
乌梁素海叶绿素a的时空分布及其与环境因子的关系
2019-06-18 10:59蒋鑫艳李畅游史小红孙标赵胜男孙驰
生态环境学报 2019年5期
蒋鑫艳,李畅游,史小红,孙标,赵胜男,孙驰
内蒙古农业大学,内蒙古 呼和浩特 010018
浮游植物在湖泊生态系统中起着能量转换与物质循环的作用(秦伯强等,2011;刘波等,2008;吕唤春等,2003),其中,藻类植物通过叶绿素吸收和传递光能进行光合作用。叶绿素a(Chl.a)为所有浮游植物均具有的光合色素,常被作为表征水环境质量的水生生物指标之一,其含量可以体现及评价水体富营养化状况,水域内源污染、浮游藻类(光能自养浮游植物)过量繁殖必须参考这一参数(Reynolds,1984;Salmaso et al.,2006;Kruskopf et al.,2006;Katsiapi et al.,2016)。富营养化一直以来是国内外湖泊面临的主要水环境问题之一(雒文生等,2009),尽管近年来乌梁素海水质稍有改善(李红悦等,2018),但仍不容小觑,其水体质量直接影响着当地的水产养殖经济和黑鹳等珍稀鸟类的生存环境;又因该湖区具有缓冲和自然净化作用致其出水影响着黄河水质,是黄河中上游重要的调水、蓄水、保水场地,其重要性不言而喻,故研究其污染现状具有实际指导意义。
乌梁素海是蒙新高原湖泊中重要的科研对象之一,有关该水域Chl.a开展的研究大多集中在非冰封期,而结合冬季冰封期分析 Chl.a含量的分布特征及其与环境因子的相互关系尚不多见。如张晓晶等(2010)发现乌梁素海2008年非冰封期Chl.a含量呈夏季>秋季>春季,空间分布呈东北高西南低;相关分析表明Chl.a与生化需氧量、悬浮物、浊度、总磷呈极显著相关,与透明度呈极显著负相关;综合逐步回归方程表明其主要影响因子为浊度、悬浮物、总磷、总氮、硝酸盐氮和亚硝酸盐氮。冯伟莹等(2013)采用 2006-2010年非冰封期 5-10月乌梁素海监测数据,对Chl.a含量的时空分布及其与总氮、总磷的相关关系进行分析,发现乌梁素海Chl.a含量呈秋季≈春季>夏季,空间分布呈北部高于南部,且不同时期 Chl.a含量与氮磷相关性不同。毛旭锋等(2015)发现乌梁素海 2013年Chl.a的含量呈7月>9月>5月,这主要与该地区气候有关,空间上Chl.a浓度通常在入湖处最高,出水口最低,其含量变化受多种因子共同影响。
乌梁素海作为中国北方寒旱区典型湖泊的代表,地处中国北方高纬度地区。与南方湖泊相比,除自然地理特征具有显著差异外,该地区受北温带大陆性干旱半干旱气候影响,四季更迭明显,冬季寒冷干燥、历时长。如太湖常年水温保持在5 ℃以上,冬季温和湿润,湖面无结冰现象(王震等,2014),而乌梁素海于每年10月下旬会进入长达5个月的冰封期,水温低至-0.3 ℃,冰层厚度可高达水深的 2/3,阻挡了大气与水体的物质交换。加之冰封期湖水的结冰与消融过程会对冰下水体的营养盐分布产生间接作用(张岩等,2011),导致在冰层环境和冰下水层中出现污染物迁移的现象,进而影响湖泊冰封期的生态系统,使得乌梁素海冰封期水体理化指标有别于同时期南方湖泊,仅研究非冰封期乌梁素海水质现状不能代表其整体情况,故对该湖区冰封时期水体进行研究有一定的必要性和实际价值。
本研究于 2014-2017年连续采集乌梁素海春夏秋三季的表层水体及冰封期冬季的冰下水体,分析 Chl.a含量的时空分布及其与环境因子之间的关系,旨在了解生态恢复前后的4年间乌梁素海藻类生物量变化及其影响因素,为寒旱灌区草型浅水湖泊的污染防治提供理论参考。
1 材料与方法
1.1 研究区概况
乌梁素海,中国第八大淡水湖泊,地理坐标40°36′-41°03′N,108°43′-108°57′E,是地球同一纬度的最大湿地,黄河流域最大的淡水湖泊,同时也是全球荒漠半荒漠地区极为少见的大型多功能草藻型湖泊(王鑫磊,2012)。湖区库容量为2.5×108-3.0×108m3,水域整体面积约为305.97 km2,2017年实测平均水深 1.5 m。乌梁素海地处河套灌区,其所在流域太阳辐射强度大、流域内降雨少而蒸发大。四季更替明显、气温变化差异大,多年平均气温为5.2 ℃,每年的1月份为当地最冷月,平均气温在-11-15 ℃之间,全年无霜期为 152 d。湖面开始出现结冰封冻现象大约在每年 11月,次年 1月下旬冰体厚度达到最大,3月冰面开始逐渐消融,3月下旬完全解冻融化。
1.2 水样采集与指标测定
选取19个采样点,采样位置见图1,图中总排干、八排干和九排干渠道是乌梁素海主要入湖河流。采样时间:2014-2017年的春季(5月)、夏季(6-8月)、秋季(9-11月)、冬季(次年1月)。采样频率为每月中下旬。采样方式为船只采样,采样垂向位置在水面以下0.5 m处,其中,冬季样品为冰下水体。

图1 监测点位布设Fig. 1 Location of field observation sites
样品监测项目主要包括叶绿素a(Chl.a)、总磷(TP)、溶解无机磷(DIP)、正磷酸盐(DTP)、总氮(TN)、悬浮物(SS)、水温(WT)、电导率(EC)、溶解氧(DO)、总溶解性固体(TDS)、盐度(Sal)、透明度(SD)、冰层厚度(IT);其中,Chl.a的测定采用丙酮萃取分光光度法,TP、DIP、DTP采用钼酸铵分光光度法(GB11893-89),TN采用碱性过硫酸钾消解紫外分光光度法(GB11893-89),SS采用重量法(GB11901-89),其余为现场可测试指标。
1.3 数据统计与分析方法
利用Excel 2010办公软件进行数据统计与处理;利用ArcGIS 10.5中反距离权重法将数学函数与制定变量的点进行拟合,绘制氮磷及Chl.a空间分布图;利用SPSS 19.0分析Chl.a和理化环境因子的相关性并建立以Chl.a为因变量的逐步回归方程。
2 结果与分析
2.1 乌梁素海Chl.a含量时间分布
对2014-2017年各季节Chl.a含量取均值分析其季节分布,结果如图2所示,2014年、2015年、2017年冬季冰封期Chl.a含量最高,非冰封期夏秋两季次之,春季Chl.a含量最低。2016年春夏秋冬四季Chl.a质量浓度均值分别为7.672、4.897、4.408、6.869 μg·L-1,全年Chl.a含量处于较低水平,变化趋势较其他年份更平稳,表现为春季最高,冬季冰封期稍次之,这与其他年份所表现出的规律略有不同。此外,2014-2016年,夏季Chl.a含量大于秋季,2017年则反之。综上,乌梁素海Chl.a时间分布大致呈冬季冰封期高于非冰封期各季节。
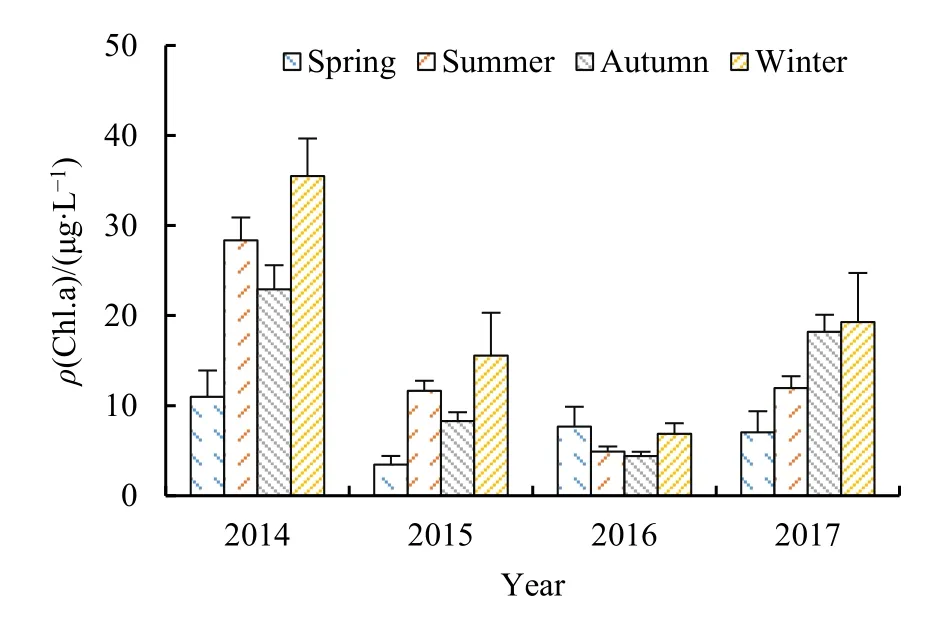
图2 2014-2017年Chl.a浓度季节变化Fig. 2 Seasonal variation of Chl.a concentration in 2014-2017
取各年 Chl.a浓度均值分析其年际变化趋势及年内变化范围,结果见表 1,2014-2017年各年Chl.a质量浓度均值分别为25.157、9.846、5.270、14.521 μg·L-1,2014-2016 年 Chl.a 含量呈逐年递减趋势,但2017年又稍有回升。其中,2014年最高、最低、平均 Chl.a含量均高于其他年份,2015年、2017年Chl.a含量变化范围基本一致,2016年Chl.a含量(30.917 μg·L-1)峰值远低于其他年份。

表1 2014-2017年Chl.a浓度年际变化Table 1 Annual variation of Chl.a concentration in 2014-2017
2.2 乌梁素海Chl.a含量空间分布
以经度 40°57′30″和 40°53′30″为界限,将乌梁素海分为湖北区、湖心区和湖南区三部分,具体划分见图1。湖北区各测点(J11等)处于排干入湖处,灌区90%以上的补给水源经总排干汇入(杨志岩等,2009a;杨志岩等,2009b),主要承接上游黄河补水、生活污水和农田退水,其中农田灌溉退水携带着大量的化肥、农药等污染物,以致该区水体透明度较差,氮磷浓度高,富营养化程度严重;湖心区包括芦苇区、沉水植物密集区及其周围的明水域,水体较澄清;湖南区水质逐渐转良,仅有极少量水体排入,其出水最终汇入黄河,从而影响黄河水质。
对不同湖区各季节 Chl.a含量取均值分析其空间维度变化,图3结果表明:2014年(除秋季外)、2015年、2017年夏秋冬三季Chl.a含量从湖北到湖南逐渐降低,春季为湖北区>湖南区>湖心区;2016年湖心区与湖南区Chl.a含量空间变化差异不大,但同年夏秋两季Chl.a含量呈湖北区略低于湖南区。此外,除2016年秋季外,2014-2017年各季节Chl.a浓度峰值均出现于湖北区监测点。综上,湖北区Chl.a含量对全湖区贡献最大,湖心区次之,湖南区较低。
2.3 冰封期乌梁素海Chl.a与TN、TP含量的空间变化及冰层厚度分布
通过反距离权重法IDW 拟合插值,对乌梁素海冬季冰封期Chl.a、TN、TP进行空间变异性分析,以了解乌梁素海冰封期富营养化现状和变化趋势。图4结果表明:2014-2017年冰封期Chl.a含量空间分布呈现出由湖北区向湖南区逐渐递减的现象,这与图5、图6所示同时期氮磷营养盐空间分布规律相似,可见Chl.a含量空间分布一定程度上受TN、TP含量的控制。

图3 2014-2017年Chl.a质量浓度空间变化Fig. 3 Spatial variation of Chl.a concentration from 2014 to 2017

图4 2014-2017年冰封期Chl.a的IDW插值图Fig. 4 IDW interpolation of Chl.a concentration distribution during ice-sealed period in 2014-2017
乌梁素海是蒙新高原典型的寒旱区湖泊,2014-2017年冬季冰封期的冰层厚度平均值为 0.449 m,冰层厚度范围为0.249-0.73 m,冰下水体平均水温0.93 ℃,水温范围为-0.30-3.70 ℃。冰层厚度通过对氮磷等养分产生作用而间接影响浮游植物生长及Chl.a水平。图7所示为冰封期冰层厚度分布:时间上,冰层厚度受当年冬季气温影响,2014-2016年逐年下降,2017年又大幅回升;空间上,冰层厚度分布呈湖北区>湖心区>湖南区。结合图2、图 4、图 7,冰层厚度时空分布规律与冬季冰封期Chl.a含量时空分布具有高度一致性。
2.4 乌梁素海Chl.a含量与环境因子的关系
受水生态系统本身复杂性的影响,水体中各理化因子会对 Chl.a含量产生直接或间接的作用,且不同理化因子的作用程度也不尽相同;与此同时,Chl.a含量的变化也会对环境因子产生一定反作用。故利用SPSS统计分析软件对Chl.a与有关环境指标进行相关性分析和逐步回归分析。其中,冬季冰封期 Chl.a含量时空分布与冰层厚度时空分布规律相一致,故考虑在冬季引入冰层厚度指标进行分析。

图5 2014-2017年冰封期TN的IDW插值图Fig. 5 IDW interpolation of TN concentration distribution during ice-sealed period in 2014-2017
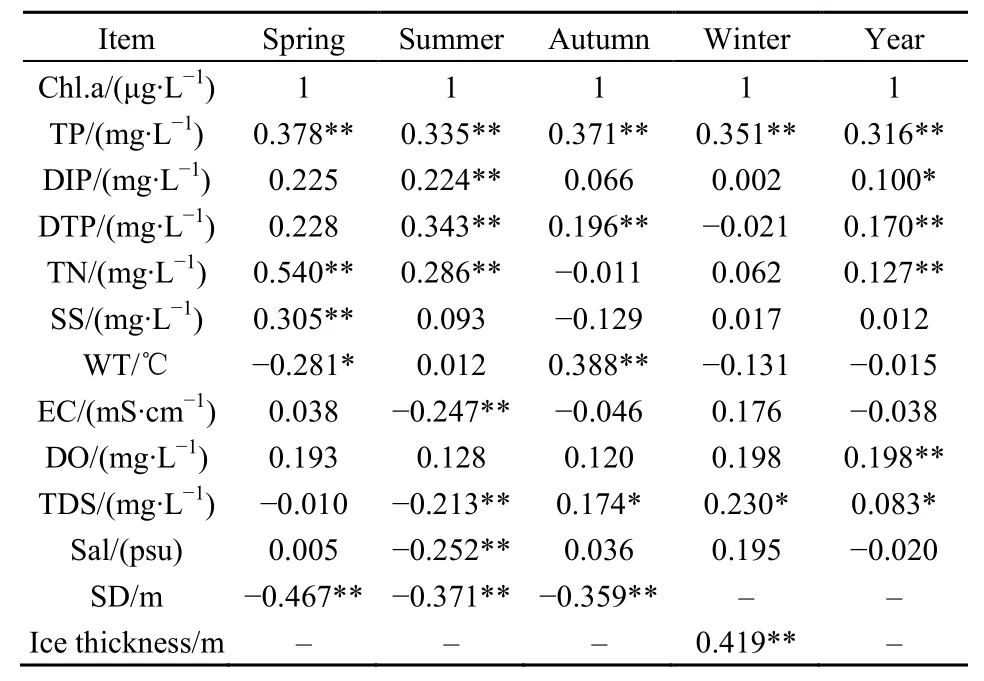
表2 Chl.a与环境因子的相关性分析Table 2 Correlation analysis of Chl.a and the related environmental factor
对四季及多年 Chl.a和各项环境指标进行相关性分析,可以揭示指标间共同变化的一致性程度。由表2相关性分析结果,在非冰封期春季,Chl.a与TP、TN、SS呈极显著正相关,与SD呈极显著负相关;在夏季,Chl.a与TP、DIP、DTP、TN呈极显著正相关,与EC、TDS、Sal、SD呈极显著负相关,可见夏季浮游植物的生长受多项环境理化因子共同作用;在秋季,Chl.a与TP、DTP、WT呈极显著正相关,与TDS呈显著正相关,与SD呈极显著负相关;在冬季冰封期,Chl.a与 TP、冰层厚度呈极显著正相关,与TDS呈显著正相关;多年Chl.a与营养盐TP、DIP、DTP、TN呈不同程度的相关性,且与TP的相关性最为显著,同时与TDS、DO也有显著的正相关性。如图8所示,2014-2017年冬季冰下水体中TDS、DO均大于其他季节。
对 Chl.a和各项环境指标进行多元线性回归分析,通过逐步筛选建立函数控制关系,以期明确各回归方程的共同环境理化因子有SD和DO;冰封期冬季有冰层厚度、EC及营养盐TN、TP、DTP入选回归方程;建立的多年回归方程只有TP和DO入选。

图6 2014-2017年冰封期TP的IDW插值图Fig. 6 IDW interpolation of TP concentration distribution during ice-sealed period in 2014-2017

图7 2014-2017年冰封期冰层厚度分布图Fig. 7 Distribution of ice thickness during ice-sealed period in 2014-2017
3 讨论
3.1 乌梁素海Chl.a含量时间分布

图8 2014-2017年TDS、DO浓度季节变化Fig. 8 Seasonal variation of TDS and DO concentrations in 2014-2017

表3 Chl.a与环境因子的逐步回归方程Table 3 The stepwise regression equation of Chl.a and the related environmental factors
2014年、2015年和2017年乌梁素海Chl.a含量的季节变化表现为冬季冰封期>夏秋两季>春季。这与南方湖泊研究有所差异:如王震等(2014)发现太湖 Chl.a含量季节分布规律总体呈现夏季>秋冬两季>春季;张光贵(2016)发现洞庭湖Chl.a含量总体呈现出夏季>秋季>冬季>春季的分布。这主要是由于河套灌区气候特征、湖水自身水质、植物生长周期、以及非冰封期农田浇灌退水等综合因素导致。春夏秋三季分别是浮游植物的生长期、旺盛期和成熟衰亡期,Chl.a含量的变动大致遵循其生长周期规律。内蒙古春季因多风沙而素有“风季”之称,多风天气使得该浅水湖受到频繁交换与扰动,给浮游植物生长带来不利影响。此外,河套灌区春浇的农田退水虽携带大量氮磷等营养元素进入湖体,但因浮游植物处于生长适应期,对营养元素的需求利用处于较低水平,故表征藻类数量的 Chl.a含量低于其他季节。夏季雨水充沛、温度适宜、日照时间长,处于生长旺盛期的浮游生物充分利用了春浇滞留的营养元素,数量大幅增加;进入秋季,随着温度的降低,浮游植物进入成熟期开始衰亡,数量减少,但2017年出现秋季Chl.a含量略大于夏季的现象,可能与该年秋季氮磷入湖量有关,这也正体现了湖泊水生态的复杂性。冬季冰封期乌梁素海 Chl.a含量高于其他时期,其一是由于秋季浮游植物的衰老引起对养分吸收的滞后,与此同时,大型水生植物的枯萎凋亡加剧了冬季水体中的氮磷等营养盐含量的升高;其二,结冰封冻是导致当年冰封期Chl.a含量大于非冰封期的重要原因。此外,随着次年春季天气回暖,冰体逐渐解冻消融,其融水会对营养盐及Chl.a含量起到一定的稀释作用,使冬季Chl.a含量仍处于较高水平。至于2016年春季 Chl.a含量稍高于冬季冰封期,可能是因为冬季采样期间,受到当时气温影响,冰体仍处于生长初期阶段,冰层厚度并未达到峰值,TN、TP和Chl.a等营养盐的迁移排出受到暂时性抑制。综上,近 4年来,冬季乌梁素海水体更易发生富营养化。
以水环境指示性指标 Chl.a为媒介识别乌梁素海水环境的季节变化状况,发现治理期间 2014-2016年叶绿素a呈逐年递减趋势,但2017年又略有回升,说明对湖内围网养殖的控制起到了作用,该水域的生态治理修复工作取得了一定成效,故应继续合理管控渔业养殖作业。基于冰封期水体富营养化总体上较其他时期严重这一现象,可以说明冰封期不失为更合适的治理期,故建议水环境保护工作者对冬季的水环境污染防治加以重视,可以考虑在封冻前打捞衰败的草藻,封冻后适当加大生态补给水量。
3.2 乌梁素海Chl.a含量空间分布
藻类细胞通过获取和储存栖息地底部或游离态的养分供自身在水体中生长所需(Grover,2017),在其生长环境条件得到一定程度满足时,氮磷等营养盐是否充沛就成为种群生长繁殖和数量多寡的主要控制因素。种群随机运动与营养盐的扩散速率相同,使得水体中氮磷等营养物质在不同区域的变动引起以其为食的浮游藻类生长繁殖出现相应变动。
湖北区 Chl.a含量对全湖区贡献最大,湖心区次之,湖南区较低。这与以往研究结果相一致,如毛旭锋等(2015)发现乌梁素海 Chl.a浓度空间分布呈由西北向东南方向降低,仅少数监测点位随流动过程逆向升高。在非冰封期(夏秋两季),由于人类生产活动的干预,大量氮磷等营养物质从湖北区的排干进入,途经整个湖区后再到湖区南端的乌毛计排出时,由于沿途有浮游藻类、芦苇等大型水生植物的吸收同化和降解转化,使得夏秋两季乌梁素海Chl.a含量空间分布大体呈湖北区>湖心区>湖南区。冬季冰封期Chl.a含量空间分布与夏秋两季一致,但原因有所不同:冬季人为活动影响减弱,补给水源主要为地下水,其次为污废水,入流量少,造成的扰动小,从而表征藻类数量的 Chl.a出现同于水体中内源性氮磷养分的轨迹变化。
春季Chl.a含量空间分布区别于其他三季,表现为湖北区>湖南区>湖心区。首要影响因素为氮磷营养盐水平,故湖北区Chl.a含量最大。其次是风力作用和水动力条件:春季乌梁素海气候干燥、风沙大,风速表现为湖心区>湖南区>湖北区,直接影响水流速度同步变化;外加乌梁素海春浇时期,湖北区、湖心区承接排干入水,对水体造成的扰动较湖南区强烈。两者同时作用致使湖心区浮游藻类未能及时、充分吸收氮磷营养盐,而湖南区藻类具有相对优势,直接体现为湖南区Chl.a含量大于湖心区。
研究发现,Chl.a峰值及其较高含量几乎全部出现于湖北区,而较低水平的Chl.a含量多分布于湖南区,故乌梁素海是具有明显空间异质性的草藻型湖泊。需要说明的是,总排干入湖口处一些监测点位Chl.a之所以出现异常高含量,是因为乌梁素海位于河套灌区的末端,直接承接上游总排干、八排干渠道来水的外源污染,与湖中心相较而言有着不可忽视的人类生产活动的影响,使得浮游藻类的生长受到一定程度的干扰,最终体现为Chl.a含量的特殊变动。
此外,2014-2017年湖北区Chl.a含量递减趋势明显,可见水域源头治理成效显著,这与近年来相关单位对湖北区入湖处的污染控制有着密不可分的关系。
3.3 冰封期乌梁素海Chl.a含量与冰层厚度的关系
乌梁素海冰封期与非冰封期 Chl.a含量的时空分布有区别于其他湖泊的独特性(田伟东等,2016;黄慧琴等,2016;朱广伟等,2018;钱昊钟等,2013),尤其是冰封时期。乌梁素海冰封期Chl.a含量空间分布与氮磷空间分布特征有很强烈的同步性,由北向南呈现出逐渐降低的趋势,表明Chl.a与氮磷有一定空间相关性。这是由于藻类植物通过吸收同化氮磷等营养盐后开始快速生长(李畅游等,2005;肖博文等,2015;Popova et al.,2010),营养盐的分布会直接影响浮游植物在水体中的种群数量分布(Grover,2017):这是由于蒙新高原冰封期独特的气候原因及风力作用,冰层表面并未有大量积雪覆盖,冰层虽厚但透明而洁净,使得在白天靠近冰层的冰下水体能够接受一定程度和时间的光照,为浮游藻类的生长繁殖创造了光照条件;其次,虽然冬季气温不适宜微生物生长,但厚实的冰层实则为冰下水体的浮游藻类提供了保温屏障,缓解了恶劣天气的影响(韩知明等,2017);再之,氮磷等营养盐在靠近冰层的冰下水体中浓缩富集,给浮游藻类生长繁殖提供了物质基础,加之由芦苇和草藻的残体转化而成的腐殖质在分解时,释放出的养分也可供给浮游藻类吸收利用,释放出的二氧化碳可加强其光合作用。因此,冬季浮游植物并未出现大量衰败消亡之势。加之冬季冰盖层对外界风力作用的阻隔导致水动力减弱,且汇入水量骤减、水体流动缓慢,使得营养盐停留时间长、水质基本处于稳定状态,故Chl.a含量空间分布与营养盐分布一致。
此外,对于冬季冰封期Chl.a高含量集中在湖北区(特别是入湖口)附近的现象,发现冰层厚度空间分布、氮磷营养盐空间分布与Chl.a含量空间分布规律大体一致,即冰层越厚,水体中 Chl.a、TN和TP含量也越高,仅从感官上可以明显看到冰下水体颜色较浑浊,与洁净冰层对比明显。这是缘于冰封期低温冷冻过程会引起冰下水体中的污染物产生浓缩效应,进而出现氮磷营养盐、无机物等由冰层向冰下水体迁移排出的现象。张岩等(2011)从结晶学和热力学角度探讨了湖泊结冰过程中污染物的迁移机理及特征,发现冰体生长期间,伴随着冰层厚度的增加,Chl.a、TN和TP由冰层逐渐向水层迁移,在水体中不断聚集蓄存,使得冰下水层中Chl.a、TN和TP的含量分别是其对应冰层中的1.64、2.06、1.50倍(张岩等,2011)。由表2可知,冰层厚度对 Chl.a含量的贡献最大,其相关系数为0.419,有着间接的正向促进作用,这进一步佐证了冰封期Chl.a含量空间分布与冰层厚度关系密切。
综上,结冰过程会对冰下水体的营养盐分布产生间接作用,即污染物由冰体内部向水体转移,故冰封期水体冻结过程会促使冰下水体的富营养化愈加严重,水质状况恶化。由结冰引起的湖泊水深下降会影响水域环境承载力,进一步加重湖泊冰下水体富营养;湖面覆盖的大量厚而密集的积冰,不仅阻碍了大气复氧,而且抑制了水体流动与交换,进一步加剧了冰下水体水环境的恶化程度。通过对冬季污染物迁移特性的了解,发现乌梁素海冬季富营养化的原因与其他季节有所不同,水域保护工作者在进行治理时应对冰层厚度的影响给予一定重视。
3.4 乌梁素海Chl.a含量与TN、TP等理化环境因子的关系
基于《地表水环境质量标准》(GB3838—2002),发现冰封期TN含量超过Ⅴ类标准;冰封期TP含量在Ⅲ类到Ⅳ类标准之间,且其变幅同于冰封期Chl.a年际变化趋势,表明Chl.a含量受TP影响更显著。
Chl.a与环境因子的相关性分析表明,非冰封期春夏两季Chl.a水平受氮磷营养盐共同作用。其中春季Chl.a含量与表征营养水平的TN的相关性最显著,TP稍次之,即表明春季氮盐对浮游植物生长繁殖的控制更突出。夏季则与其反之,Chl.a水平受磷盐主控;在秋冬两季,Chl.a含量与TP显著相关,与TN无相关性,表明磷盐的影响占主导作用;以多年水平来看,Chl.a含量与TP的相关性最显著,磷盐控制作用更大,但同时发现各年冬季冰下水体TN含量均高于其他季节。冬季TDS含量高表明冰下水体积累了大量溶解态的有机和无机养分。
结合逐步回归方程分析入选因子,非冰封期春夏秋三季,SD与Chl.a呈极显著负相关,相关系数分别为-0.467、-0.371、-0.359,这是因为水体透明度低会使浮游藻类光合作用降低、生长速率减慢,反之,水体透明度亦会因为藻类植物大量繁殖而降低。四季DO与Chl.a呈正相关关系但并不显著,相关系数为0.193、0.128、0.120、0.198,这是因为存在着多种因素综合影响。在春夏秋三季,除了浮游藻类光合作用释放氧气外,表层水体与大气接触,氧气交换迅速使得水汽交接界面的溶解氧处于动态平衡,且水温对溶解氧有一定的调节作用;至于冬季,厚实冰层的覆盖使得自然曝气形成的大气复氧过程受到限制,故DO处于过饱和状态则可能与水体中的氧气在低温条件下溶解度较高以及浮游植物进行的光合作用有关。不同于本研究,张晓晶等(2010)发现 2008年乌梁素海非冰封期所有监测点Chl.a含量与DO呈显著负相关,这可能取决于当时水域环境状况。
4 结论
乌梁素海是时空异质性较大的草藻型富营养化湖泊。时间上,2014年、2015年和2017年冰封期乌梁素海Chl.a含量高于非冰封期,即冬季>夏秋两季>春季,冬季水域更易发生富营养化;2014-2016年Chl.a含量呈逐年递减趋势,2017年又稍有回升,可见水域环境状况整体上有所改善,水体自净能力得到逐步恢复。空间上,夏秋冬三季 Chl.a含量表现为湖北区>湖心区>湖南区,春季为湖北区>湖南区>湖心区。
冰封期乌梁素海Chl.a与TN、TP具有一定的空间相关性。Chl.a含量空间分布受营养盐水平、冰层厚度、水动力学特征等因素综合影响,且湖北区冰层越厚,对Chl.a和氮磷营养盐在冰下水体中的富集效果也越强烈。今后对乌梁素海的研究应加深对冰封期各水质指标变异规律的了解,以期更好地进行该时期水域富营养化防控,实现水环境生态可持续发展。
结合相关性和回归分析,四季Chl.a含量受氮磷营养盐不同程度的控制,总体上多年 Chl.a含量与 TP的相关性最显著,多元回归方程为YChl.a=58.896XTP+0.762XDO+1.873。非冰封期春夏秋三季,入选回归方程的共同环境理化因子有SD和DO;冰封期冬季,冰层厚度对Chl.a含量的影响最显著,其相关系数为0.419。
猜你喜欢
疯狂英语·新悦读(2022年1期)2022-11-22
百科探秘·海底世界(2022年6期)2022-06-24
环境工程技术学报(2022年3期)2022-06-05
疯狂英语·新阅版(2022年1期)2022-01-26
疯狂英语·新读写(2021年8期)2021-11-05
奇闻怪事(2021年2期)2021-04-20
家教世界(2018年16期)2018-06-20
海外星云(2016年7期)2016-12-01
小朋友·聪明学堂(2015年12期)2016-01-07
应用海洋学学报(2015年3期)2015-11-22
