而猪油、混合油组鱼体肝脏可能受到一定程度的损伤,猪油、混合油不适宜作为大菱鲆的饲料脂肪源。
猜你喜欢
中国生殖健康(2020年5期)2021-01-18
农村百事通(2019年3期)2019-03-26
中国水产(2019年3期)2019-03-25
环球时报(2018-08-30)2018-08-30
商周刊(2017年6期)2017-08-22
现代营销·信息版(2017年2期)2017-03-17
农村百事通(2016年18期)2016-11-05
河北渔业(2016年5期)2016-09-08
河北渔业(2016年4期)2016-07-30
家庭医药(2016年7期)2016-07-14
