
钾镁胁迫对巴西橡胶树花药苗生理特性及叶绿体超微结构的影响
2019-07-09 11:24薛欣欣吴小平罗雪华王文斌王大鹏张永发赵春梅
热带作物学报 2019年8期
薛欣欣 吴小平 罗雪华 王文斌 王大鹏 张永发 赵春梅


摘 要 本研究通過营养液培养试验,重点揭示橡胶树的部分生理特性、叶绿体超微结构等指标对钾、镁缺乏的响应机制。以巴西橡胶树热研7-33-97花药苗(简称花药苗)为试验材料,设置缺钾镁(-K-Mg)、缺钾(-K)、缺镁(-Mg)、正常(CK)4个处理,测定花药苗株高、茎粗、叶绿素含量、钾镁含量、可溶性糖组分、叶绿体超微结构等指标。结果表明:(1)与CK相比,-K-Mg处理显著降低花药苗株高和茎粗,降幅分别为12.2%和15.7%,叶片出现衰亡、凋落现象;-K-Mg、-K和-Mg处理叶片的光合色素含量均显著下降(P<0.05)。(2)钾、镁胁迫均显著降低花药苗各器官钾、镁含量,钾供应极显著提高各器官镁含量,而镁供应对各器官钾含量无显著影响。(3)与CK相比,-K-Mg、-K、-Mg处理的葡萄糖、蔗糖和果糖含量均显著升高,钾、镁交互作用对果糖含量影响显著。(4)与CK相比,-K-Mg、-K及-Mg处理显著降低花药苗叶绿体长度,降幅分别为26.9%、38.1%和19.6%;嗜饿体数目显著增加,增幅分别为211.8%、767.7%和138.2%;缺镁条件下叶绿体中淀粉粒数目增加。综上所述,钾、镁胁迫对橡胶树花药苗生长、叶绿体发育均具有明显的抑制作用,钾、镁供应对2种元素吸收方面存在明显的钾对镁的单方面拮抗。生产上重视钾、镁的合理施用及其二者的交互作用,对改善橡胶树的生长状况具有重要的实践意义。
关键词 钾;镁;养分胁迫;橡胶树;生理特性;叶绿体超微结构
中图分类号 Q945.78; S59 文献标识码 A
Abstract A hydroponic experiment was conducted to clarify the response mechanism of potassium and magnesium deficiency affecting on some physiological characteristics and chloroplast ultrastrucure of rubber trees. The anther culture seedling of Hevea brasiliensis 7-33-97 was selected as the research material, and four treatments of -K-Mg, -K, -Mg, CK (control) were designed. The plant height, stem diameter, chlorophyll content, nutrients concentration, soluble sugar components and chloroplast ultrastrucure of rubber seedling were analyzed, respectively. Compared with CK, the plant height and stem diameter significantly decreased by 12.2% and 15.7% respectively, and the premature senescence and dying off of seedling leaves appeared under the condition of -K-Mg. The chlorophyll content of the leaf under -K-Mg, -K and -Mg significantly decreased (P<0.05). The concentrations of potassium and magnesium in different organs decreased significantly under potassium and magnesium deficiency. Potassium supply decreased significantly magnesium concentrations in different organs, while no significant effects in potassium concentration by the magnesium supply. Treatments of -K-Mg, -K and -Mg significantly increased the contents of glucose, sucrose and fructose in seedling leaves compared with CK. There was significant effect of interaction between K and Mg on the content of fructose. The treatments of -K-Mg, -K and -Mg significantly decreased chloroplast length by 26.9%, 38.1% and 19.6%, and significantly increased the number of plastoglobulus by 211.8%, 767.7% and 138.2%, respectively. Meanwhile, the treatment of -Mg significantly increased the number of starch granules. In summary, the deficiency of potassium and magnesium significantly inhibited the development of plant growth and chloroplast. Potassium had an antagonism effect on the absorption of magnesium under normal magnesium level. Therefore, the rational application of potassium and magnesium and the interaction should be emphasized in production, which could improve the plantation condition of rubber trees.
Keywords potassium; magnesium; nutrient deficiency; Hevea brasiliensis; physiological characteristics; chloroplast ultrastructure
DOI 10.3969/j.issn.1000-2561.2019.08.008
钾和镁是植物生长发育必需的营养元素,二者均参与植物一系列的生理代谢过程,在光合作用、酶活性、叶绿体结构等方面发挥重要作用[1-4]。随着复种指数、产量水平的提升,农业生产中钾、镁营养的缺乏及离子间的不平衡逐渐成为我国农作物产量的重要限制因子[5]。我国缺钾土壤主要分布在长江以南地区,该区域有3/4的土壤处于缺钾水平[6],而缺镁土壤类型主要包括砖红壤、赤红壤和部分红壤,全国有54%的土壤需要不同程度地补充镁素肥料[7]。研究发现,钾和镁胁迫对碳水化合物在源库间的运输至关重要[8-9],缺钾和缺镁均会造成植物叶片中大量积累碳水化合物,抑制了编码光合酶的基因表达,进而降低叶绿素含量和光合能力[10]。钾离子和镁离子间存在明显的拮抗作用。Ding等[11]研究表明,高钾低镁处理会造成水稻缺镁,进而降低叶绿素含量和光合活性,而镁水平高低则对钾无显著影响。
巴西橡胶树(Hevea brasiliensis)是我国热带、亚热带地区重要的经济作物,其所产生的胶乳是重要的工业原料和战略物资[12]。刺激割胶技术应用近30 a来,我国胶乳产量得到大幅度提升,但同时胶乳收获带走了土壤中大量养分[13]。研究证实,现行的胶园施肥管理措施不合理,造成土壤肥力明显下降,土壤酸化加剧[14-15]。土壤养分的缺乏对胶乳产量及品质的提升造成潜在威胁[16-17]。其中,钾和镁以离子态存在,较其他离子极易流失,同时,橡胶树对缺钾和缺镁均比较敏感[18]。早期学者研究表明,土壤钾素供应不足时,胶树生长受抑制,开割树的胶乳在割口上早凝,排胶不畅,胶乳机械稳定性差,再生皮厚度降低、叶片黄化甚至脱落、产胶量下降[19];缺镁造成橡胶树黄叶病发病率增加、割期明显缩短、树皮及木质部的淀粉含量下降、产量下降[20]。钾和镁之间存在拮抗作用,比值失调影响养分的吸收利用及植株的生长[21]。因此,重视钾、镁营养及其相互作用对橡胶树生长发育的影响机制,对改善橡胶树体营养状况,提升胶乳产量及品质至关重要。然而,以往研究钾镁缺乏对橡胶树的影响大多停留在现象上,而对其生理响应及微观结构研究较少[20]。新一代种植材料-橡胶花药苗,是通过组织培养、经胚体发生途径获得的体细胞植株,是一种长势很好的速生材料,但其营养特性仍有待阐明。本研究以橡胶树热研7-33-97花药苗为试验材料,重点探讨钾镁胁迫对橡胶树花药苗的部分生理特性及叶绿体超微结构的影响,以期为橡胶树生产中钾镁的合理施用提供理论依据。
1 材料与方法
1.1 材料
巴西橡胶树热研7-33-97花药苗(简称花药苗),由中国热带农业科学院橡胶研究所华玉伟博士、徐正伟提供。
1.2 方法
1.2.1 试验设计 试验于2017年10月至2108年1月在中国热带农业科学院橡胶研究所人工气候箱中进行。选取生长良好且长势一致的3月龄花药苗植株,用自来冲洗掉附着在根系上的泥沙,再用去离子水冲洗,然后移栽到盛有5 L营养液的黑色塑料桶中进行培养,2株/桶,用脱脂棉固定。营养液按Hogland and Arnon(1950)配方:浓度为KCl 8.00 mmol/L、7H2O·MgSO4 2.00 mmol/L、4H2O·Ca(NO3)2 4.00 mmol/L、(NH4)2SO4 3.50 mmol/L、EDTA-Fe 0.05 mmol/L、H3BO3 0.10 μmol/L、MnSO4 0.10 μmol/L、7H2O·ZnSO4 0.03 μmol/L、Na2MoO4 1.00 μmol/L、5H2O·CuSO4 0.10 μmol/L,所用试剂均为分析纯。
人工气候箱内设常温(25 ℃/20 ℃)(昼/夜),光照强度(5000 lx),光周期为12 h/12 h(昼/夜),相对湿度85%。培养期间,采用通气泵每3 h通气1次,营养液每3 d更换1次。试验开始后第1周用1/4全量营养液预培养,第2周用1/2全量营养液处理,第3周之后用全量营养液培养,第4周开始进行胁迫处理,胁迫处理时间约60 d。试验设置4个处理,分别为缺钾镁(-K-Mg,KCl 0 mmol/L、MgSO4 0 mmol/L)、缺钾(-K,KCl 0 mmol/L)、缺镁(-Mg,MgSO4 0 mmol/L)、正常(CK),脅迫处理的其他营养液及CK处理按上述配方配制。每个处理设3次重复,每个重复10盆,共120盆。培养结束后,每个处理调查株高、茎粗等生长指标。
1.2.2 样品采集及测定 (1)钾和镁含量测定。将根、茎干、茎皮、叶、叶柄分开,分别于105 ℃杀青30 min,75 ℃烘干至恒重,烘干样品经粉碎、过1.0 mm筛后, 称取约0.1000 g植物样品, 采用H2SO4-H2O2消煮、原子吸收分光光度计测定植物样品钾和镁含量[22]。
(2)叶绿素含量测定。选择代表叶片,除去叶脉,选择叶片中部1/3处部位的样品,剪碎、称取0.1000 g的新鲜叶片样品,采用酒精∶丙酮=1∶1浸提24 h,测定叶绿素a和叶绿素b含量[23]。
(3)葡萄糖、果糖、蔗糖测定。称取1.000 g磨碎的干样,加入5 mL 80%乙腈提取30 min,10 000 r/min离心10 min,共提取3次,蒸干,加入5 mL 80%乙腈,取上清液过0.22 ?m有机滤膜,滤液采用Waters-e2695型高效液相色谱仪测定可溶性糖组分含量。上机检测,进样量10 ?m。高效液相参数设定为:XBrindgeTM,Amide色谱柱(250 mm×4.6 mm,3.5 μm);流动相:乙腈-水(75∶25,V/V),在水相中加入0.2%的三乙胺,平衡pH;柱温为30 ℃,进样量为10 μL。乙腈和水的体积比为75∶25(V/V),流速为1 mL/min。蒸发光检测器:气体流速2 L/min,漂移管的温度为85 ℃,增益值为2。
(4)叶绿体超微结构观察。取花药苗成熟叶叶尖到叶基的中间部位(避开叶脉),切成1 mm× 2 mm的样块6~8片,放入pH 7.2的磷酸缓冲液配制成的4%戊二醛溶液的固定液中,具体试验步骤参考Lianopulou等[24]的方法。将样品在4 ℃固定24 h,用PBS缓冲液冲洗3次,经过30%—
50%—70%—80%—90%—95%—100%各级酒精进行梯度脱水,无水环氧丙烷置换酒精,然后进行树脂渗透,最后用环氧树脂包埋,放入烘箱中聚合。包埋后的材料用LKB型超薄切片机切片,经醋酸双氧铀和柠檬酸染色后,于JEM-1400Flash型电镜下观察。材料处理和电镜观察均在中国热带农业科学院环境与植物保护研究所电镜平台上完成。
1.3 数据处理
用Photoshop软件测量叶绿体的长度(纵向最远端两点之间的距离)和厚度(叶绿体横向中部最厚处的平均值)。采用SPSS 20.0软件对数据进行差异显著性分析,并用LSD法作多重比较,使用Origin 8.0软件制图。
2 结果与分析
2.1 钾镁胁迫对花药苗生长发育的影响
花药苗在不同处理下的生长发育情况如图1所示。不同钾、镁胁迫处理对花药苗的生长均具有明显抑制作用,最直观的症状是叶片失绿,其中以-K-Mg和-K处理最为明显,同时叶面积也有不同程度减小。-K-Mg处理叶片呈提前衰亡、凋落的现象,-Mg处理主要表现为上部叶片失绿,而CK处理的叶片深绿,长势良好。
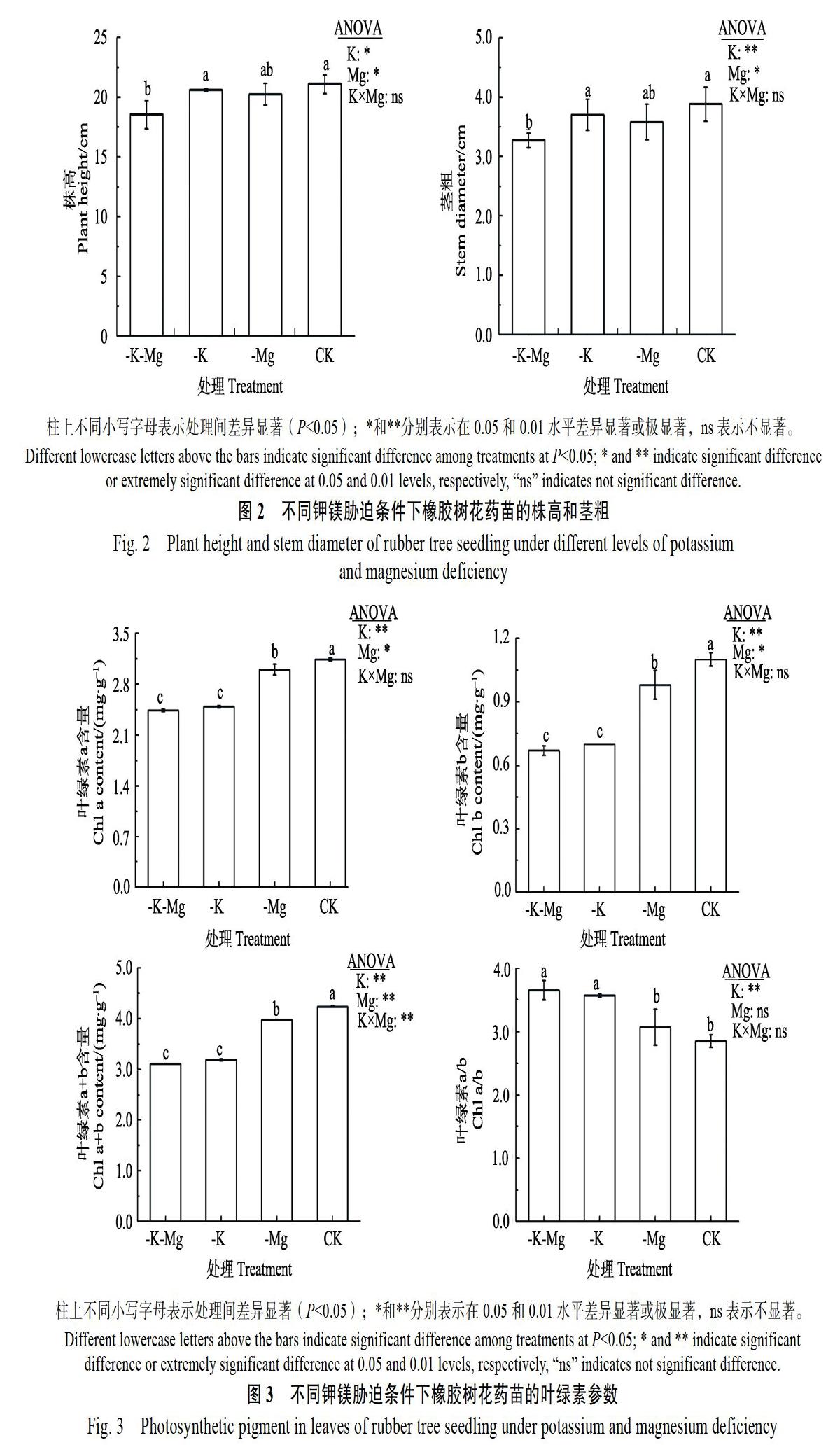
不同钾、镁胁迫处理均降低花药苗株高和茎粗(图2)。与CK比较,-K-Mg处理的株高和茎粗显著下降(P<0.05),降幅分别为12.2%和15.7%;-K-Mg、-Mg及CK间株高和茎粗差异不显著。方差分析显示,K、Mg供应水平对花药苗的株高和茎粗有显著(P<0.05)或极显著影响(P<0.01),K、Mg交互作用对橡胶树组培苗的株高和茎粗无显著影响。
2.2 钾镁胁迫对花药苗叶绿素含量的影响
不同钾、镁胁迫对叶绿素含量均有显著(P<0.05)或极显著(P<0.01)影响(图3)。钾缺乏条件下叶片叶绿素a、b及a+b含量显著降低。与CK处理相比,-K-Mg、-K、-Mg处理的叶绿素a含量降幅分别为22.3%、20.6%、4.6%,叶绿素b含量降幅分别为39.4%、36.7%、11.3%,叶绿素a+b含量降幅分别为26.8%、24.8%、6.3%。供钾条件下,镁缺乏均显著降低叶绿素a、b和a+b含量。方差分析表明,钾、镁交互作用对叶柱上不同小写字母表示处理间差异显著(P<0.05);*和**分别表示在0.05和0.01水平差异显著或极显著,ns表示不显著。
2.3 钾镁胁迫对花药苗各器官钾镁含量的影响
钾、镁缺乏分别显著降低花药苗各器官钾、镁含量(表1)。与CK相比,-K-Mg处理叶片、茎干、茎皮、叶柄和根的钾含量降幅分别为:34.6%、33.2%、39.6%、47.1%、24.3%,-K处理钾含量降幅分别为30.7%、35.9%、34.4%、41.4%、24.6%;-K-Mg处理叶片、茎干、茎皮、叶柄和根的镁含量降幅分别为:17.8%、33.2%、39.2%、37.2%、29.7%,-Mg处理钾含量降幅分别为22.8%、35.7%、44.4%、43.3%、45.8%。方差分析显示,无论施钾与否,镁供应水平对各器官钾含量无显著影响,而施镁条件下,钾供应显著或极显著降低各器官镁含量(P<0.05,P<0.01),方差分析显示,钾镁交互作用对茎干、茎皮和根的镁含量有显著或极显著影响(P<0.05,P< 0.01)。
2.4 钾镁胁迫对花药苗叶片可溶性糖组分的影响
钾、镁缺乏显著增加花药苗叶片的葡萄糖、蔗糖和果糖含量(P<0.01)(表2)。与CK相比,-K-Mg处理叶片的葡萄糖、蔗糖和果糖含量增幅分别为66.5%、16.8%和98.9%,-Mg处理增幅分别为22.4%、30.9%、81.9%,而-K处理显著提高葡萄糖和果糖含量,增幅分别为33.1%和49.0%,对蔗糖含量无显著影响。交互作用分析显示,钾镁交互作用对花藥苗叶片葡萄糖和蔗糖含量无显著影响,而对果糖含量具有极显著影响。
2.5 钾镁胁迫对花药苗叶片叶绿体超微结构的影响
花药苗叶绿体超微结构如图4所示。与CK(图4D)相比,钾、镁缺乏(图4A,图4B,图4C)显著增加叶绿体中的嗜饿体数目;-K-Mg(图4A)和-Mg(图4C)处理叶绿体中的淀粉粒数目明显增加,而-Mg处理的淀粉粒明显增大、叶绿体膨胀变形,出现解体的现象。-K(图4B)处理的叶绿体中未见到淀粉粒的积累。
通过对叶绿体结构参数的分析发现(表3),-K-Mg、-K及-Mg处理的叶绿体长度较CK显著降低,由正常的5.96 μm降为4.36、3.69、4.79 μm,降幅分别为26.9%、38.1%和19.6%。方差分析显示,钾及钾镁的交互作用对叶绿体长度均有极显著影响(P<0.01)。-Mg处理的叶绿体厚度较CK显著增加,增幅为16.1%。-K-Mg、-K处理与CK处理间叶绿体厚度无显著差异,镁对叶绿体厚度有极显著影响(P<0.01)。-K-Mg、-K及-Mg处理的单位叶绿体内嗜饿体数目较CK显著增加,增幅分别为211.8%、767.7%和138.2%;方差分析显示,钾、镁及钾镁交互作用对单位叶绿体内嗜饿体数目有极显著影响(P<0.01)。-K-Mg、-Mg处理的单位叶绿体内淀粉粒数目较CK显著增加,增幅分别为232.4%和243.2%,而-K处理的叶绿体内未观察到淀粉粒;方差分析显示,镁对淀粉粒数目有极显著影响(P<0.01)。
3 讨论
钾素对橡胶树的生长、产胶、排胶、抗风、抗病均有一定的影响;镁是叶绿素的重要组成部分,对橡胶树的光合作用起到直接作用,缺镁影响橡胶树的生长及产胶[25]。本研究发现,钾和镁的缺乏明显抑制花药苗的生长发育。首先,钾和镁缺乏明显影响叶片的正常发育,缺钾处理的花药苗叶片整体黄化,缺镁处理的花药苗上部叶片出现黄化现象,而钾和镁均缺乏条件下的花药苗叶片出现枯萎、衰亡现象。其次,研究结果表明钾、镁或钾镁共同缺乏均会造成叶绿素含量较CK显著降低。以往在成齡树上的研究也证实了钾和镁的缺乏会造成橡胶树黄叶病,而当土壤钾素过于丰富时,会抑制对镁的吸收,进而加剧黄叶病现象;钾、镁的缺乏还会造成橡胶树落叶期和抽芽期提前[26]。本研究结果还发现,钾和镁缺乏明显抑制了花药苗的株高和茎粗。以往研究表明,橡胶树缺钾造成茎围增长慢,延长了非生产期,造成开割时间推后,进而大幅降低胶乳产量[19]。根据本研究结果,钾、镁缺乏对橡胶树花药苗的生长发育具有明显的影响,生产中应重视钾和镁养分的合理供应。
钾、镁离子间的拮抗作用抑制橡胶树对镁的吸收,使得钾镁比率不平衡,从而影响橡胶树的生长和产胶[20]。本研究证实,钾、镁缺乏均显著或极显著降低花药苗各器官钾、镁含量;无论施钾与否,镁供应水平对橡胶树组培苗各部位钾含量均无显著影响,而钾供应水平对各器官镁含量均有显著降低的作用,该结果与丁玉川[5]在水稻上的研究结果相似。Ohno等[27]对小麦的研究表明,增加外源营养液中K+浓度会降低小麦地上部Mg2+含量,对根部Mg2+含量无明显影响;然而外源Mg2+浓度则对地上部和根部K+含量无影响。Ding等[11]对水稻的研究也表明,在高钾供应条件下,水稻叶片镁含量显著降低。研究发现,叶肉细胞Mg2+摄取有2个系统,一个是非选择性离子通道,另一个是H+/Mg2+的交换器(MHX),后者在低镁浓度下作用,非选择性离子通道也转运钾和钙,这些可能解释高钾和高钙抑制植物利用镁的现象[28]。
植株体内的可溶性糖含量是反映植株碳代谢水平的重要指标,其在抵抗环境胁迫时发挥重要作用。本研究结果表明,钾、镁缺乏显著增加花药苗叶片的可溶性糖含量,钾和镁的交互作用对叶片果糖含量具有极显著影响。该结果与前人的研究结果相似,吴敏等[29]对橡胶树幼苗的研究表明,缺钾胁迫造成橡胶树叶片和茎皮等部位的可溶性糖含量显著增加。薛欣欣等[30]对水稻的研究也证实,缺钾造成水稻叶片可溶性糖含量大量积累,比叶重增加,叶片增厚,导致稻谷产量显著下降。Pettigrew [31]研究表明,缺钾造成光合源器官的可溶性糖含量增加(主要为葡萄糖)。Ding等[11]的研究表明,缺镁造成水稻叶片可溶性糖含量显著增加。因此,当钾和镁胁迫时,可溶性糖在源器官中大量积累,限制了光合产物在源-库间的转运,进而影响植株的生长发育。
叶绿体结构和类囊体结构是保证植物光能吸收与转换的前提,钾和镁是叶绿体合成过程中的关键酶或辅酶基的活化剂,钾和镁缺乏会影响叶绿素的形成,进而影响植株光合作用。本研究结果表明,钾和镁的胁迫造成叶绿体中的嗜饿体数量增加,叶绿体长度减小;缺镁胁迫条件下,叶绿体中的淀粉粒数目明显增加,淀粉粒增大、叶绿体膨胀变形,出现解体的现象。Hamzah等[32]早期也证实了缺镁条件下,橡胶树幼苗叶片栅栏细胞比对照缩短、略厚,细胞中叶绿体数目减少,叶绿体的结构不完整,包被膜和其他膜呈解体状态,栅栏细胞中基质层状组织发育脆弱;缺钾条件下,橡胶树幼苗叶片的叶绿体体积明显小于对照处理。由此证实,钾和镁缺乏对橡胶树叶绿体结构产生明显影响,进而造成光合作用受限,最终影响植株正常的生长发育及胶乳的形成和产量。
4 结论
缺钾(-K)条件下,橡胶树花药苗长势弱、叶片黄化、叶绿素含量降低,各器官的钾含量显著降低但镁含量显著增加,叶片葡萄糖、果糖含量显著增加;叶绿体中的嗜饿体数量明显增多、叶绿体长度、厚度显著降低。
缺镁(-Mg)条件下,花药苗长势受限,各组织镁含量显著降低,叶片葡萄糖、蔗糖和果糖含量显著增加;叶绿体中的嗜饿体和淀粉粒数量显著增加,叶绿体长度显著降低而厚度显著增加。
缺钾镁(-K-Mg)条件下,橡胶树花药苗长势最弱,叶片出现衰亡;各器官钾和镁含量显著降低,叶片葡萄糖、蔗糖和果糖含量显著增加;叶绿体长度显著降低,嗜饿体和淀粉粒数目显著增加。
钾与镁离子交互作用对总叶绿素含量具有明显的正效应,但是对植株镁含量及果糖含量具有明显的负效应,同时钾的供应在镁供应提高镁含量的效应方面具有明显的单方面拮抗作用。
致谢:特别感谢中国热带农业科学院橡胶研究所华玉伟博士、徐正伟等为本研究提供橡胶花药苗供试材料。
参考文献
Tsialtas I T, Shabala S, Baxevanos D, et al. Effect of potassium fertilization on leaf physiology, fiber yield and quality in cotton (Gossypium hirsutum L.) under irrigated Mediterranean conditions[J]. Field Crops Research, 2016, 193(7): 94-103.
Farzadfar S, Zarinkamar F, Behmanesh M, et al. Magnesium and manganese interactively modulate parthenolide accumulation and the antioxidant defense system in the leaves of Tanacetum parthenium[J]. Journal of Plant Physiology, 2016, 202(9): 10-20.
Tatagiba S D, DaMatta F M, Rodrigues F A. Magnesium decreases leaf scald symptoms on rice leaves and preserves their photosynthetic performance[J]. Plant Physiology and Biochemistry, 2016, 108(11): 49-56.
Lu Z, Ren T, Pan Y, et al. Differences on photosynthetic limitations between leaf margins and leaf centers under potassium deficiency for Brassica napus L.[J]. Scientific Reports, 2016, 6(2): 21725.
丁玉川. 水稻鎂营养特性及镁钾营养互作效应研究[D]. 南京: 南京农业大学, 2007.
Z?rb C, Senbayram M, Peiter E. Potassium in agriculture–status and perspectives[J]. Journal of Plant Physiology, 2014, 171(9): 656-669.
白由路, 金继运, 杨俐苹. 我国土壤有效镁含量及分布状况与含镁肥料的应用前景研究[J]. 土壤肥料, 2004(2): 3-5.
Pan Y, Lu Z, Lu J, et al. Effects of low sink demand on leaf photosynthesis under potassium deficiency[J]. Plant Physiology and Biochemistry, 2017, 113(4): 110-121.
Ding Y, Luo W, Xu G. Characterisation of magnesium nutrition and interaction of magnesium and potassium in rice[J]. Annals of Applied Biology, 2006, 149(2): 111-123.
李文娟. 钾素提高玉米茎腐病抗性的营养与分子生理机制[D]. 北京: 中国农业科学院, 2009.
Ding Y C, Chang C R, Luo W, et al. High potassium aggravates the oxidative stress induced by magnesium deficiency in rice leaves[J]. Pedosphere, 2008, 18(3): 316-327.
李国华, 田耀华, 倪书邦, 等. 橡胶树生理生态学研究进展[J]. 生态环境学报, 2009, 18(3): 1146-1154.
罗雪华, 刘云清, 蔡秀娟, 等. 刺激割胶与RRIM600矿质养分流失的关系[J]. 热带农业科学, 2006, 26(1): 1-4, 24.
林钊沐, 罗 微, 茶正早. 海南橡胶树钾素营养状况及钾肥施用效果[J]. 热带农业科学, 2003, 23(5): 18-21, 65.
曹建华, 蒋菊生, 陶忠良, 等. 不同年龄橡胶树胶乳矿质养分的流失[J]. 热带作物学报, 2010, 31(1): 1-5.
林 葆. 我国平衡施肥中的中量和微量营养元素问题[C]. 中国平衡施肥报告会, 2001.
何 鹏. 低磷胁迫下橡胶树的生理应答及SSH文库构建与分析[D]. 海口: 海南大学, 2011.
吴 敏. 巴西橡胶树幼苗对低钾胁迫的生理响应及差异表达基因分析[D]. 海口: 海南大学, 2011.
王国烘. 我国植胶区的钾素营养问题[J]. 云南热作科技, 1991, 14(3): 11-14, 34.
许能琨. 氮镁磷钾肥对橡胶幼树生长量、叶片矿质养分含量和镁缺乏症的影响[J]. 热带作物学报, 1981, 2(2): 24-34.
邢 慧, 蒋菊生, 麦全法. 橡胶林钾素研究进展[J]. 热带农业科学, 2012, 32(4):42-48, 56.
鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000.
李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000.
Lianopoulou V, Bosabalidis A M, Patakas A, et al. Effects of chilling stress on leaf morphology, anatomy, ultrastructure, gas exchange, and essential oils in the seasonally dimorphic plant Teucrium polium (Lamiaceae)[J]. Acta Physiologiae Plantarum, 2014, 36(8): 2271-2281.
周艳飞. 云南橡胶树栽培[M]. 昆明: 云南大学出版社, 2008.
沈利宗, 李玉珍. 橡胶树富钾缺镁黄叶症的试验总结[J]. 热带农业科技, 1989, 12(3): 12-16.
Ohno T, Grunes D L. Potassium-magnesium interactions affecting nutrient uptake by wheat forage[J]. Soil Science Society of America Journal, 1985, 49(6): 685-690.
盛翊佳, 丁亚文, 傅媛烨, 等. 植物应答镁营养胁迫机理的研究进展[J]. 植物学研究, 2015, 4(5): 97-106.
吴 敏, 何 鹏, 韦家少, 等. 钾水平对巴西橡胶树幼苗叶片叶绿素及可溶性蛋白含量的影响[J]. 中国农学通报, 2011, 27(28): 58-62.
薛欣欣, 鲁剑巍, 李小坤, 等. 不同施氮量下缺钾对水稻叶片营养及生理性状的影响[J]. 植物营养与肥料学报, 2016, 22(6): 1494-1502.
Pettigrew W T. Potassium deficiency increases specific leaf weights and leaf glucose levels in field-grown cotton[J]. Agronomy Journal, 1999, 91(6): 962-968.
Hamzah S B, Gomez J B, 孔宪章, 等. 缺矿物元素的橡胶树叶片的超微结构Ⅲ、量的分析[J]. 热带农业科技, 1985, 5(3): 41-45.
猜你喜欢
农民致富之友(2019年29期)2019-10-21
散文选刊·下半月(2018年2期)2018-02-23
世界热带农业信息(2018年7期)2018-01-19
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
湖北农业科学(2016年20期)2017-02-15
山东农业科学(2016年12期)2017-01-21
世界热带农业信息(2016年6期)2016-07-02
湖北农业科学(2015年17期)2015-10-09
热带农业工程(2014年5期)2014-12-31
