
三江源农牧交错区一个种植周期的垂穗披碱草人工草地CO2通量变化特征
2019-06-18 10:59贺福全李奇陈懂懂赵新全罗彩云许茜陈昕赵亮丁生祥赵金忠
生态环境学报 2019年5期
贺福全 ,李奇,陈懂懂,赵新全,罗彩云,许茜 ,陈昕 ,赵亮*,丁生祥,赵金忠
1. 中国科学院西北高原生物研究所,青海 西宁 810008;2. 中国科学院大学,北京 100049;3. 青海省同德气象局,青海 同德 813299
人类对土地的有效利用和管理有助于生物圈与大气之间CO2交换的季节性变化(Zeng et al.,2014),草地管理与 CO2吸收和排放密切相关(Tubiello et al.,2015;Wollenberg et al.,2016)。草地有效管理可以提高碳汇能力(尽管年增量会随着时间的推移而减少)(Hortnagl et al.,2018;Smith,2014),施肥可使土壤碳含量增加 5%,封育增加6.3%,耕地转化为人工草地增加6.4%,退化草地恢复治理增加42.8%(Wang et al.,2011)。近年来,为了恢复三江源区退化草地,三江源区的人工草地面积在不断扩大,目前约为67000 hm2(Zhao et al.,2018)。Li et al.(2005)报道人工草地建植20年后土壤碳储量也有所下降,也有少量文献对三江源区的人工草地 CO2通量研究进行了报道(赵亮等,2008;吴力博,2010),认为人工草地是一个弱的碳汇。这些已发表的文献报道均是在较短的时间序列上开展的研究。
在过去的 50年中,由于气候突变和人为活动增加,青藏高原超过50%草地退化或沙化(Harris,2010;秦大河等,2014),这不仅会严重影响草地生态系统生产力和服务,还危及到畜牧生产,地方经济和区域可持续性发展(Zhao et al.,2018)。在极度退化的“黑土型”退化草地上建植人工草地是可行的,也是快速恢复其植被的主要途径(马玉寿等,2002,王启基等,2000)。另外,中国实施的“退耕还林(草)”工程,需求大量的适宜草种。为了保证工程的实施,在三江源东部农牧交错区发展草种繁育产业。通过各种农艺措施,种植多年生禾本科草种,为了保证草种的产量,以5年为一个种植周期。Luyssaert et al.(2007)表明净生态系统CO2交换(Net ecosystem CO2exchange,NEE)在全球尺度上是由气候因素决定的,而在站点尺度上由站点性质和管理措施决定的,因此关于三江源东部农牧交错区多年生人工草地单个站点管理措施与碳收支年际变异关系的研究是很有必要的。
目前,对青藏高原不同天然植被类型的净生态系统CO2交换进行了长期的研究(Kato et al.,2004,2006;Zhao et al.,2005a,2005b,2006,2010;Li et al.,2016;张法伟等,2008;柴曦等,2018),认为生态系统的源汇功能很大程度受生长季的降水量、强度和分配的影响(Kato et al.,2006;Zhao et al.,2006)。赵亮等(2007)通过比较青藏高原不同天然植被类型的净生态系统CO2交换,认为净生态系统CO2交换由自下而上的气候因子(温度、降水)和自上而下的生物因子(群落结构、叶面积和放牧)两条途径控制。然而,这些研究都集中一个比较稳定的群落结构,但对于一个不稳定且有人类干扰的植被群落,净生态系统CO2交换的变化特征受到的关注尚不足,Xu et al.(2017)也表明,建植 3-5年的人工草地群落结构单一,且不稳定。Zhao et al.(2015)通过碳同位素标记法证明物种的生活史及物种丰富度影响生态系统的碳转移(Xu et al.,2017)。由此看来,变化的群落结构对生态系统碳收支存在着一定影响。本研究采用涡度相关法技术定量分析三江源东部农牧交错区多年生人工草地5年CO2通量及其环境要素变化,分析了一个完整的种植周期碳收支状况,旨在探究人工草地碳收支的日、季节和年际变化特征和规律,分析环境因子和人工草地管理措施对碳收支的影响。
1 材料和方法
1.1 研究区概况

图1 观测样地Fig. 1 Observation site
试验样地位于三江源东部青海省海南州同德县同德牧场(35°15′11″N,100°41′57″E,海拔为 3305 m),该区域属典型高原大陆性气候,辐射强烈,无绝对无霜期,年均温为3.7 ℃,年降水量为427.2 mm,土壤类型为暗棕壤。人工草地于2008年建成,由翻耕草带和未翻耕草带组成,翻耕草带和未翻耕草带宽均 4 m,相间分布(图 1)。翻耕草带每 5年重新翻耕(5年为一个种植周期),草带为单播的垂穗披碱草(Elymus nutans)。在6月初种植,种植当年不收割牧草,其他年份8月底收割牧草。未翻耕草带主要物种为兰石草(Lancea tibetica)、紫花苜蓿(Medicago ruthenica)、冷蒿(Artemisia frigida)和垂穗披碱草。本研究利用2012-2016年完整的观测数据进行分析。2011年原本是种植第一年,由于数据缺测,而 2016年重新翻耕种植,故以 2016年代替 2011年作为参考年(1yr);2012年为种植第2年(2ed yr),2013年为种植第3年(3rd yr),2014年为种植第4年(4th yr),2015年为种植第5年(5th yr)。
1.2 研究方法
1.2.1 通量观测
涡度相关和常规气象观测系统置于地势平坦、视野开阔并具有足够大“风浪区”的人工草地中心。采用三维超声风速仪(CSAT3,CSI,USA)和开路红外气体分析仪(LI-7500,LI-COR,USA)监测生态系统净CO2交换的变化,采样频率为10 Hz,每30分钟输出1次平均值,数据储存于数据采集仪(CR3000,Campbell Scientific,Logan,UT,USA)中。同时,其他环境数据利用微气象法,存储于数据采集仪(CR1000,Campbell Scientific Inc)中,观测各要素及其使用仪器和观测高度(表 1)。每年7月底将分别用高纯度的N2(99.999%)、标准CO2气体来标定CO2分析仪,用露点仪(LI-610,LI-Cor,USA)标定H2O的量程。
1.2.2 数据处理和插补
为保证数据的可靠性,所有微气象数据和通量数据均进行数据质量控制。数据预处理过程中去除夜间(PPFD<10 µmol·m-2·s-1)的数据,并舍弃摩擦风速u*<0.1 m·s-1时的数据。为了确保可靠地处理通量数据,采用了 ChinaFLUX推荐的通量数据处理的标准方法(Yu et al.,2006)。其中包括三维坐标旋转(Zhu et al.,2005)、WPL 校正(Webb et al.,1980)、存储项计算、异常值剔除、剔除夜间CO2通量负值(生态系统碳吸收)数据(Reichstein et al.,2005)、缺失数据插补(Falge et al.,2001)和NEE通量拆分(Reichstein et al.,2005)。利用环境变量和有效通量数据,运用非线性函数估算缺失的CO2通量数据。使用公式(1)插补夜间缺失的通量数据(Yu et al.,2008)。公式(1)将利用白天有效数据和光合有效辐射(PAR)拟合得到的公式(2)插补白天缺失的通量数据。
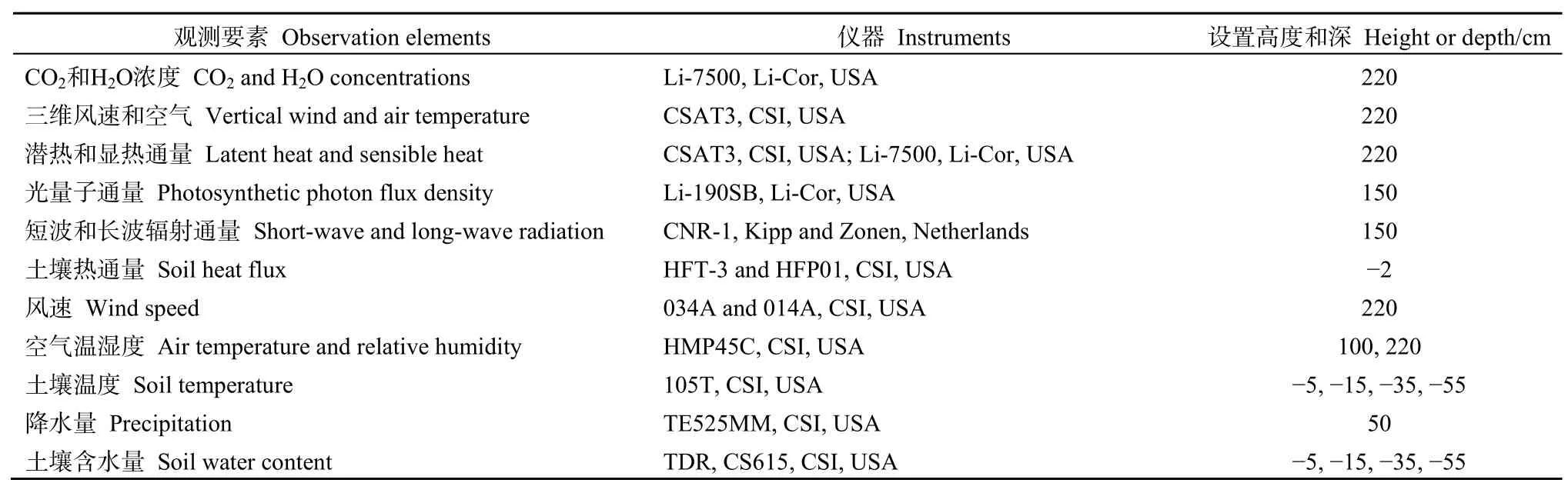
表1 观测要素所使用的仪器及其安装高度Table 1 Measurements of meteorological parameters, instruments and location
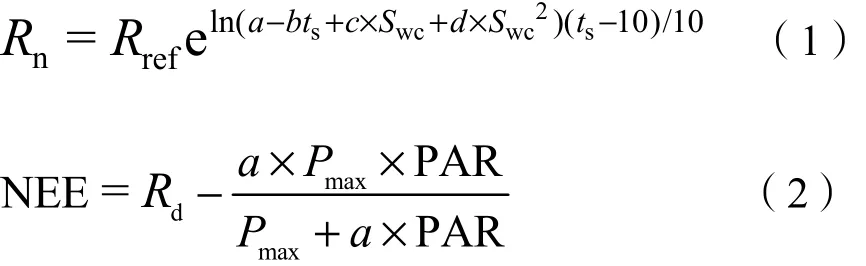
式中,Rn为夜间缺失CO2通量及白天R通量;Rref为土壤温度10 ℃时的参考生态系统呼吸速率;Rd是生态系统暗呼吸率;a和Pmax是表观量子产率和饱和光合成速率。ts为5 cm深处的土壤温度,Swc为5 cm深处的土壤含水量(m3·m-3),a、b、c和d是适合全年数据的特定地点参数,其中b>0,d≤0。
涡度相关技术测得的 CO2通量是净生态系统CO2交换量(NEE),即生态系统光合作用所固定的CO2(GPP)与生态系统呼吸排放的 CO2(R)间的收支差额。利用夜间的有效观测数据和公式(1)可获得 R与土壤温度(ts)和土壤含水量(Swc)之间的关系函数,再利用此函数估算生长季白天的生态系统呼吸。生态系统日总呼吸量是生态系统昼夜呼吸量的总和。用日间的NEE加上日间的R,则可得到生态系统的光合生力,即生态系统的光合固碳量:

能量闭合程度分析是检验通量数据质量的有效手段。能量平衡方程:

式中,Hs为显热通量;LE为潜热通量;Rn为净辐射;G为土壤热通量,土壤热通量是两块热通量板平均值,冠层热储量忽略不计。能量闭合方程斜率分别为 0.76(2012)、0.76(2013)、0.73(2014)、0.72(2015)、0.89(2016)。能量闭合斜率在可接受的范围之内(0.7-0.9)(Aubinet et al.,2000;Wilson et al.,2002)。闭合斜率偏小,可能是由于忽略了冠层热储量和仪器系统误差造成的。2012、2013、2014、2016年有效数据在 75%以上,2015年有效数据为70%。
根据多年观测和文献总结(Zhang et al.,218),该人工草地生长季为4月中旬至9月初,返青期为4月20日-6月30日,生长季旺期为7月1日-8月31日,枯黄期为9月1日-10月15日,非生长季节为1月1日-4月19日和10月16日-12月31日。生长季长度根据播种和收割的时间,以及总初级生产力(Gross primary productivity,GPP),将GPP>1 g·m-2(以C计)定义为生长季,生长季天数即生长季长度。
1.2.3 生物量及群落结构测定
在每年8月中旬进行地上生物量和植被群落的测定,先进行群落调查,采用样方法,随机选择 4个50 cm×50 cm的样方,调查物种组成、群落总盖度、各物种高度和盖度,用于Simpson指数的计算。物种盖度调查采用目测法,样方随机设置,每个样方每个物种随机选取5株测量高度。待群落调查完,进行地上生物量的测定,用剪刀齐地面剪下装入自封袋中,带回实验室于 65 ℃恒温烘箱内烘干至恒重后称重。多样性计算公式如下:


式中,Pi表示某个草地类型中第i个物种的相对重要值;Ni表示该草地类型中的第i个物种的重要值;N表示该草地类型中所有物种重要值之和。
1.2.4 统计分析
采用线性回归、多元逐步回归、单因素方差分析以及简易的结构方程模型进行统计分析。线性回归和多元逐步回归在SPSS 15.0(SPSS,Chicago,USA)中进行。应用结构方程模型采用R软件“sem”评价生长季长度(Growing season length,GSL)对NEE年际变异的直接和间接影响(丁金枝,2017)。
2 结果
2.1 非生物因子与生物因子特征

图2 一个种植周期年均温(MAT,℃,a)、年降水量(MAP,mm,a)、地上生物量(AGB,g·m-2,b)、辛普森多样性指数(Simpson index,c)和生长季长度(GSL,days,d)变化特征(除了种植第二年)Fig. 2 Statistical characteristics of mean annual temperature (MAT, ℃,a), mean annual precipitation (MAP, mm, a), green aboveground biomass(AGB, g/m2, b), plant community characteristics (Simpson’s index, c), and growing season length (GSL, days, d) over a plant cycle and the age of planting except for the second planting year
图2a-图2d显示了年均温(MAT,a)、年降水量(MAP,a)、地上生物量(AGB,b)、Simpson多样性指数(SP,c)和生长季长度(GSL,d)随种植年限的变化状况。由图可知,2ed yr环境因子MAT低于其他年份(<10%平均温度),1yr、2ed yr的 MAP高于其他年份(>10%平均降水量)。生物因子 AGB在一个种植周期内存在明显差异,4th yrAGB [(658.00±114.76) g11-2]显著高于其他年份,3rd yr AGB [(322.83±55.28) g·m-2]和 5th yr[(530.22±156.56) g·m-2]之间差异不显著,而显著高于 1yr [(44.49±21.17) g·m-2](图 2c),4 年平均 AGB为[(388.88± 267.94) g·m-2]。3rd yr Simpson 指数为最高(0.73±0.09),显著高于 1yr (0.58±0.15)、5th yr(0.61±0.11)和 4th yr (0.56±0.09),而其他年份之间差异不显著(图2d),平均SP为(0.62±0.08)。生态系统的稳定性与群落生物量的变异性相反,稳定的群落一般都具有高多样性(Bai et al.,2004),人工草地在5年内,AGB波动较大且SP偏低,群落处于不稳定状态。Xu et al.(2017)研究表明,建植3-5年的人工草地群落结构变得单一且不稳定。图2d显示,一个种植周期内GSL随着种植年限的增加而增加。
由图3可知,空气温度ta(Air temperature,ta,图3a)、5 cm土壤温度5 cm-ts(soil temperature,ts,图3b)和光量子密度通量PPFD(photosynthetic photon flux density,PPFD,图3e)均呈单峰曲线,变化不明显。同时,可以看出降水量P(图3c)常年分配不均匀,P的峰值出现时间基本与ta和ts的峰值一致,而土壤含水量Swc(图3c)的变化趋势与P相同。饱和水汽压差VPD(图3d)的变化趋势没有明显规律,生长季高于非生长季。进一步分析发现,降水量主要集中在5-9月,占全年的87%,P与ta的峰值均出现在8月前后,雨热同期(图3c)。
2.2 CO2通量变化特征
2.2.1 NEE日变化
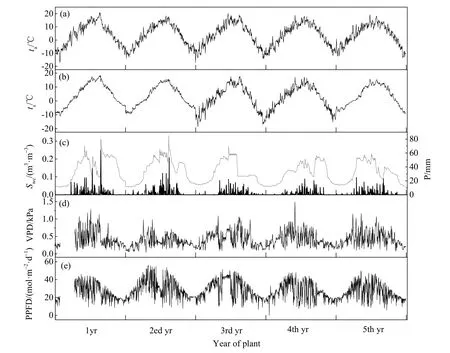
图3 主要环境因子空气温度(ta,℃,a),5 cm土壤温度(ts,℃,b),5 cm土壤含水量(Swc,m3·m-3,c),降水量(P,mm,c)饱和水汽压(VPD,kpa,d)和光量子通量密度(PPFD,mol·m-2·d-1,e)的季节变化Fig. 3 Air temperature (ta, ℃, a), average 5 cm soil temperature (ts, ℃, b), soil water content at a depth of 5 cm (Swc, m3·m-3, c), and precipitation (P, mm, c),vapor pressure deficit (VPD, kPa, e) and photosynthetic photon flux density (PPFD, mol·m-2·d-1, d) over a production cycle and the age of planting
图4 所示为不同季节不同种植年份NEE的日变化。在植物生长阶段,日变化规律明显,表现为白天吸收,夜间排放,上午8:00左右NEE由正值(释放)转换为负值(吸收),中午前后(10:30-14:00)吸收量达到最大,随后吸收量开始减少,大约在19:00NEE由负值转换为正值,正负转换时间月际变化明显(图4a-图4c)。在植物非生长阶段,NEE的日变化不明显,变化幅度很小,表现为向大气中释放CO2(图4d)。在返青期,NEE日变化幅度差异较大(图4a)。平均碳吸收时间为(9.3±1.64)h,5th yr吸收峰值为最大(-0.24 mg·m-2·s-1),出现在 11:00。吸收峰值最小的为 1yr(-0.07 mg·m-2·s-1),出现在 10:30,5th yr的吸收峰值是 1yr的3.4倍。除2ed yr外,5th yr的吸收峰值显着高于其他年份,而2ed yr的吸收峰值显著高于1 yr(图4a)。在生长季旺期,不同种植年份之间NEE日变化差异明显(图4b)。平均碳吸收时间为(9.5±1.12)h,2ed yr吸收峰值为最大(-0.73 mg·m-2·s-1),出现在 12:00,而 3rd yr的为最小(-0.47 mg·m-2·s-1),出现在12:00,2ed yr的吸收峰值是3rd yr的1.6倍。2ed yr吸收峰值明显高于其他年份,而4、5th yr吸收峰值显著高于1、3rd yr(图4b)。枯黄期NEE的日变化在不同年份之间差异较大(图 4c),平均碳吸收时间为(8.6±1.14) h,1yr吸收峰值为最高(-0.33 mg·m-2·s-1),出现在 13:00 。5th yr吸收峰值为最低(-0.13 mg·m-2·s-1),出现在 11:30,1yr 吸收峰值是5th yr的2.6倍。除4th yr外,1yr的吸收峰值显著高于其他年份(图 4c)。在非生长季节,不同种植年份之间 NEE日变化非常弱,几乎没有碳吸收,表现为向大气中释放CO2(图4d)。
2.2.2 NEE的季节变化
由图5可知,人工草地生态系统生长季,GPP、R和NEE峰值出现时间不同。GPP在植物生长阶段呈先增加后减小的单峰曲线,在3月底4月初开始增加,3rd yr GPP 在 6 月达到最大值(208.4 g·m-2),高于其他年份,2、5th yr GPP在7月达到峰值,最大值分别为 174.94、169.56 g·m-2,而 1、4th yr GPP在8月才到达最大值(图5a),最大值分别为130.13、146.48 g·m-2。R在1-2月和11-12月基本保持恒定,3月开始随着温度的回升而增加,1、2、3rd yr R 在 8 月达到最大值(79.17、111.16、156.52 g·m-2),4、5th yr R 在 7 月达到最大值(89.87、88.98 g·m-2),3rd yr R最大值为156.5 g·m-2,高于其他年份(图5b)。NEE在1-4月中旬和9月上旬至12月均为正值(碳源),3月开始增加,到4月底5月初达到最大值,随后减小转为负值(碳汇)。就逐日变化而言,不同年份从碳源转为碳汇再转为碳源(由正→负→正)的时间不同,1yr为7月初(7月1日),2ed yr为5月底(5月31日),3rd yr为5月初(大约为5月5日),4、5th yr都在5月下旬(大约为5月25日和5月21日)转为碳汇(正→负)。由于收割,2-4th yr在9月初(约9月4日前后)均转为碳源(负→正),而1yr在9月底(约9月29日)才转为碳排放。NEE到达吸收峰值的时间和 GPP相同,3rd yr在6月到达最大碳吸收,为-88.1 g·m-2,而2、5th yr,在7月达到吸收峰值(-67.63、-80.58 g·m-2),1、4th yr在 8月达到吸收峰值,分别为(-50.97、-60.22 g·m-2)(图 5c)。
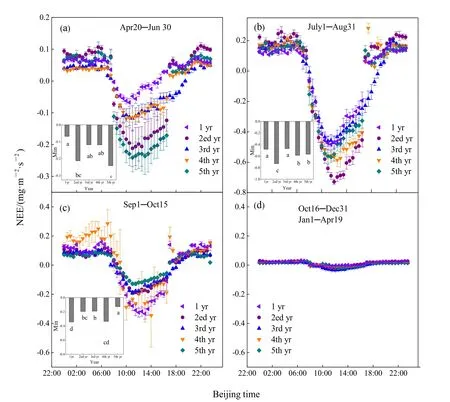
图4 一个种植周期人工草地不同季节净生态系统交换(NEE)有效数据的平均日变化趋势Fig. 4 Average diurnal trends of each season in net ecosystem exchange (NEE) over a plant cycle in the artificial grassland
2.2.3 NEE的年际变化
由图6a可知,GPP、R和NEE随着种植年限的增加呈现先增加后降低的趋势,在第3年峰值达到最大,随后降低()。除了种植第一年,其他年份GPP均高于R,第3年GPP远高于R,因此碳汇强度最强,为-128.52 g·m-2,而种植第一年为 47 g·m-2,是一个弱的碳源。总体而言,该生态系统在一个种植周期内为碳汇,累计固碳180.4 g·m-2。
2.2.4 R/GPP
由图 6b可知,不同种植年份在同一月中,R/GPP比值存在明显差异。总体而言,5-9月,R/GPP比值整体呈先减小后增加的趋势,减小的幅度大于增加的幅度。1yr,5月,GPP为0,因而没有R/GPP比值,除了6月,7-9月比值均小于1,说明7-9月GPP>R;2ed yr,5-9月R/GPP比值先减小后增加的趋势很明显,5月和9月R/GPP>1,6-8月R/GPP<1。3rd yr,5-9月R/GPP比值均小于1,整个生长季GPP始终大于R。4th yr和5th yr,R/GPP比值亦呈先减小后增加的趋势,4th yr比值在5月大于1,而5th yr小于1,在6-8月该比值小于1,到9月均大于1。在5月2ed yr R/GPP比值远高于其他年份,而在6月1yr该比值出现一个极值,远高于其他年份。
2.3 CO2通量季节变异的因子分析
2.3.1 生长季节生物和环境因子对CO2通量的影响
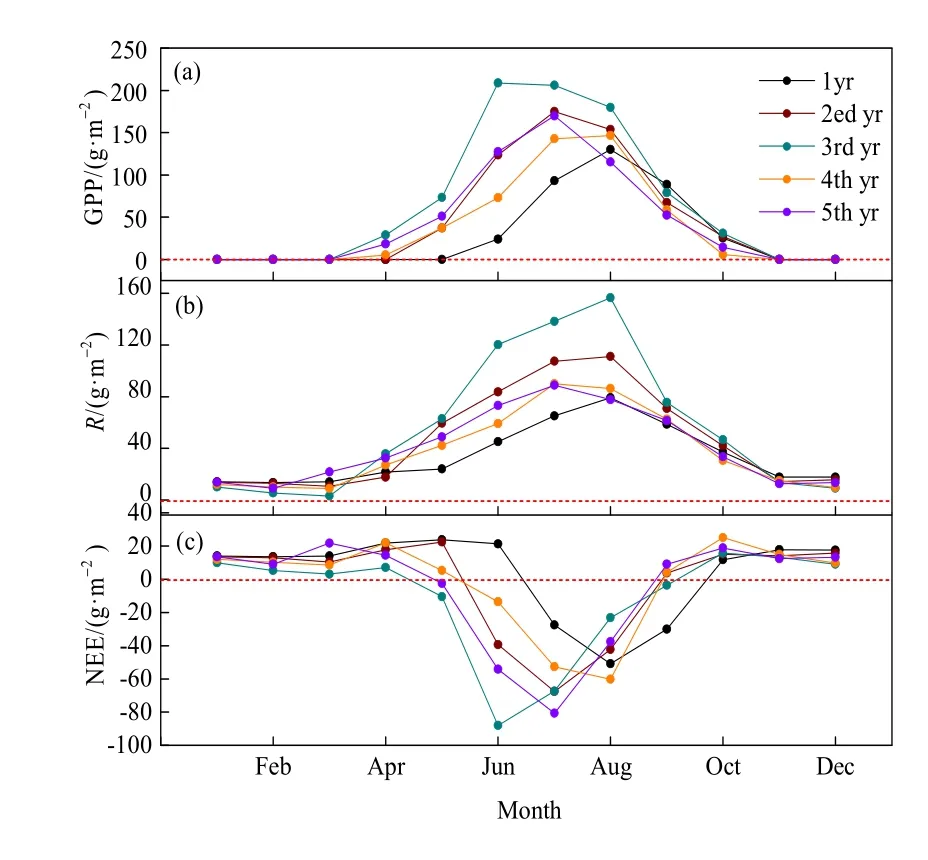
图5 一个种植周期人工草地生态系统总初级生产力(GPP,a),生态系统呼(R,b)和生态系统净CO2交换量(NEE,c)的季节变化Fig. 5 Seasonal changes in gross primary productivity (GPP, a),ecosystem respiration (R, b) and net ecosystem exchange (NEE, c) of artificial grassland ecosystems over a plant cycle
逐步回归分析表明,GPP主要受空气温度ta、植被多样性指数 SP和生长季长度(GSL)共同控制(r2=0.92,P=0.008,表2)。生长季R约占全年R的 81.2%,主要受土壤温度 ts控制(R2=0.80,P=0.025,表2)。而NEE与植被多样性指数SP呈显著的正相关关系(r2=0.8,P=0.026,表2)。
NEE与日均温呈正相关(r2=0.65,P=0.001,图7a),随着日均温的升高,CO2吸收量明显增加。经过5年的对比发现,在种植第二年NEE对温度的响应最强烈(斜率K最大),随着种植年限延长,NEE对温度的响应趋于平缓(K值趋于平缓)(图7b)。研究区降水量主要集中在5-9月,占全年降水量的87.78%,通过分析月降水量与NEE关系发现,NEE与月降水量呈正相关(r2=0.39,P=0.012,图7c),随着降水量的增加,碳吸收能力增强。
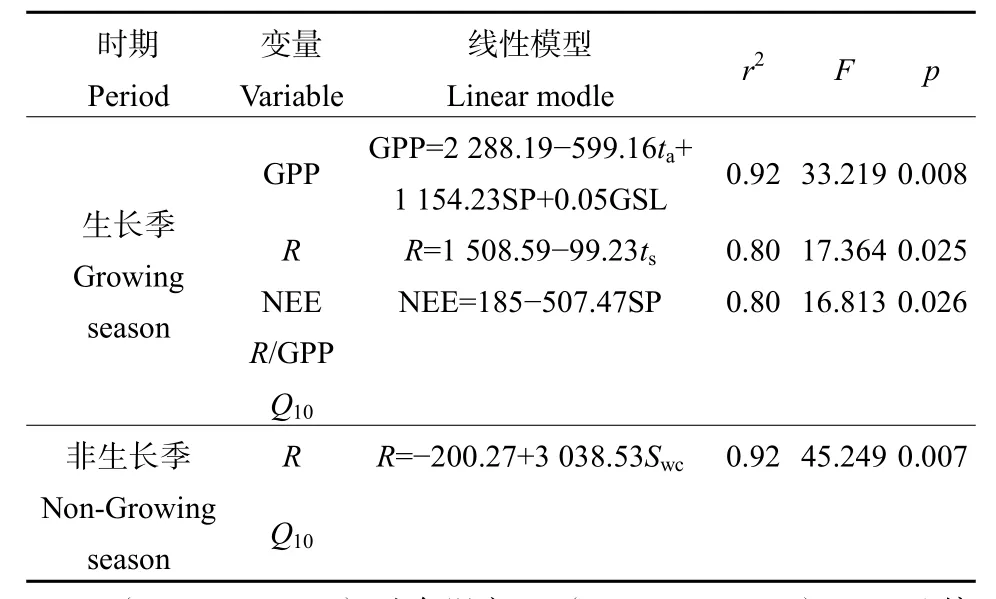
表2 多元逐步回归分析Table 2 Multiple stepwise regression analysis
2.3.2 非生长季节环境因子对CO2通量的控制
在非生长季节,人工草地生态系统没有光合作用,只有呼吸作用,NEE只是生态系统呼吸作用释放的CO2量,非生长季R占全年R的18.8%。根据逐步回归分析结果,非生长季R主要受土壤含水量Swc的控制(r2=0.92,P=0.007,表 2)。
2.3.3 年际NEE的影响因子
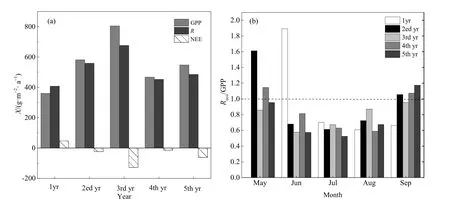
图6 一个种植周期总初级生产力(GPP),生态系统呼(R)和生态系统净CO2交换量 (NEE)年际变化(a)和5-9月各月生态系统呼吸与总初级生产力比值(R/GPP)变化(b)Fig. 6 Annual changes in gross primary productivity (GPP), ecosystem respiration (R) and net exchange ecosystems (NEE) (a) and variation of ecosystem respiration (R)/gross primary production (GPP) for each month from May to September (b) in a plant cycle
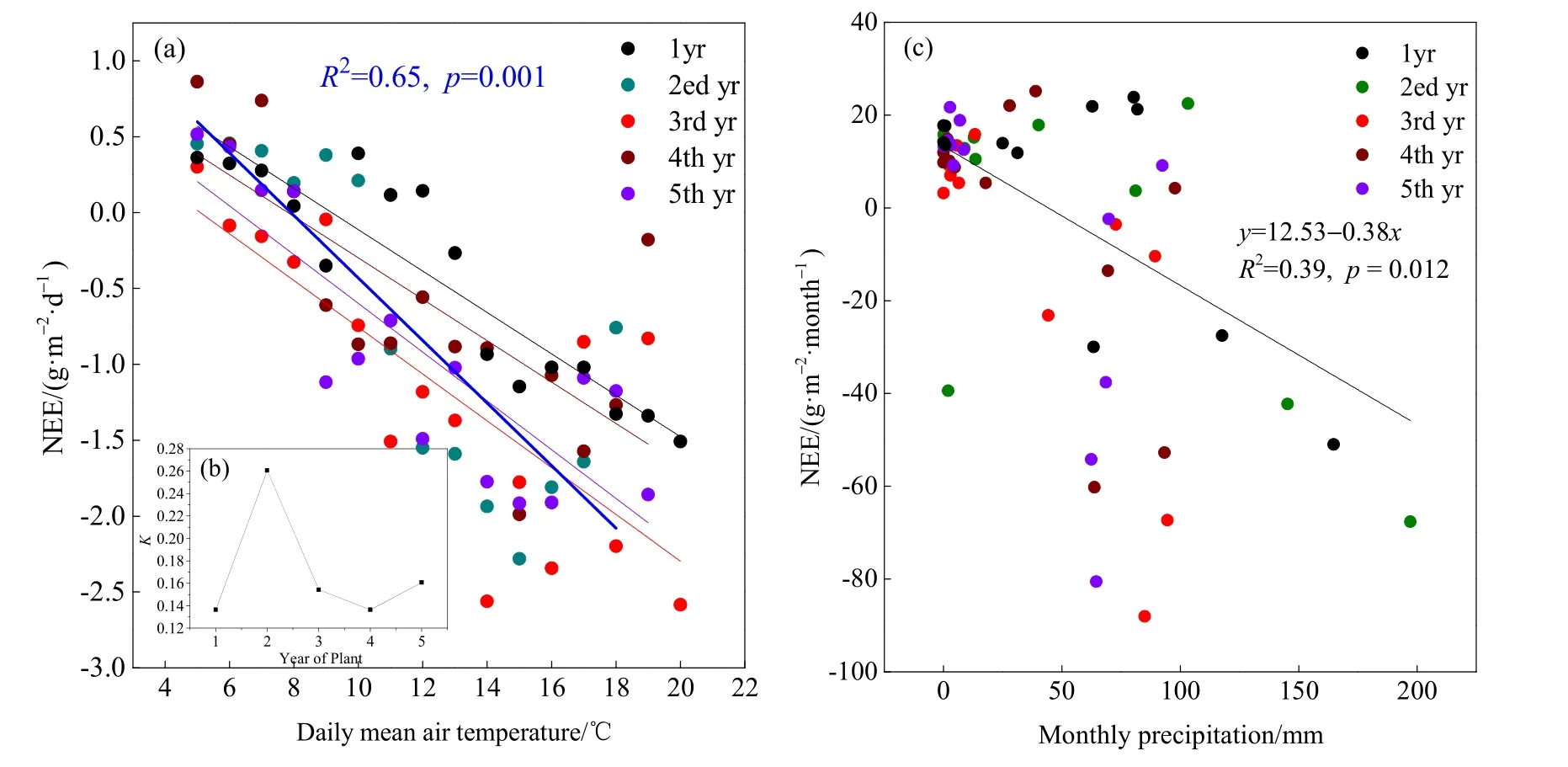
图7 日均温-NEE(a)、种植年份-温度响应斜率(b)以及NEE-月降水量(c)之间的关系Fig. 7 Relationship between day of NEE with day of temperature (a), temperature response slope and year of plant(b), monthly NEE with monthly precipitation (c)
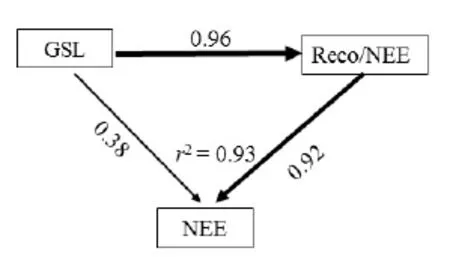
图8 生长季长度(GSL)对非生长季呼吸与生长季净生态系统CO2交换比值(R/NEE)对年际NEE影响的结构方程模型Fig. 8 Structure equation modeling of the relationship between the growing season length (GSL), non-growing season R and growing season NEE (R/NEE) and annual NEE of the artificial grassland
结构方程模型可以用来确定 NEE及其主要控制因子和相对贡献量。相关分析表明,MAT、MAP、AGB和SP与NEE的年际变异关系不显著。发现生长季长度(GSL)和非生长季呼吸与生长季净生态系统CO2交换比值(R/NEE)共同控制着年际NEE的变化(图8)。GSL通过影响R/NEE对NEE产生的间接影响(0.92)大于对其产生的直接影响(0.38),GSL和R/NEE共同解释了NEE年际变异的93%。
3 讨论
3.1 NEE的日变化
三江源农牧交错区多年生垂穗披碱草人工草地的 NEE日变化规律与已报道的青藏高原天然草地,如高寒矮嵩草(Kobresia humilis)草甸(Kato et al.,2006)、高寒金露梅(Potentilla fruticosa)灌丛草甸(Zhao et al.,2006)、高寒藏嵩草(Kobresia tibetica )沼泽化草甸( Zhao et al.,2010;Niu et al.,2017;张法伟等,2008)和多年生垂穗披碱草人工草地(赵亮等,2008;吴力博等,2010)的变化规律是一致的,这是由于光照的日变化、植物对大气CO2的吸收和土壤CO2排放的季节变化不同,导致NEE的日变化在不同季节的变化差异明显(Kato et al.,2004;Zhao et al.,2005a)。然而,本研究发现三江源农牧交错区多年生垂穗披碱草人工草地NEE存在明显的年际差异。在返青期,5th yr的CO2最大吸收速率是1yr的3.4倍,由于1yr种植时间较晚,差异较大。而在生长季旺期2ed yr的吸收峰值是3rd yr的1.6倍,高于沼泽化草甸的变异系数(1.2)(Niu et al.,2017)。枯黄期1yr的吸收峰值是5th yr的2.6倍,因为1yr不收割,其他年份均要收割,1yr夜间 CO2最大排放速率达 0.16 mg·m-2·s-1,远高于其他年份,是 5th yr (0.09 mg CO2·m-2·s-1)的1.7倍。3-5年的人工草地群落结构单一,且不稳定(Xu et al.,2017),可能5th yr植物群落处于退化的状态,导致吸收和释放速率较低。
人工草地CO2的昼夜最大吸收速率和最大排放速率都在生长季旺期,分别为 0.47-0.73 mg·m-2·s-1和 0.17-0.28 mg·m-2·s-1,高于青藏高原东南部的人工草地白天最大吸收速率0.35 mg·m-2·s-1,和夜间最大释放速率 0.22 mg·m-2·s-1(吴力博等,2010)。与青藏高原的天然草地相比,最大吸收率高于矮嵩草草甸 0.37 mg·m-2·s-1,金露梅灌丛草甸(0.36-0.48) mg·m-2·s-1和沼泽化草甸0.48 mg·m-2·s-1(Zhao et al.,2005a)。但低于青藏高原中部的沼泽化草甸白天最大吸收速率0.73-0.9 mg·m-2·s-1和夜间最大排放速率 0.20-0.29 mg·m-2·s-1(Niu et al.,2017)。因植被类型、环境因子不同,造成日出后植物产生光合现象时间的不一致性,植物对大气中CO2的吸收和释放时间均有所不同。
3.2 NEE的季节变化
最新研究表明,对土地进行有效管理有助于CO2的季节性变化(Zeng et al.,2014),青藏高原草甸生态系统地-气CO2的NEE具有明显的季节变化。这一特征在人工草地生态系统中表现得更加明显。在不考虑环境因子的变化下,随着种植年龄的增加,人工草地生态系统开始固碳的时间提前,1yr开始固碳时间最晚(7月初),其次为2ed yr,3rd yr最早,4、5th yr较为滞后,除了1yr,其他年份均在5月就开始碳固定,与高寒矮嵩草草甸生态系统相近(5月中旬)(Zhao et al.,2005a),早于高寒金露梅灌丛草甸(6月上旬)(柴曦等,2018)和高寒藏嵩草沼泽化湿地(7月上旬)(Zhao et al.,2005a)。1yr在 6月种植,故当年开始固碳的时间明显晚于其他年份。由于收割,2-4th yr在9月初停止固碳而开始碳排放,与高寒藏嵩草沼泽化湿地相近(9月上旬)(Zhao et al.,2005a),早于高寒金露梅灌丛草甸(9月中旬)(Zhao et al.,2005a),1yr(不收割)在9月底才转为碳排放,与高寒矮嵩草草甸相近(9月下旬)(Zhao et al.,2005a)。由此可以看出,对草地的管理直接影响着碳源汇(源→汇→源)的转换时间。
3.3 NEE年际变化
不同草地生态系统年际 NEE差异的原因是比较复杂的,这主要是因为它受到草地植被类型、生物环境因子以及草地管理策略(施肥、刈割和放牧等)等多方面因素的影响(Jensen et al.,2017)。对于人工管理的生态系统而言,管理方式(播种,施肥和刈割)可能是控制生态系统NEE差异的主要因子。我们发现,人工草地生态系统NEE年际变异主要受 GSL调控,GSL通过影响R/NEE间接影响 NEE(图 8)。这与最近研究结果类似,高寒金露梅灌丛草甸10年的数据表明,NEE年际变异主要受GSL的控制(Li et al.,2016)。不同的是,人工草地生长季受人为干扰较强,6月播种(1yr),开始固碳的时间明显晚于其他年份,相比高寒金露梅灌丛草甸、草原化草甸(柴曦等,2018),开始固碳时间晚了近1-2个月,从而缩短了生长季和碳吸收时期。而且翻耕使得大量土壤中封存的碳裸露在空气中,加快了土壤碳的分解,增加了土壤呼吸,使得R高于GPP,故种植第一年表现为碳源,其他年份GPP均高于R,表现为碳汇。除了1yr,其他年份8月底收割牧草,提前结束了生长季和碳吸收,相比与天然草地,人工草地的生长季长度和碳吸收时期明显短于高寒灌丛和草甸生态系统1-2个月,牧草收割后,停止生长,而呼吸依旧进行,加上 9-10月水热依然充沛,从而加速了生态系统的呼吸,抵消了之前大量的碳吸收。
3.4 CO2通量控制因子
在众多生物因子和非生物因子中,ta、SP和GSL直接影响着GPP变异,这与青藏高原天然草地研究不同,ts和光合有效辐射直接影响着海北高寒金露梅灌丛草甸生态系统GPP的年际变异(Li et al.,2016),而在当雄高寒草原化草甸生态系统中,ts和Swc是控制GPP的主要因子(柴曦等,2018)。本研究既考虑了生物因子植被的多样性特征,又考虑了环境因子和人为干扰的生长季长度GSL,从多角度解释了GPP的季节变异。植被多样性指数作为表征生物群落结构的重要指数,在群落结构相对稳定的天然草地中难以被定量分析,而建植5年的人工草地群落处于不稳定状态,植被多样性指数随着种植年限增加而变化。本研究表明植被多样性指数(SP)是影响生长季 GPP的重要生物因子。同时 SP直接影响着生长季NEE,随着植被多样性的增加,固碳能力逐渐增强,这一结果表明在人工草地中,群落的多样性直接影响着 CO2通量变化。长期研究发现,垂穗披碱草人工草地群落随着种植年限的增加,在种植 5年以后,优势种垂穗披碱草逐渐被入侵杂草取代,物种数先降低后增加(温军,2012),物种数和多样性指数随着种植年份的增加而变化明显,优势种不断发生变化,总光合能力也随之发生变化,导致碳交换通量发生变化。同位素标记研究表明,物种丰富度和根冠比决定着碳周转(Zhao et al.,2015),而且还表明物种组成和群落结构影响着生产力(Zou et al.,2014),由此可见,物种组成、群落结构和植物多样性与生态系统生产力和碳循环密切相关。在退化草地恢复和草地管理时考虑多种牧草混播的方式,可能对生态系统功能产生重大影响。
生长季R约占全年R的81.2%,低于高寒金露梅灌丛草甸的97%(Li et al.,2016)和高寒矮嵩草草甸的86.8%-88.2%(Wang et al.,2015)。本研究表明ts是影响生长季R的决定性因子,直接影响着生长季的R,这与高寒金露梅灌丛草甸生态系统中的结果是一致的(柴曦等,2018)。而非生长季R占全年R的18.8%,高于高寒金露梅灌丛草甸的7%(Li et al.,2016)和高寒矮嵩草草甸的11.8%-13.2%(Wang et al.,2015),可能是人工草地非生长季长度大于灌丛和草甸导致,加之翻耕引起的大量土壤碳排放,增加了非生长季的R。在非生长季中Swc是影响R的决定性因子,这一结果表明在三江源区非生长季R主要受土壤水分的影响,这可能与季节性冻融过程有关,最近研究也表明青藏高原草地非生长季土壤呼吸受冻融过程的控制(Wang et al.,2015)。可以看出人工草地的R,是草地碳循环的一个重要环节,R的季节变异主要受土壤温度和含水量的影响。
人工草地 NEE随着日均温的增加而增加(图7a)。最新研究表明温度是陆地NEE年际变异的主控因子(Jung et al.,2017),在寒冷的青藏高原,温度成为了驱动植被进行光合和呼吸作用的重要环境因子。经过5年的对比发现,2ed yr NEE对温度的响应最为强烈(斜率K为最大),随着种植年限的增加,其对温度的响应趋于平缓(图 7b),可能是因为该年降水量分配原因,生长季降水量集中,降水强度大(图 3c)。由于环境因子之间有一定的内在联系,共同影响着生态系统的碳收支。水分是影响生态系统碳收支的一个重要环境因子,该研究区年降水量主要集中在 5-9月,在此期间水分条件良好,因此,在生长季水分不会对生态系统构成胁迫。相关研究也表明青藏高原降水主要集中在生长季,不会对生态系统造成水分胁迫(李英年,2004;吴力博等,2010)。
4 结论
青藏高原垂穗披碱草人工草地生态系统是一个碳汇,一个种植周期累计固碳180.4 g·m-2(以C计)。草地管理措施(播种和收割)通过改变生长季季节长度影响 NEE的年际变异。随着种植年限的延长碳汇能力先增加后减小,在种植第3年碳汇能力达到最大。三江源区人工草地的建植可以增加草地生态系统的固碳能力。同时,本研究表明,种植 5-6年后,优势种垂穗披碱草逐渐被入侵杂草取代,生产效益低下,收益低于投入。不论从固碳功能还是生产功能,每 5-6年重新翻耕人工草地是“生态-生产”的双赢模式,人工草地更新是三江源农牧交错区一个生产和生态相结合的有效产业发展模式。因此,在三江源农牧交错区发展多年生草种产业,不但解决了青藏高原及其周边地区的退化草地治理中草种的需求,而且建植的人工草地可增加碳封存量。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
小猕猴智力画刊(2021年11期)2021-11-28
空间科学学报(2021年2期)2021-07-21
海外文摘·文学版(2021年3期)2021-05-19
活力(2019年21期)2019-04-01
儿童故事画报·自然探秘(2016年4期)2016-06-24
科学启蒙(2016年5期)2016-05-10
中国学术期刊文摘(2016年1期)2016-02-13
中国学术期刊文摘(2016年8期)2016-02-13
