
增温对南亚热带混交林4个树种养分含量及化学计量的影响
2019-06-18 10:59李旭列志旸吴婷黄楚敏刘世忠刘菊秀傅松玲李义勇
生态环境学报 2019年5期
李旭 ,列志旸,吴婷,黄楚敏,刘世忠,刘菊秀,傅松玲,李义勇*
1. 安徽农业大学林学与园林学院,安徽 合肥 230036;2. 中国科学院华南植物园/退化生态系统植被恢复与管理重点实验室,广东 广州 510650
目前,大量监测与模型结果表明,全球气候变暖已成为毋庸置疑的事实。据气候模型预计,截至2050年,全球气温还将升高1-4 ℃(IPCC,2014),其中热带、亚热带地区增温幅度可能高于全球水平(Chen et al.,2015)。中国地表气温也发生了显著的变化,近百年来增温幅度和增温趋势皆高于全球平均水平(丁一汇等,2016)。温度的升高会改变植物体中生化过程的速率(Zhao et al.,2012),从而影响植物的养分吸收、代谢过程和生物量积累,其中植物的养分吸收对温度升高较为敏感(Dong et al.,2001;Xu et al.,2006)。因此,研究增温下的植物体内养分含量的改变对了解及预测植物在气候变暖下的响应和适应有重要意义。
碳(C)、氮(N)和磷(P)是生态化学计量学领域研究的主要对象,其中N和P是植物生长最重要的2个限制因子(Elser,2000)。植物的C/N和C/P可在一定程度上可反映单位养分供应量所能达到的生产力,N/P值反映了环境对植物生长的养分供应状况(Güsewell,2004;Tang et al.,2018)。气温上升影响植物的新陈代谢从而改变养分在植物器官中的含量(Gavito et al.,2005;Jónsdóttir et al.,2005)。Sardans et al.(2006,2008)的研究也发现,增温提高了欧石楠属灌木(Erica multiflora)叶的P含量。陈廷廷等(2018)研究增温对幼龄杉木(Cunninghamia lanceolata)细根养分含量的影响,发现增温使得杉木细根N含量升高,P含量降低,N/P升高,从而使杉木生长受P限制。前人的研究结果不一致与种间适应性差异有关。此外,由于树木的不同器官所行使的功能不同,同种树种的不同器官也具有不同的生态化学计量内稳性特征(张婷婷等,2018)。不同树种及各器官对增温的响应有所差异,植物的化学计量特征对增温的响应会对群落结构乃至生态系统功能变迁起到重要的推动作用(Sistla et al.,2012)。因此,阐明增温背景下植物养分含量及化学计量比的变化情况有助于理解植物在全球变暖背景下的响应和适应机制。江肖洁等(2014;2016)通过开顶箱增温法对牛皮杜鹃(Rhododendron aureum)、笃斯越桔(Vaccinium uliginosum)和东亚仙女木(Dryas octopetala var.asiatica)的研究发现,3种植物叶片的 C、N和 P化学计量学特性对增温的响应模式和尺度没有表现出一致性,K、Ca和Mg含量变化对增温的响应方式也不同,增温对3种植物的影响存在一定的种间差异性。由于这些营养元素都能限制植物的生长,南亚热带降水量大,对土壤盐基离子淋溶较严重,因此,除植物各器官中C、N和P化学计量学特性外,探清增温下K、Ca和Mg在各器官中的含量变化也极其重要。
温度是生态系统过程驱动的关键因素,以往有关增温实验的研究(Allison et al.,2010;Sardans et al.,2006;Sardans et al.,2008;Zhou et al.,2013),研究区域多局限于高纬度温带地区,有关热带及亚热带区域的研究相对较少(张莉等,2018;贝昭贤等,2018)。热带及亚热带森林虽然只占据地球陆地面积15%,但其植被碳库占陆地植物碳库2/3(Pan et al.,2013)。热带区域相对温带区域季节温差较小,有研究认为热带物种相对其他地区物种对气候变暖更加敏感(Wright et al.,2009;Krause et al.,2013)。相关研究之间虽存在一些相似结论,但均未总结出较统一的规律(Gavito et al.,2005;Jónsdóttir et al.,2005;Sardans et al.,2006;江肖洁等,2016)。因此,在热带亚热带地区开展增温实验,对准确预测未来全球变化背景下森林生态系统的响应有重要生态学意义。基于此,选择南亚热带针阔叶混交林4个关键树种为研究对象,研究了模拟增温对其叶、枝和根的主要养分元素含量及计量比的影响,旨在探清南亚热带针阔叶混交林不同树种的养分元素含量及其计量比对增温的响应情况。
1 材料与方法
1.1 研究区概况
研究地位于广东省肇庆市鼎湖山森林生态系统定位研究站,地理坐标为东经 112°30′39″-112°33′41″,北纬 23°09′21″-23°11′30″,属低山丘陵地貌,山体陡峭,构造线呈东北-西南走向,山峰海拔一般为450-600 m,最高峰(鸡笼山)海拔1000.3 m,平均坡度为30°-45°。鼎湖山自然保护区气候类型受东南季风控制,属亚热带季风型气候。年平均温度 21.4 ℃,最高月均温(7月)为28.0 ℃,最冷月均温(1月)为 12.6 ℃。年平均降雨量为1927 mm,主要集中在4-9月,约占全年降雨量的80%;年平均蒸发量1115 mm,年平均相对湿度80.3%。由于海拔上升所导致的小气候的改变,在不同的海拔梯度上形成了不同的林型,自上而下分别形成了山地常绿阔叶林、针阔叶混交林和季风常绿阔叶林。选择位于海拔300 m 处的针阔叶混交林为研究对象,其土壤类型为赤红壤,土层厚40-80 cm,土壤pH值为4.6,表土层有机质含量为4.3%(刘菊秀等,2013)。
1.2 试验设计
主要通过植物移植和土壤移位,在海拔300 m和30 m分别构建模拟针阔叶混交林,利用海拔梯度形成的温度差异研究其对森林生态系统的影响。在2012年1月,分别在海拔300 m和30 m,选择坡向、坡度一致的约30 m×30 m区域,去除区域内所有植物,形成相对独立的空旷地带。每个海拔分别建造3个样方(3 m长×3 m宽×0.8 m深),样方地下部分四周及底部铺上水泥和瓷砖,防止隔离样方内土壤与周围土壤的水分和养分交换。2012年4月,在位于海拔约300 m的混交林,按照不同层次(0-20、20-40、40-70 cm)分别收集土壤,分层混匀后填埋于所有样方内。土壤填埋后,移植该混交林中6个树种的约1年生树苗,每个树种选择6株树苗,按照相同株行距随机定植于样方内,每个样方内各树种种植位置相同。
本研究所选择的 4个树种分别是红枝蒲桃(Syzygium rehderianum)、红锥(Castanopsis hystrix)、马尾松(Pinus massoniana)和木荷(Schima superba)。其中,马尾松为针叶树种,其余均为阔叶树种。更多试验设计相关介绍详见文献(刘菊秀等,2013;Li et al.,2017;Fang et al.,2016)。
从2012年平台建立至2015年,通过对试验样地空气和土壤温度的测定,发现增温样地平均气温、月气温最小值以及月气温最大值分别比对照样地高约0.90、1.29、0.95 ℃,月均表层土壤(0-5 cm)温度约升高1.27 ℃(Li et al.,2017)。
1.3 植物养分含量的测定和计算
于2014年和2015年的12月采用全收割法对所有样方内的每个树种,随机选取1株长势良好的植株进行收割。收割后,按照叶、枝和根分别取样,然后带回实验室,用去离子水清洗干净,于 60 ℃烘箱内烘干至恒定质量,称质量并磨碎,取样测定全C、N、P、K、Ca和Mg含量。采用重铬酸钾外加热法进行全C含量测定,采用凯氏定氮法(Xu et al.,1993)进行全N含量测定,采用钼蓝比色法测定进行全P含量测定。全K、Ca和Mg含量使用等离子体发射光谱仪(ICP-AES;Optima-2000 DV,PerkinElmer,USA)测定。
1.4 数据处理
文中所有数据均运用Microsoft Excel 2010和SPSS 17.0统计软件进行分析,图表于 Sigmaplot 12.5及Microsoft Excel 2010中完成。采用多因素重复测量方差分析法(Multi-way Repeated Measures ANOVA)分析两个年份的不同处理(增温与对照)、不同树种(红锥、红枝蒲桃、马尾松和木荷)和不同器官(叶、枝和根)C、N、P、C/N、C/P、N/P、K、Ca和Mg含量及其比值的差异;采用单因素方差分析法(One-way ANOVA)分析每个年份增温与对照间各指标的差异。显著性水平设定为α=0.05。
2 结果与分析
2.1 增温对南亚热带针阔叶混交林中不同树种各器官C、N和P含量的影响
结果表明,增温降低了各树种的C和N含量,升高了P含量(P<0.05)(表1)。各树种C含量(425.05-514.45 mg·g-1)的变化幅度较小,叶的N和P含量大于枝和根(图1)。不同树种对增温的响应有索差异,红锥和木荷的 C、N、P含量对增温响应大于红枝蒲桃。具体而言,增温显著降低了红枝蒲桃2015年枝的N含量,升高了根的P含量(P<0.05)。增温显著降低了红锥2014年根和2015年叶的C含量,2014年枝、根和2015年根的N含量(P<0.05);升高了2014年叶、枝和2015年枝的P含量(P<0.05)。增温显著降低了马尾松2014年枝和2015年叶的C含量(P<0.05);升高了2015年叶、枝的P含量(P<0.05)。增温显著降低了木荷2014年根和2015年叶的C含量,2014年叶的N和P含量(P<0.05),升高了2015年根的N含量,叶的P含量(P<0.05)。
2.2 增温对南亚热带针阔叶混交林中不同树种各器官化学计量比的影响
增温显著降低了各树种各器官的 C/P和 N/P(P<0.05)(表1)。各树种叶的C/N低于枝和根(图2)。具体而言,增温显著降低了红枝蒲桃2015年根的C/P(P<0.05)。增温显著升高了红锥2014年和2015年根的C/N(P<0.05);降低了2014年叶、枝和2015年枝的C/P,2014年叶、枝、根和2015年根的N/P(P<0.05)。增温显著降低了马尾松2015年叶的 C/P,叶和枝的 N/P(P<0.05)。增温显著升高了木荷2014年叶的C/N、C/P(P<0.05);降低了2015年根的C/N,2014年和2015年根的C/P(P<0.05)。各树种在化学计量比上对增温的响应表现为红锥>木荷>马尾松>红枝蒲桃。
2.3 增温对南亚热带针阔叶混交林中不同树种各器官K、Ca和Mg含量的影响
增温升高了各树种叶的 Ca含量,其他养分在不同器官没有表现出一致的规律(图 3)。整体而言,不同树种各器官间,叶的K含量大于枝和根。具体而言,增温显著升高了红锥2015年枝的K含量(P<0.01)。增温显著升高了马尾松2014年叶和根的K,2014年和2015年叶的Ca含量(P<0.05);降低了2015年根的Mg含量(P<0.05)。增温显著升高了木荷2015年根的K含量,2014年叶和2015年根的 Mg含量(P<0.05);降低了 2014年枝的Ca含量(P<0.05)。
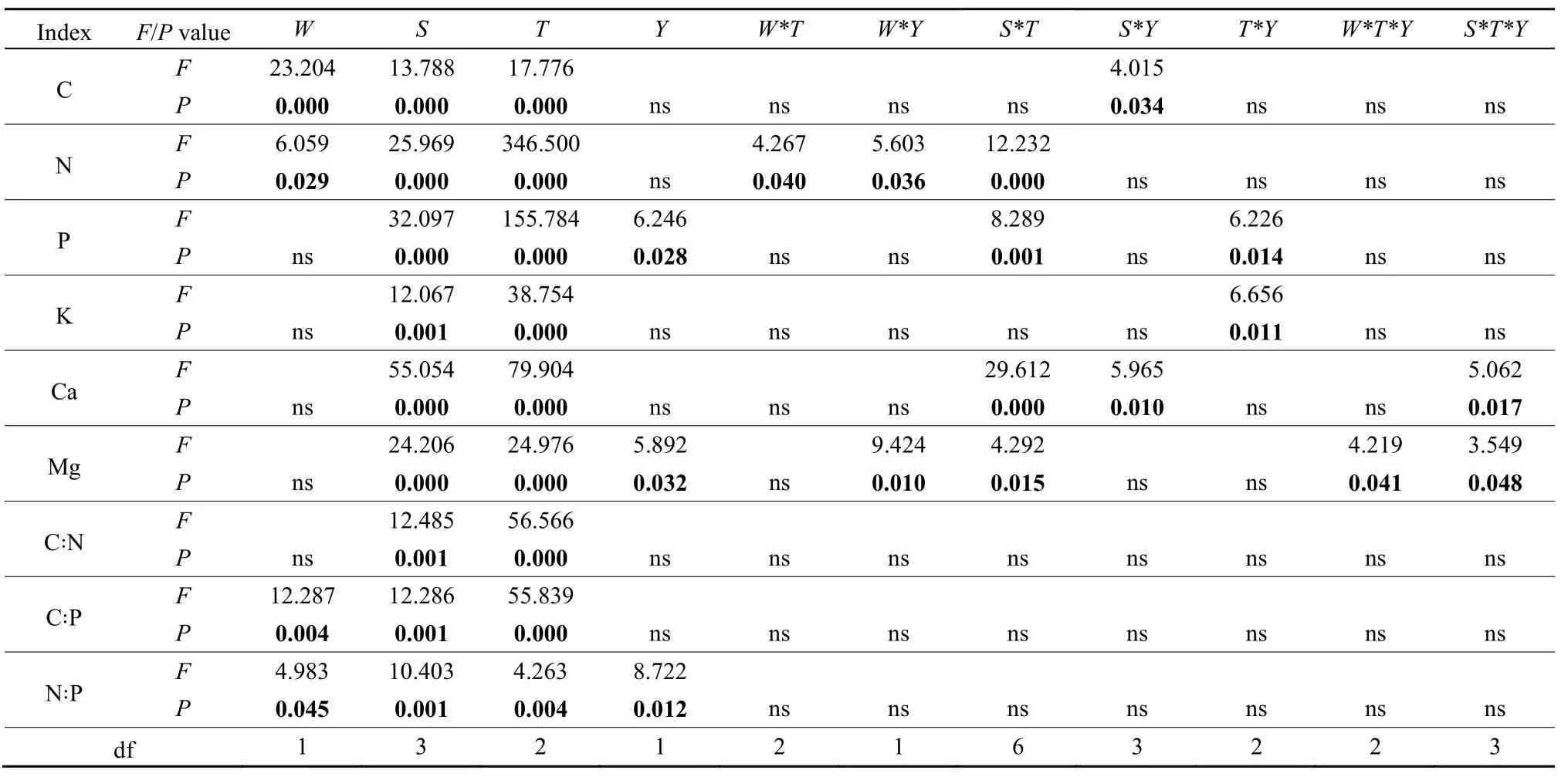
表1 增温对不同树种养分含量及化学计量比的影响(多因素方差分析)Table 1 Effect of warming on nutrient concentrations and stoichiometric of different tree species (Multi-way ANOVA)
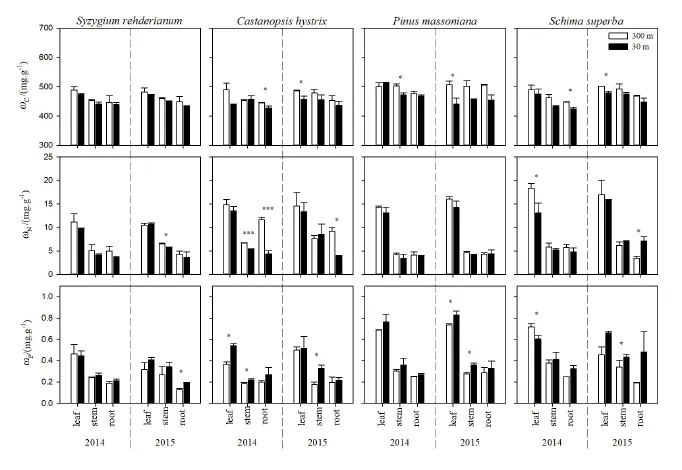
图1 增温对不同树种的C、N、P含量的影响Fig. 1 Effect of warming on carbon (C), nitrogen (N) and phosphorus (P) concentrations of different tree species
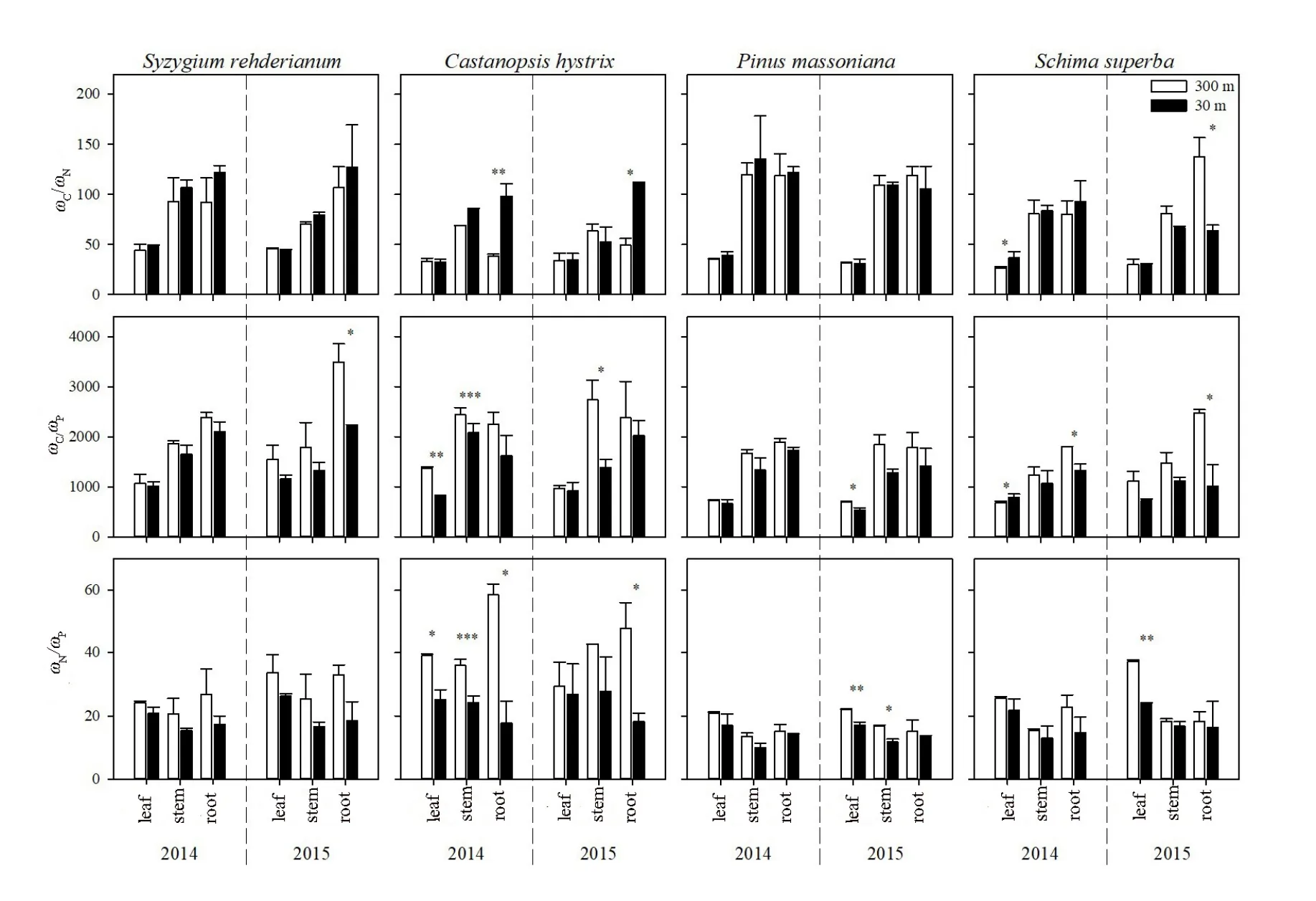
图2 增温对不同树种的CNP化学计量比的影响Fig. 2 Effect of warming on C/N, C/P and N/P of different tree species
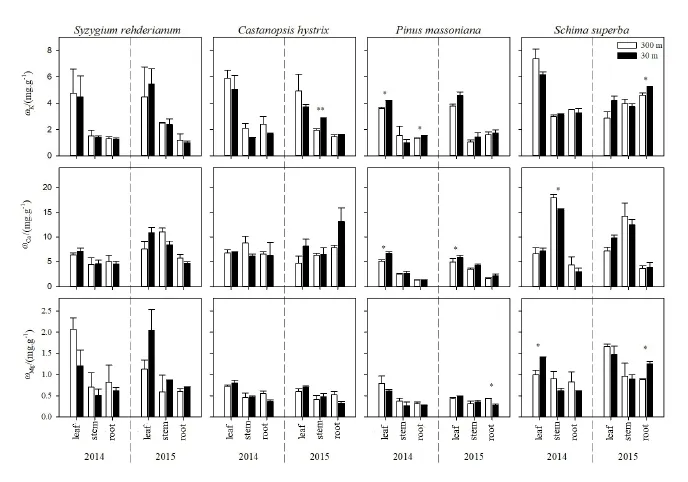
图3 增温对不同树种的K、Ca、Mg含量的影响Fig. 3 Effect of warming on K, Ca and Mg concentrations of different tree species
3 讨论
3.1 增温对南亚热带针阔叶混交林不同树种各器官C、N和P含量及化学计量比的影响
叶片养分含量的动态变化与树木生长状态息息相关。本研究发现增温降低了不同树种各器官C和N含量,这与增温引起生长速率加快导致的不同树种各器官C和N含量被“稀释”有关,与他人研究结果类似(Han et al.,2005;Poorter et al.,2006;Li et al.,2017)。另外,植物N含量降低还可能是由于增温使土壤N含量显著降低(Liu et al.,2017),降低了植物对N的吸收。此外,增温升高了不同树种各器官的P含量,说明增温促进了植物对P的吸收。贝昭贤等(2018)在亚热带进行的模拟增温试验结果表明,增温促进了有机物的分解,有效P含量显著增加。植物体内的P主要是根系从土壤中吸收而来,南亚热带地区是典型的缺P土壤生境(Liu et al.,2013),增温可能引起土壤有效P的增加(Jiang et al.,2016; Liu et al.,2017b),有利于植物吸收 P。
C、N和P化学计量比是生态系统过程及其功能的重要特征,它体现了C的积累动态及N、P养分限制格局,同时也揭示了生长速率与养分分配的关系(张慧等,2016)。通常情况下,植物叶C/N和C/P能反映植物N和P的养分利用效率,反映了植物在吸收营养过程中对碳的同化能力和养分利用效率(Wang et al.,2014)。研究发现增温升高了各树种叶的C/N且均高于全球平均水平(22.5)(Elser et al.,2000)。说明增温使4个树种N的利用效率升高。这可能是由于土壤有效态N含量降低(Liu et al.,2017),从而降低植物对N的吸收,导致植物叶C/N升高。Liu et al.(2017)研究证明了增温会促进微生物对凋落物的分解,本研究发现的C/N升高可能缓解了这种效应。此外,增温降低了各树种的C/P,这可能是增温使土壤有效P含量增加,促进了植物对P的吸收。由于植物体内C含量减少和P含量增加,使得各树种器官C/P降低,从而有利于植物生长以获取更多的养分和水分资源。研究还发现,增温还降低了植物的N/P,这是由于植物 N含量的降低和植物 P含量的升高共同导致的,与多数研究者的结论相同(Elser,2000;Han et al.,2005;田地等,2018)。
增温对不同元素的吸收及运输影响程度不同,可能改变元素在器官间的分配以及不同元素间的关系。叶片通过光合作用固定CO2合成糖类,通过韧皮部向下运输,供给到其他器官用于呼吸代谢及生长利用;N和P主要通过根系吸收,随着水分通过木质部向上运输,用于光合作用、蛋白质合成及提供能量物质等(王凯等,2018)。叶片、细根是植物代谢十分活跃的器官,而枝和主根系是基本的结构支撑和养分储存器官。这些器官间N和P的计量关系紧密(Craine et al.,2005;Kerkhoff et al.,2006),这种计量关系取决于各器官的生长特性和功能属性。其中,水分对养分的稀释作用,导致茎干和根的N和P含量低于叶(Kerkhoff et al.,2006)。这是由于植物在不同器官间分配养分时,通常会优先传输大量的营养元素到叶片中,以维持植物生产功能的稳定(Schreeg et al.,2014;Mo et al.,2015)。本研究中,不同树种叶的C、N和P含量均大于枝和根,与前人研究一致(田地等,2018)。叶的N和P含量大于枝和根,叶的C/N低于枝和根,不同树种各器官间的C/N以根最高,叶最低。养分含量在各器官的分布规律证实了不同器官具有不同的生理机能与养分吸收和蓄积能力,使得养分在同一树种不同器官中的分布特征存在差异(郑跃芳等,2017)。增温显著降低了各树种各器官的C/P和N/P,说明这4个树种应对增温的策略可能是更多的营养分配到叶,以提高植物生长速率,从而在群落中形成优势。4个树种在C、N和P化学计量比对增温的响应上表现为红锥>木荷>马尾松>红枝蒲桃,说明增温对南亚热带4个主要树种的影响存在一定的差异性。
3.2 增温对南亚热带针阔叶混交林不同树种各器官K、Ca和Mg含量的影响
研究发现不同树种各器官养分含量总体上表现为 Ca>K>Mg>P,与罗绪强等(2018)的研究结果一致。这可能是由于植物对不同养分的需求量不同,K、Ca和Mg是植物体内必需的大量矿质营养元素,也是限制全球植物生产的主要营养物质(Vitousek et al.,1991),而南亚热带植物具有P供应不足的生境特征(Liu et al.,2013)。增温对各树种器官K含量的影响不同,叶的K含量大于枝和根。这可能是由于增温加快了微生物对凋落物的分解速率(Liu et al.,2017),加快了养分元素的循环,另一方面,增温样地植物生长加快(Li et al.,2017),植物体内的K主要来源于根系从土壤中的吸收。而K流动性较大,由于生长速率及生物量分配的不同,可能导致K流向叶和枝(Maathuis,2009;Karley et al.,2009)。增温升高了各树种叶的 Ca含量,这可能是由于增温加快了植物生长速率,叶片生物量增加迅速,且增温加快了微生物对凋落物的分解速率(Liu et al.,2017),加快了养分元素的循环。Ca作为细胞壁的主要成分,还参与气孔调节(Maathuis,2009),这使得Ca可能会流向叶片。不同树种对增温的响应有差异,马尾松和木荷的K、Ca和Mg含量对增温的响应大于红枝蒲桃和红锥。
4 结论
本研究结果揭示了南亚热带针阔叶混交林主要树种养分含量及化学计量学特征对增温的响应特征,发现不同树种、不同器官中的养分元素及化学计量学特征对增温的响应并不完全相同,说明增温对南亚热带主要乡土树种的影响存在一定的种间差异性,这种差异可能会进一步改变森林群落的结构和功能。总体而言,本研究说明增温促进了南亚热带混交林4个树种的生长,然而本次研究仅跟踪了2年,为了进一步了解全球变化背景下,植物养分和化学计量关系对生态系统功能的影响,需要进行长期的研究。
猜你喜欢
流行色(2021年8期)2021-11-09
今日中国·西班牙文版(2021年4期)2021-04-09
汉语世界(The World of Chinese)(2019年5期)2019-11-11
亚热带农业研究(2019年2期)2019-09-05
水土保持研究(2019年3期)2019-05-22
现代园艺(2018年2期)2018-03-15
少儿科学周刊·少年版(2018年12期)2018-01-26
星星·诗歌原创(2017年5期)2017-08-13
现代园艺(2017年11期)2017-06-28
科学中国人(2017年36期)2017-06-09
