
作物高油品种选育策略研究进展
2019-06-28 01:03:48张江江严江涛赵立宁李德芳
河南农业科学 2019年6期
张江江,常 丽,严江涛,赵立宁,李德芳
(中国农业科学院 麻类研究所,湖南 长沙 410205)
作为维持植物体生命活动的基础代谢之一,油脂代谢在植物生命周期中起着重要作用。种子脂肪是植物种子储存能量的主要形式,通过调控种子油脂含量以及脂肪酸组成相关途径可以实现作物高油品种的培育工作[1]。随着石油等不可再生能源的日趋减少,对新能源进行开发颇为急切,而植物油脂(如菜籽油)中有一些脂肪酸链长度与柴油接近,因此,植物油脂可作为生物能源的重要原料[2-3]。随着全世界人口的急速增加以及科技的日新月异,对食用油和生物燃料的量的需求以及质的追求也在不断提高。不饱和脂肪酸食用油对人体健康有益,而长链饱和脂肪酸对人体具有一定的危害,富含高油酸的食用油受到了追捧,其原因是高油酸食用油能有效预防高血脂、冠心病等心脑血管疾病[4]。因此,培育高油酸和特殊脂肪酸含量的作物品种具有重要的现实意义。
植物油脂的代谢调控过程复杂,涉及大量生物学过程和相关基因,这增加了植物油脂代谢解析及遗传机制研究的难度。目前,已有大量与油脂代谢相关的基因被鉴定和克隆出来。种子油脂的累积和组成由大量的基因和代谢因素调控,过去的15 a间,随着模式植物拟南芥全基因组测序后,在拟南芥中已经鉴定了多种与油脂合成相关的途径,并克隆和验证了相关途径中的一些关键基因[5]。归纳了植物油脂合成的一般途径和油脂改良的主要方式,从碳源分配、脂肪酸从头合成、三酰甘油的合成效率3个方面总结了植物油脂合成的关键步骤,并对这3个过程中关键基因的研究进展进行了综述,概括了作物高油品种选育的策略,旨在为作物高油品种的选育提供借鉴和帮助,并为植物油脂研究提供理论支撑和技术指导。
1 植物中油脂合成的一般途径
一般认为,油脂是油和脂肪的统称,三酰甘油是植物中储存油脂的主要形式,在很多植物中,三酰甘油为种子萌发、花粉发育、有性生殖等过程提供能量[6]。其主要储存在种子中,在其他营养组织中含量较少[7]。目前,在油料作物研究中发现,油脂的生物合成途径主要分为2种,一种是依赖于酰基-CoA的Kennedy途径,一种是不依赖于酰基-CoA的合成途径[8]。Kennedy生物合成途径的主要步骤包括:脂肪酸在质体中的合成、脂肪酸的碳链修饰、油脂的组装、油脂的富集与运输。其中,乙酰-CoA羧化酶(ACCase)和二酰基甘油酰基转移酶(DGAT)是2个关键的限速酶,它们能够直接调控种子形成过程中的油脂合成。在不依赖于酰基-CoA的合成途径中,磷脂二酰基甘油酰基转移酶(PDAT)是其主要限速酶。
2 植物油脂改良的主要方式
目前,植物油脂改良主要有以下几种方式,一是传统育种,优点是可定向改良油脂相关的某些性状,缺点是周期长、工作量大;二是通过诱变途径创造突变体,优点是周期较短,不需要很大的工作量,缺点是无法控制变异方向;三是基因编辑技术,优点是周期短,且能做到精准改良油脂相关性状,缺点是转基因的食品安全问题还有待考证。
利用传统回交技术可以实现对作物高油品种的选育。以花生为例(图1),经鉴定,ahFAD2A和ahFAD2B是与油脂合成有关的基因,选取AABB和aabb基因型的材料作为亲本,其中以AABB为轮回亲本进行回交育种,同时对回交后代株系进行基因型分型,再利用KASP分型鉴定,获得aabb基因型的单株,最后自交得到aabb基因型的纯合株系[9],利用此方法同样可以将其他油脂合成相关基因导入到目标品种中进行作物油脂的改良。
通过诱变育种可以获得与油脂合成相关的突变体。中国育种家已经通过诱变(辐射、化学)途径创制出新的富含油酸的花生突变体[10-11];除此之外,王传堂等[12]利用花生栽培种和野生种Arachisrigonii杂交,在杂交后代中鉴定出与之前FAD2BA插入型突变不同的FAD2G新插入型突变,丰富了花生FAD插入型突变的类型,为油用花生品种培育奠定了基础。

图1 高油酸花生育种流程 Fig.1 The breeding pathway of high oleic acid peanut
利用基因编辑技术,可以将与油脂合成有关的基因在目标作物中进行基因敲入或基因敲除,以达到油脂改良目的。目前,利用CRISPR/Cas9技术已经在水稻、花生、油菜、拟南芥、杨树、大豆、高粱等多个作物中实现了对目标基因的编辑[13-14]。基因编辑育种,首先利用相关载体将目标基因转入需要改良的作物中,然后对作物进行突变位点检测,自交或回交后进行等位基因位点遗传分析,直至得到纯合位点且目标性状能稳定遗传的植株。对油脂合成途径中关键基因的研究进展进行归纳,可为今后利用基因编辑技术培育作物高油品种提供重要的理论基础和现实意义。
3 作物高油品种选育的策略
作物高油品种选育主要通过以下3个方面实现。一是通过调节代谢过程中的碳源分布,使碳源更多地流向油脂合成途径,实现作物高油品种选育。早在1950年前后就有报道,种子蛋白质含量与油脂含量呈显著负相关[15]。日本学者1993年发现,大豆蛋白质含量与磷酸烯醇式丙酮酸羧化酶(PEPCase)活性紧密相关[16]。随后有学者提出“底物竞争”假说,即油脂和蛋白质均来自于葡萄糖的降解产物丙酮酸,即蛋白质合成途径和油脂合成途径存在底物竞争。这一过程主要涉及2类关键酶,即PEPCase和ACCsae的活性。但丙酮酸和磷酸烯醇式丙酮酸可以相互转变,磷酸烯醇式丙酮酸在丙酮酸激酶的作用下转变为丙酮酸;丙酮酸可以在丙酮酸羧化酶的作用下转变为草酰乙酸,然后草酰乙酸在磷酸烯醇式丙酮酸激酶的作用下生成磷酸烯醇式丙酮酸。PEPCase催化磷酸烯醇式丙酮酸合成草酰乙酸进入蛋白质合成代谢,而ACCase催化丙酮酸合成乙酰-CoA进入脂肪酸合成代谢。PEPCase和ACCase的活性分别是调控蛋白质、油脂合成的关键。二是通过调控脂肪酸从头合成关键酶选育作物高油品种。脂肪酸主要的合成场所是质体,脂肪酸的长度和饱和度都有所不同,但是最开始合成的都是饱和脂肪酸(C16和C18),这一最开始合成饱和脂肪酸的过程称为脂肪酸的从头合成[17]。油脂的主要成分是三酰甘油,而合成原料乙酰-CoA首先被运送到质体中在脂肪酸合酶(FAS)的催化作用下经过一系列反应合成脂肪酸,然后被运送到内质网上,与3-磷酸甘油(P3G)结合形成三酰甘油。在脂肪酸从头合成过程中,一系列相关基因已经被克隆和验证,这些基因的变异都会影响最终的油脂含量。三是通过调整三酰甘油的合成效率实现作物高油品种选育。游离脂肪酸在酯酰-CoA合成酶(Long-chain acyl-CoA synthetase,LACS)催化下合成酯酰-CoA,然后被送到内质网上时合成三酰甘油。三酰甘油组装的关键限速酶包括3-磷酸甘油酯酰转移酶(GPAT)和DGAT。在有些作物中,脂肪酸需要先合成磷脂酰胆碱,然后脂肪酸脱饱和,产生油酸和三酰甘油[17]。这一途径中,被鉴定较多的是GPAT、DGAT、PDAT等酶的相关基因。
植物油脂合成调控过程复杂,相关的基因和酶较多。这些基因和酶受到调控都有可能导致油脂合成受到影响。因此,可以通过基因编辑技术或者其他途径来调控这些相关基因,实现作物高油品种的选育。另外,通过分子标记辅助育种或基因编辑技术可以将与植物油脂合成有关的基因导入到目标作物中,使目标基因能够在作物中稳定表达,从而实现作物高油品种的选育。因此,对油脂合成途径关键步骤中的相关基因进行归纳,通过对这些基因的表达进行调控,实现作物高油品种的选育。
3.1 通过调控碳源流向来培育高油品种
3.1.1 PEPCase调控途径 由于PEPCase催化底物丙酮酸进入蛋白质代谢途径,高活性的PEPCase会促使更多的碳源流向蛋白质合成代谢,通过抑制PEPCase活性可提高种子油脂含量。在油菜中抑制PEPCase表达,种子含油量提高15%左右[15]。在陆地棉中,克隆了GhPEPC1和GhPEPC2基因,经验证,这2个基因都与棉纤维伸长有关[18]。ZHAO等[19]通过RNAi技术抑制GhPEPC2基因的表达,在转化植株中幼苗胚芽中的PEPCase活性显著下降,种子中PEPCase活性下降7.3%,总蛋白质含量下降5.65%,而其他农艺性状几乎不受影响;进一步对转化植株进行转录组学水平分析,发现转化植株中与油脂合成有关的基因表达均上调,而与氨基酸合成有关的基因表达均下调,并且转化植株耐盐性显著增强。通过对GhPEPC1基因进行沉默,植物表型没有明显变化,但是相比于对照组含油量增加16.7%。通过转录组学分析发现,GhPEPC1基因表达量降低导致与三酰甘油有关的基因表达量升高(图2)[18]。

图2 碳源流向示意图Fig.2 The schematic diagram of carbon source flow
在棉花品种培育工作中,可以通过筛选GhPEPC1基因表达量低的品种作为种质资源,利用传统育种培育相关品种,也可以通过基因编辑技术抑制GhPEPC1基因的表达来实现棉花高油品种的选育。
在作物高油品种选育中,对油脂含量、PEPcase活性、蛋白质含量进行分析发现,PEPCase活性与最终形成的油脂含量呈显著负相关(图3C),蛋白质含量与油脂含量也呈负相关(图3A);随着PEPCase活性的升高,蛋白质含量逐渐增加(图3B)[20]。
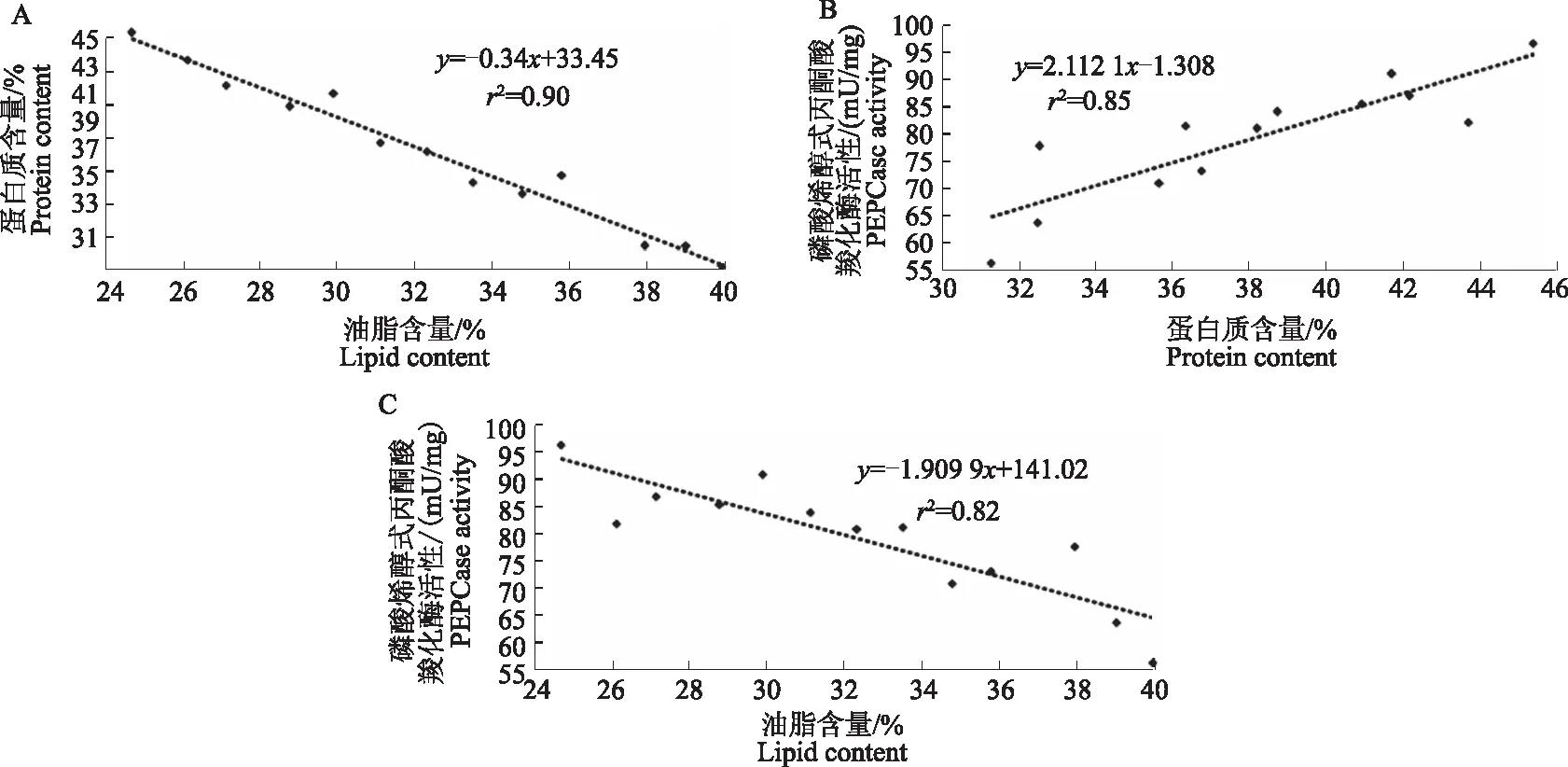
图3 磷酸烯醇式丙酮酸羧化酶活性和油脂、蛋白质含量之间的关系Fig.3 The relationship of PEPCase activity,protein content and lipid content
在海藻中通过敲除CrPEPC1基因发现,三酰甘油的表达量提高了20%,但使PEPCase活性降低了39%~50%。敲除CrPEPC1基因后,与三酰甘油合成有关的基因表达量均升高,相反地,CrPEPC1基因超表达,三酰甘油含量降低37%,PEPCase活性增加了157%~184%。在花生中通过降低AhPEPC1基因的表达量,其油脂含量增加了5.7%~10.3%;转化植株表现出更长的根系,对盐胁迫的抵抗能力也增强,因此,抑制PEPCase基因的表达是育种工作中培育作物高油品种切实可行的方案[21]。
在拟南芥和油菜中,TmDGAT1基因超表达,其含油量均有显著升高,干质量最多增加10%,油脂净含量最高提高30%。将TmDGTA1基因转入相应突变体中,含油量提高20%~50%。相关研究表明,对丝氨酸/苏氨酸位点修饰可以提高DGTA1基因的表达活性,因此可通过此方法来提高含油量[22]。
对明显低含油量的拟南芥突变体lo15571的角果和成熟种子成分进行分析发现,与野生型相比,其光合产物向脂肪酸的转化效率降低;在lo15571角果发育过程中At1g01050表达水平增强,At1g01050基因编码1个可溶性的胞质焦磷酸酶,它是淀粉、可溶性糖向油脂转化的调控基因,通过抑制At1g1050基因的表达,可引起了种子填充过程中焦磷酸的表达进而使种子含油量提高1%~4%[20]。
3.1.2 丙酮酸脱氢酶复合体调控途径 丙酮酸脱氢酶复合体(Pyruvate dehydrogenase complex,PDC)由丙酮酸脱氢酶(E1)、二氢硫辛酸乙酰转移酶(E2)和二氢硫辛酸脱氢酶(E3)3种酶组成,E2是核心催化亚基[23]。PDC和PEPCase的底物分别是丙酮酸和磷酸烯醇式丙酮酸,丙酮酸和磷酸烯醇式丙酮酸可以相互转化。因此,PDC和PEPCase之间存在竞争关系。
通过调控PDC 3个亚基的表达水平可以提高植物种子的脂肪酸含量。植物线粒体丙酮酸脱氢酶复合体(mtPDC)中E2亚基由mtE2-1、mtE2-2、mtE2-33个基因编码,但具体哪个基因对于mtE2影响最大不得而知。通过荧光定量PCR发现,敲除mtE2-1基因,PDC表达量仅为野生型中的17%,敲除mtE2-2后仅对植物成长有轻微的影响,而敲出了mtE2-3对植株没有任何影响[24]。即mtE2-1是mtPDC基因的核心组成部分,而mtE2-2、mtE2-3则或多或少对于mtE2-1存在于功能冗余。拟南芥突变体m132是由mtE2-1突变导致的,这个突变体由于mtE2-1基因的表达导致积累了过量的三羧酸循环的中间产物和一些用于蛋白质合成的氨基酸,说明了mtE2-1是mtPDC的核心组成部分[25]。分析发现,在所有植物器官中都能检测到mtPDC3个亚基的转录产物,但mtE2-2的表达量明显高于另外2个亚基的转录产物,但3个亚基基因表达水平接近,且它们之间的表达量可以互相影响[26]。
PDC催化丙酮酸合成乙酰-CoA,乙酰-CoA是三羧酸循环的物质之一,也是脂肪酸合成的前体物质。三羧酸循环途径所提供的能量,也可以用于脂肪酸合成。在线粒体和叶绿体中都检测到PDC,但活性不同。
3.2 通过调控脂肪酸从头合成关键酶选育作物高油品种
ACCase是催化乙酰-CoA到丙二酰-CoA的关键酶,同时也是限制油脂合成途径的关键酶。ACCase可以与PEPCase形成竞争关系,在作物高油品种选育过程中应尽可能地减少碳源流向蛋白质合成途径,促使更多的碳源流向脂肪酸合成的途径。通过对ACCase的调节可以实现对种子含油量的调控。作物中种子脂肪酸的合成和积累与ACCase的表达有关。ACCase由生物素羧基载体蛋白(Biotin carboxyl carrier protein,BCCP)、生物素羧化酶(Biotin carboxylase,BC)、羧基转移酶α亚基(α-subunit of carboxyltransferase,CTα)、羧基转移酶β亚基(β-subunit of carboxyltransferase,CTβ) 4个亚基组成。有关研究中,4个亚基编码的异质型ACCase被扩增,7、16、10、1个外显子分别从GhBCCP1、GhBC1、GhCTα2、GhCTβ基因中分离鉴定。这4个亚基的转录产物在所有器官中普遍表达,通过分析发现,4个亚基表达量与最终种子含油量之间存在正相关关系;GhBCCP1基因超表达,植株含油量相对于对照组显著增加21.92%;GhBC1和GhCTβ基因超表达,植株含油量较对照组均有约17%的提高[27]。
ACCase活性高的植物脂肪酸含量一般较高,因此可以通过调节ACCase活性来提高种子的最终含油量。BI等[28]发现,在大豆种子脂肪酸积累的早期到中期,高油大豆中ACCase活性显著高于低油大豆。将油菜种子特异性表达启动子和拟南芥同质性ACCase融合,在大豆转移肽的转运下,将ACCase导入油菜中,转化后植株ACCase活性比对照提高了20倍,最终种子脂肪酸含量增加5%[29]。
调控β-酮脂酰-ACP合酶(Ketoacyl-ACP synthase,KAS)相关基因的表达可以培育作物高油品种。作物中KAS由3类酶构成,分别为KASⅠ、KASⅡ、KASⅢ。KASⅠ催化4∶0-ACP 延长碳链到16∶0-ACP,KASⅡ催化16∶0-ACP延长碳链到18∶0-ACP,KASⅢ主要催化乙酰-CoA与丙二酰-CoA合成4∶0-ACP[1]。将具有合成90%中链脂肪酸功能的油菜基因转入到油菜品种Cuphea中,结果发现,野生型的隐形基因和包含了单点突变的FATB基因的2种不同单一结构参与种子KASⅢ的合成,沉默BnKASⅢ、BnFATB基因的T2株系中各种脂肪酸的含量都有提高,因此,可以通过沉默2个基因实现高油油菜品种的选育[30]。在麻风树中克隆了JcKASⅡ基因,JcKASⅡ的转录产物在所有组织中均被检测到,种子萌发过程中JcKASⅡ表达量增加,fab1是拟南芥中AtKASⅡ突变体,含有较少的C18和较多的C16,通过转入JcKASⅡ基因发现,该突变体性状得到恢复;JcKASⅡ基因超表达会促使种子和叶片中C16脂肪酸向C18脂肪酸转变,说明JcKASⅡ基因能够促使18∶0-ACP的积累[31]。不同作物品种中KAS基因的表达不同,含油量也不尽相同,根据选育作物的不同目标,调控KAS基因的表达可以实现作物高油品种的选育。
调控酰基ACP硫酯酶(acyl-ACP thioesterase,FAT)基因的表达可以实现作物高油品种的选育。对FAT调控可以实现对植物油脂品质的调节。FAT作为脂肪酸合成最后一步的催化酶,决定了合成脂肪酸的类别以及链长,其中,一种方式是FAT与ACP合成的酯酰基结合然后再与辅酶A结合,另一种方式是直接由ACP释放酯酰基,形成游离的脂肪酸。植株中存在2种FAT(FATA和FATB),与它们结合的底物具有碳链长度的特异性[32]。FATA参与C18∶1-ACP和C18∶0-ACP硫酯键的形成,同时,对于C18∶0-ACP的调节活性较强而对C18∶1-ACP的调节活性相对较弱;FATB参与饱和酯酰碳链的释放以及非饱和酯酰碳链的合成,如16∶0-ACP硫酯酶的形成和释放[33]。培育作物高油品种对油脂品质的要求是尽可能多地提高不饱和脂肪酸比例,因此,可以通过调控FAT相关基因的表达实现作物高油品种的选育工作。
作物高油品种选育可以通过延长脂肪酸的碳链来实现。脂肪酸主要存在于质体中,但目前转化植株大多数都是基于核转化,相对于质体转化,有着表达水平高并且可以同时进行多基因转化等优点。因此,通过基因编辑技术调控脂肪酸从头合成基因的表达是培育作物高油品种的一条途径。
3.3 通过调整三酰甘油的合成效率实现作物高油品种选育
影响三酰甘油合成效率的关键酶主要有GPAT、DGAT和PDAT等,作物高油品种的选育可通过调控这些关键酶来实现。
作物高油品种的培育可以通过提高GPAT基因的表达来实现。GPAT催化三酰甘油组装的第1步反应。拟南芥敲除GPAT9(At5g60620)的突变体显示,雄性和雌性配子体致死表型一致,显示该基因有重要的膜脂合成作用,说明其参与了三酰甘油油脂的合成。拟南芥中AtGPAT9的超表达转化植株,种子含油量增加,而抑制AtGPAT9基因表达,种子含油量减少。可见,调控GPAT相关基因的表达是培育高油品种的策略之一[34]。
通过调控DGAT相关基因的表达可以实现作物高油品种的培育,DGAT催化二酰甘油与脂肪酸酰基结合成三酰甘油,是作物油脂合成的关键限速酶。植物主要有DGAT1、DGAT2、DGAT3等类型DGAT相关基因,它们都参与了油脂的合成。在拟南芥中,DGAT3在种子发育及育苗过程中起作用[35-36]。在不同物种中,DGAT2对于种子含油量的积累都有着重要作用,乌桕SsDGAT2基因在拟南芥中超表达,种子含油量较对照显著增加50%,但种子含油量增加的同时亚麻酸含量降低。因此,基于SsDGAT2调控对于乌桕高油品种的改良需要综合考虑含油量和亚麻酸含量的关系,以培育含油量高且亚麻酸含量适中的品种[37]。DGAT1基因也是油脂合成的关键基因,影响植物种子含油量积累和种子的油脂组成。乌桕SsDGAT1基因在拟南芥中超表达,种子含油量较野生型增加12.3%~14.7%,但在油菜中超表达,转化植株种子含油量相较于野生型明显下降且亚麻酸含量也出现下降趋势,说明SsDGAT1参与植物油脂合成,对于今后培育高油乌桕品种具有重要理论意义[38]。
作物高油品种选育可以通过调节PDAT来实现。PDAT是相对于DGAT的另一类二酰甘油转移酶,与DGAT功能类似,催化磷脂和二酰甘油生成溶血磷脂和三酰甘油。亚麻中PDAT基因过量表达会导致不饱和脂肪酸18∶3n-3向18∶2n-6的转变,但对总含油量没有影响。沉默PDAT基因能得到更高的不饱和脂肪酸(18∶3n-3)含量[39]。在亚麻品种中鉴定了5个PDAT基因(CsPDAT1-A、CsPDAT1-B、CsPDAT1-C、CsPDAT2-A、CsPDAT2B),通过相关试验发现,CsPDATs基因在增加种子含油量和提高作物抗逆性方面起重要作用[40]。
研究发现,PDAT1基因能够补充由于DGAT1基因缺失而减少的种子含油量,改变了子叶中的脂肪酸组成,但最终种子含油量不受影响[39]。因此,在通过调控三酰甘油合成效率选育作物高油品种时应综合考虑PDAT、DGAT基因的表达。
4 小结与展望
综上,总结了近几年关于作物油脂合成的研究进展,归纳了作物高油脂品种选育的策略,并从碳源流向、脂肪酸从头合成、三酰甘油的合成效率三方面详细综述了培育作物高油品种的方法策略,为作物高油品种和特殊脂肪酸品种的选育提供了理论基础。随着社会的发展和人类的进步,不可再生资源减少日趋严峻,而植物柴油的开发是解决未来能源问题的一个很有希望的方向,培育作物高油品种是解决自然资源短缺的手段之一;同时,不饱和脂肪酸、亚麻酸含量高的植物对人体健康有益,这也是作物育种工作未来的方向之一。
猜你喜欢
工业微生物(2024年1期)2024-02-29 07:36:50
生物加工过程(2023年6期)2023-12-11 03:27:52
农业科技通讯(2023年1期)2023-02-12 07:10:04
牡丹江医学院学报(2021年5期)2021-12-05 08:01:51
农民致富之友(2019年18期)2019-07-31 01:32:15
发明与创新(2019年9期)2019-03-26 02:22:48
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:04:59
科学种养(2014年4期)2014-06-09 03:09:19
作物研究(2014年6期)2014-03-01 03:39:12
中国粮油学报(2013年2期)2013-09-17 02:30:38
