
强烈度火后喀纳斯泰加林草本层物种丰富度和地上生物量变化
2019-06-18 10:59刘小菊潘存德
生态环境学报 2019年5期
刘小菊 ,潘存德
1. 新疆农业大学林学与园艺学院,新疆 乌鲁木齐 830052;2. 新疆农业职业技术学院园林科技学院,新疆 昌吉 831100
林下植被是森林生态系统不可缺少的重要组成部分,在森林生态系统的营养元素的循环、积累和改善林内环境、维持生态系统的多样性、生态系统功能的稳定性及森林的演替等方面具有十分重要的作用(Chastain et al.,2006;Brockerhoff et al.,2008;Liu et al.,2014;Thrippleton et al.,2016)。林下草本层的研究多集中于林下植被多样性和生态结构方面(Ombega et al.,2017;Xu et al.,2017;金艳强等,2017),而对林下植被生物量研究相对较少,林下植被生物量只是森林生态系统的一小部分,但对生态系统的稳定性和多样性起着重要作用(陶冶等,2013)。
随着演替的正向进行,物种多样性因物种组成的改变将发生强烈变化,森林演替顶级群落拥有最大的碳库组成及最稳定的碳库结构(高一飞等,2016)。有研究发现,在森林次生演替过程中,随着林龄的增大,林下乔木和草本层生物量持续增大(史山丹等,2012),因此森林的生物量随着演替的进行持续增长,固碳能力不断增强。冠层结构影响林下植物的生长,也影响林下植物生物量的分配。因此,研究不同森林群落草本层生物量和物种多样性的变化及其与林分因子的关系,对于发挥林下植物的碳汇功能具有重要指导意义。
喀纳斯山地森林和其他北方森林一样,火干扰是构成其森林演替和生生不息的主要源动力(Gromtsev,2002),火干扰对森林群落的物种组成和生物量动态变化起着举足轻重的作用。已有学者对喀纳斯泰加林的火干扰史、火干扰烈度(刘翠玲等,2009)、火干扰与树种结构的关系(刘翠玲,2009)、火干扰烈度与植物种生态位关系(刘景等,2017)和火成演替群落分类(郭珂等,2019)进行了研究,然而,有关火干扰背景下喀纳斯泰加林林下草本层生物量和物种多样性方面的研究鲜有报道。喀纳斯泰加林火成演替一般要经历阔叶混交林、阔叶针叶混交林、针叶阔叶混交林和针叶混交林4个阶段(师瑞峰等,2009)。本研究以西伯利亚落叶松(Larix sibirica)火疤木的存在为研究前提,因此,调查中不涉及阔叶混交林。鉴于此,本文在确定火干扰背景下喀纳斯泰加林森林群落类型的基础上,探讨强烈度火干扰后(火后)40年不同森林群落草本层的碳汇功能以及草本层物种多样性的变化,为北方森林碳库的精准估算提供数据支撑,为森林可持续经营科学依据。
1 研究区概况与方法
1.1 研究区概况
喀纳斯国家自然保护区(86°54′-87°54′E,48°35′-49°11′N)位于新疆维吾尔自治区阿勒泰地区布尔津县境内。保护区地处欧亚大陆腹地,属温带高寒山区气候。年均气温-0.2 ℃,年均降水量1065 mm,年均蒸发量1097 mm,无霜期80-108 d。保护区内已知的维管束植物有83科298属798种。乔木优势树种为西伯利亚落叶松、西伯利亚云杉(Picea obovata)和西伯利亚红松(Pinus sibirica),伴生有西伯利亚冷杉(Abies sibirica)、疣枝桦(Betula pendula)和山杨(Populus tremula)等。灌木优势种有大叶绣线菊(Spiraea chamaedryfolia)、密刺蔷薇(Rosa spinosissima)、阿尔泰忍冬(Lonicera cearulea)和红果越桔(Vaccinium hirtum)等。草本优势种有黑穗苔草(Carex atrata)、白花砧草(Galium boreale)、白喉乌头(Aconitum leucostomum)和老芒麦(Elymus sibiricus)等。
1.2 研究方法
2016年和2017年6-8月,采用典型样方法在喀纳斯自然保护区未受过人为干扰的火烧迹地中设置382个火干扰样方。火干扰样方及其附近存在5株及5株以上的西伯利亚落叶松火疤木,且火疤的形成年份相同是设置样方的前提条件。样方大小为30 m×30 m,样方边界距离林缘至少50 m以上。同时,在每个样方的中心及4个角嵌套设置1 m×1 m草本层样方。
采用林木火疤年龄分析法确定样方火干扰的发生时间(徐化成等,1997),调查年份与火疤木成疤时间的差值称为火后时间。根据火疤木的外在属性因子(火疤高度、火疤宽度、火疤深度)确定林火烈度(fire severity)(刘翠玲,2009):强烈度(strong severity)、中烈度(moderate severity)、弱烈度(weak severity),根据林火烈度对调查样方进行分类。
记录样方的经纬度、海拔、坡度、坡向和坡位。采用每木调查法记录样方内大于1.3 m的所有树种的种类、数量、高度和胸高直径;样方内灌木的种类、数量、高度。目测并记录样方的灌木分种盖度。计算林冠层样方的郁闭度(stand coverage,Cove)、林分平均胸径(stand average diameter at breast height,DBH)和林分密度(stand density,SD)。记录草本层样方植物种名、数量、频度和草本层的盖度(herbaceous layer coverage,HC)和分种盖度。在草本层样方内用剪刀齐地面刈割并按莎草科、禾本科、豆科、杂草类4个功能群分类收集,同时收集小样方内的所有凋落物。将收集的草本植物和凋落物在105 ℃下杀青半小时后,置于85 ℃烘箱烘至恒重后称重,记录生物量(精度0.001 g)。同时,在每个样方沿坡体方向分别在上、中、下3个坡位各挖1个土壤剖面,记录土壤剖面特征,分别于0-10、10-25、25-70 cm土层用100 cm3的土壤环刀进行取样测定土壤容重(soil bulk density,BD),用木铲取3个剖面同一土层的混合土样1 kg,风干后去除杂物,过筛后进行测定分析。土壤性质测定如下:pH值——电位法(GB7859-87,pH);土壤有机质——重铬酸钾氧化-外加热法(GB7857-87,soil organic matter,SOM);全氮含量——半微量凯氏法(GB7173-87,total nitrogen,TN);全钾含量——火焰光度法(GB7854-87,total potassium,TK);全磷含量——钼提抗比色法(GB7852—87,total phosphorus,TP);土层温度(soil temperature,t)和土壤体积水分含量(soil water,WC)采用Hydra便携式土壤温/湿度测定仪(美国Eutech)在北京时间14:00左右测定。
1.3 数据分析
382个样方中共记录172种维管束植物,采用重要值作为群落分类依据(式1和式2):


式中,S为物种个数;相对优势度、相对高度和相对频度均为相对值;相对优势度为乔木的胸高断面的相对积值。
所有数据的统计分析与处理均在 Office 2007办公软件和SPSS 22.0程序中进行,采用单因素方差分析(One-way ANOVA)检验不同森林群落各变量的差异性;运用Originpro 2018作图。
已有研究将喀纳斯泰加林火成演替群落划分为6个群落类型:I. 西伯利亚云杉+疣枝桦-红果越桔-黑穗苔草+老芒麦群落;Ⅱ. 疣枝桦+西伯利亚云杉-红果越桔+大叶绣线菊-黑穗苔草群落;III. 西伯利亚落叶松+疣枝桦-红果越桔+密刺蔷薇-黑穗苔草+寄奴花群落;IV. 西伯利亚红松+西伯利亚落叶松-红果越桔+北极花(Linnaea borealis)-老芒麦+寄奴花(Cerastium pauciflorum)群落;V. 西伯利亚落叶松-红果越桔+阿尔泰忍冬-细叶野豌豆(Vicia tenuifolia)+老芒麦群落;VI. 西伯利亚落叶松+西伯利亚云杉-红果越桔-老芒麦+黑穗苔草群落(郭珂等,2019)。以此为依据根据植物重要值将382个样方划分到6个群落类型中,归属于火成演替的前期、中期和后期。为了分析火后不同森林群落草本层物种组成和地上生物量的变化,保证立地条件的一致性是前提条件。此次调查的样方中经历强烈度火干扰的样方数量最多。因此,以强烈度、火后时间40年、海拔1599-1704 m、坡度1-10 ℃、中坡、阴坡为条件选择样方,共筛选出 42个样方分析不同森林群落林下草本层物种丰富度和地上生物量变化。样方主要特征见表1。
对不同森林群落的林分特征、草本层盖度和土壤理化性质(3层的加权平均值)进行方差分析,结果表明,不同森林群落间仅郁闭度、林分密度、SOM和TN存在显著差异,其他因子间都不存在显著差异(表2)。因此,从这12个因子中选择郁闭度、林分密度、SOM和TN进行相关分析。
2 结果与分析
2.1 不同森林群落草本层丰富度和凋落物生物量的变化
6种森林群落的草本层物种丰富度存在显著差异(P<0.05),分别为:Ⅰ (26.0±1.6)、Ⅱ(25.1±1.4)、Ⅳ (22.2±2.2)、Ⅴ (21.6±1.5)、Ⅲ (21.2±1.3)、Ⅵ(19.3±2.5)(图1)。森林群落Ⅰ林下草本层丰富度最大,它处于火成演替的中期(针叶阔叶林)。强烈度火烧后,烧死大部分的乔木,形成面积较大的林窗/林隙,疣枝桦和五蕊柳(Salix pentandra)等先锋树种进入火烧迹地,喜阳的灌木和草本迅速定殖。随着演替的顺向进行,阔叶树种为西伯利亚云杉幼苗萌发创造了遮荫条件,形成针叶阔叶混交林,不耐荫的草本植物逐渐进入到林分中,草本层物种丰富度呈增大趋势。强烈度的火烧后40年,林分中的阳性和耐荫草本植物数量较多,因此其物种丰富度最大。Ⅵ和Ⅵ是以西伯利亚落叶松为优势种的针叶混交林,火烧残留了一部分大径级的西伯利亚落叶松,形成的林窗/林隙相对较小,进入的草本植物种类少。因此,草本层物种丰富度低。

表1 研究样方的主要特征Table 1 The main characteristics of study plots
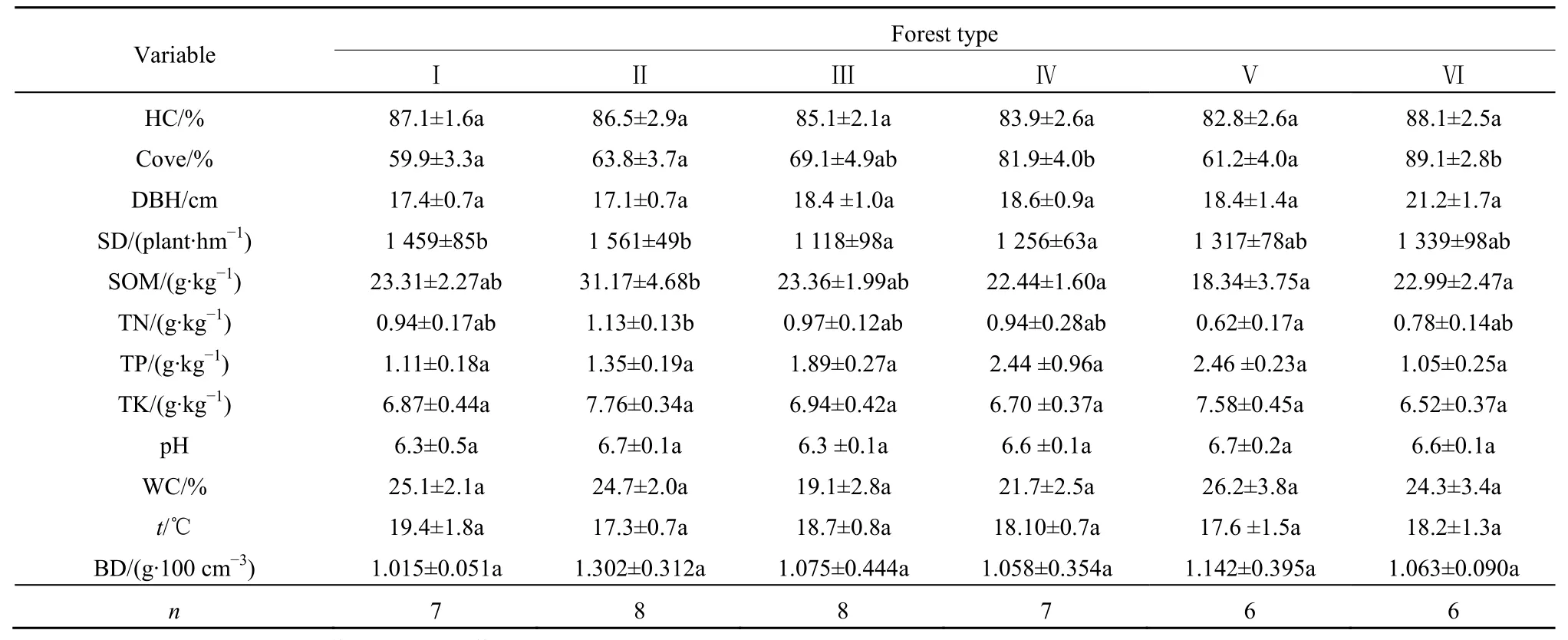
表2 不同森林群落林分特征、草本层盖度和土壤理化性质的变化Table 2 Changes of stand characteristics, HC and soil chemical and physical properties of different forest types

图1 不同森林类型草本层丰富度和凋落物生物量Fig. 1 Herbaceous layer species richness and litter biomass
对比 6种森林群落类型的凋落物生物量发现存在显著差异(P<0.05),排序为:Ⅱ>Ⅲ>Ⅰ>Ⅳ>Ⅴ>Ⅵ,与草本层物种丰富度变化并不相同,最大值(7.548±1.086) t·hm-2出现在群落Ⅱ中(图1)。Ⅱ和Ⅲ中疣枝桦占据一定优势地位,疣枝桦的生长速度快,新陈代谢也快,对凋落物的贡献大。Ⅵ的凋落物生物量最小,则是因为这些群落中阔叶林树种和灌木的种类和数量少,对凋落物的贡献相对小。
2.2 不同森林群落草本层各功能群和和总地上生物量的变化
草本层各功能群和草本层地上总生物量在6种森林群落类型间存在显著差异(P<0.05)(图2),排序为:I (0.853±0.015) t·hm-2>Ⅴ (0.719±0.033)t·hm-2>Ⅵ (0.639±0.021) t·hm-2>Ⅳ (0.596±0.023)t·hm-2>Ⅱ (0.568±0.023) t·hm-2>Ⅲ (0.478±0.017)t·hm-2。草本层地上总生物量的最大值出现在群落I中,与草本层物种丰富度的表现相同,但总变化趋势不同。各功能群地上生物量对草本层地上总生物量的贡献率因群落不同而异,豆科对草本层地上总生物量的贡献率为 2.21%-5.66%,禾本科为17.44%-24.95%,莎草科为 30.17%-57.37%,杂草为18.05%-48.48%。I、Ⅱ和V中莎草科的贡献率最大,Ⅲ中禾本科、莎草科和杂草的贡献率接近,草本层地上总生物量却最小,Ⅴ和Ⅵ中杂草的贡献率大于其他3个功能群。Ⅴ和Ⅵ为针叶混交林,林下生长有白喉乌头(Aconitum leucostomum)、西伯利亚耧斗菜(Aquilegia sibirica)、下延叶古当归(Archangelica decurrens)、阿尔泰藜芦(Veratrum lobelianum)、山地阿魏(Ferula akitschkensis)、直立老鹳草(Geranium rectum)等这一类个体较大的双子叶植物,因此,杂草的地上生物量较大。豆科在V中贡献最大(5.66%),这是因为林下有新疆山黧豆(Lathyrus gmelinii)和狭叶山黧豆(L.krylovii),它们的个体较其他豆科植物大得多。莎草科在所有群落中贡献率都大,表明黑穗苔草的适应性强,能在喀纳斯的森林群落中快速生长并占据优势地位。

图2 不同森林群落草本层各功能群和总地上生物量Fig. 2 Herbaceous layer functional groups and total aboveground biomass in different forest types
2.3 草本层地上生物量、物种丰富度、凋落物生物量、冠层结构、SOM和TN间的关系
采用相关分析解释6个森林群落中草本层地上生物量、物种丰富度、凋落物生物量、郁闭度、林分密度、SOM和TN之间的相关关系。森林群落类型Ⅰ中,林分密度与草本层物种丰富度呈显著负相关(r=-0.561,P=0.026);Ⅱ中草本层丰富度与郁闭度和林分密度呈显著负相关(r=-0.623,P=0.044;r=-0.579,P=0.032),SOM 与 TN呈显著正相关(r=0.865,P=0.001);Ⅲ中禾本科与豆科呈显著正相关(r=0.585,P=0.046),SOM与TN呈显著正相关(r=0.643,P=0.024);Ⅳ中杂草地上生物量与草本层丰富度呈显著正相关(r=0.544,P=0.039);TN与林分密度和 SOM 呈显著正相关(r=0.485,P=0.032;r=0.539,P=0.020);Ⅴ中杂草地上生物量与 SOM、TN和草本层丰富度呈显著正相关(r=0.874,P=0.023;r=0.866,P=0.026;r=0.891,P=0.017),与禾本科呈显著负相关(r=-0.893,P=0.027),草本层物种丰富度与郁闭度呈显著负相关(r=-0.883,P=0.020;SOM 与 TN呈显著正相关(r=0.830,P=0.041);Ⅵ中草本层物种丰富度与郁闭度呈显著负相关(r=-0.823,P=0.047),莎草科地上生物量与SOM呈显著正相关(r=0.962,P=0.009)。以上结果表明:喀纳斯泰加林强烈度火后 40年的林分郁闭度和林分密度对草本层物种丰富度的提高起抑制作用,草本层地上生物量受到土壤养分的限制,TN随着的SOM的增加而增加。草本层物种丰富度越高,杂草地上生物量越大。草本层地上生物量与凋落物关系不大,表明草本植物对凋落物的贡献小。
3 讨论
生态系统中生物多样性和生成力的关系一直是生态学研究最具争议的热点问题之一(Willig,2011),而生态系统的生产力水平是其功能的重要表现形式(Kokkori et al.,2012)。有控制试验表明,物种丰富度的提高会增加生物量和生产力(Cardinale et al.,2012)。与本研究结论不同,这可能是控制试验与野外调查的差异(高一飞等,2016);另外,本研究只分析了林下的草本层,而火成演替整个森林群落的物种多样性与森林生产力的关系还有待于验证。草本层物种丰富度的提高会增加杂草的地上生物量,与前人研究结论一致(Li et al.,2018),火后森林在恢复过程中,林下植物的相互作用对杂草类植物的生存起决定作用。环境条件不利的情况下物种丰富度和地上生物量呈正相关作用的可能性更大(Maestre et al.,2009),林分郁闭度和林分密度的差异,造成林下光环境的差异,林下杂草类植物间更多表现为互助关系(林敦梅等,2017),这种关系促进了杂草生物量的增加。本研究中,强烈度火后 40年的针叶阔叶林具有更高的草本层物种丰富度,有利于阳性草本植物和耐荫草本植物共同生长,当二者同时出现,物种多样性达到最大值,这在已有的研究中已得到证实(Loucks et al.,1970)。
凋落物生物量随不同森林群落的变化与物种丰富度的变化不同步,表明草本层对凋落物的贡献小。植物凋落物的积累减少群落物种的丰富度(Xiong et al.,1997),凋落物覆盖仅允许某些物种萌发且穿过凋落物层(Xiong et al.,2001)。本研究中,群落Ⅱ凋落物生物量最大,但物种丰富度最小,表明凋落物积累越多越不利于草本层物种多样性的提高。凋落物的碳储量远远大于草本层,凋落物在森林中的碳储量仅次于乔木层(雷蕾等,2011),它在森林碳汇功能中作用不可忽视。凋落物的现存量与凋落量呈正相关(黄宗胜等,2013),本研究中,阔叶针叶混交林的凋落物生物量为最大值,表明阔叶树占优势林分的凋落量大于针叶树占优势的林分。
草本层生物量是衡量草本层植物生长状况的重要指标,生物量越大表明其占有的资源越大,从长远角度考虑,其对群落的稳定与健康发展是有利的(王瑞华等,2014)。草本层地上总生物量随着演替阶段排序为:阔叶针叶林(0.568±0.014)t·hm-2<针叶阔叶林(0.633±0.020) t·hm-2<针叶混交林(0.651±0.023) t·hm-2,表明森林的生物量随着演替的进行呈增大趋势,因此,持续发挥着碳汇功能,与前人研究结论一致(Hudiburg et al.,2009)。喀纳斯泰加林火成演替群落中黑穗苔草为优势草本,但森林中竞争力强的物种生产力却未必总是贡献大(Szwagrzyk et al.,2009),而个体较大物种的数量强烈影响草本层地上总生物量。因此,黑穗苔草的地上生物量并不是在所有群落中都最大。禾本科在I和Ⅲ中的贡献率最大,表明老芒麦在针叶阔叶林中生长状况更好。草本层地上总生物量和物种丰富度的最大值都出现在针叶阔叶林中,表明针叶阔叶林更有利于维持草本植物生物多样性和生产力。
群落类型的不同,体现了生境的差异,其对林下植被的影响主要体现在林内光照、凋落物厚度和土壤性质等因素的变化(余敏等,2013)。本研究发现林分密度和郁闭度对林下草本层丰富度具有负效应,郁闭度越大越不利于草本层物种丰富度的提高。郁闭度越大的林分,进入到林下的光照越少,草本植物的生长会受到限制。当乔木层树冠盖度减少时,林下草本层的生物量增加(Shirima et al.,2015;Tessema et al.,2017)。但在本研究中,草本层地上总生物量与林分郁闭度和林分密度未表现出显著的相关性。可能是因为在强烈的火后 40年的6个森林群落中,莎草科和杂草对地上生物量的贡献率最大,但杂草中植物个体的大小决定其贡献率,而这些草本往往是中性植物,对冠层结构不是很敏感。
一些研究表明,草本植物生物量与土壤因子有显著的相关关系(Suchar et al.,2010)。本研究发现这6个森林群落的土壤理化性质中仅SOM和TN存在显著差异。因此,喀纳斯泰加林火成演替群落中草本层地上生物量与土壤养分因子表现出了一定的相关性,受SOM和TN的影响比较大,尤其是禾本科和莎草科的表现更明显,与前人研究结论一致(沙威等,2016)。
4 结论
随着喀纳斯泰加林强烈火干扰后 40年的森林群落的演替,草本层地上总生物量逐渐增加;草本层物种丰度未表现出明显的变化趋势,但二者的最大值都出现在针叶阔叶林中,表明针叶阔叶林更有利于维持草本层的物种多样性和生产力。凋落物的积累减少草本层的物种丰富度,因此清除凋落物可以提高草本层物种多样性。对喀纳斯林下草本层地上总生物量贡献最多的是莎草科和杂草,它们受林分郁闭度的影响较大。莎草科对草本层的物种丰富度贡献小,杂草对丰富度的贡献大。TN和SOM对杂草和莎草科的地上生物量有显著影响,表明火干扰对土壤中TN和SOM的影响较大,从而影响草本层的地上生物量。
综上所述,在喀纳斯泰加林强烈度火后40年的森林群落中对针叶阔叶林的枯枝落叶进行有效管理可以提高草本层生物多样性和生物量,有利于维持喀纳斯泰加林的物种多样性和提高其碳储量。本研究只分析了泰加林在强烈度火后40年不同森林群落的草本层变化,而森林植被层和土壤对不同林火烈度和火后时间的响应是今后研究的方向,可为火干扰群落的可持续经营提供更具体的科学指导。
猜你喜欢
小读者(2020年4期)2020-06-16
福建林业(2020年5期)2020-03-18
金桥(2019年12期)2019-08-13
作文周刊·小学四年级版(2019年8期)2019-04-23
热带林业(2019年4期)2019-03-05
西南林业大学学报(2018年5期)2018-10-23
国际木业(2018年3期)2018-09-10
农业环境科学学报(2017年11期)2017-12-07
中国三峡(2017年1期)2017-06-09
中华奇石(2015年10期)2015-07-09
