
西南喀斯特地区小叶黄杨与白刺花光合特性研究
2019-06-18 06:48周国富
中国饲料 2019年9期
吴 强, 李 倩, 肖 华, 周国富*
(1.贵州师范大学地理与环境科学学院,贵州贵阳 550001;2.贵州省水利投资(集团)有限责任公司,贵州贵阳550081;3.贵州师范大学喀斯特研究院,贵州贵阳550001)
光合作用对植物至关重要,植物90%以上的干物质都来自叶片的光合同化作用,利用好植物的光合特性对其产量和品质提升都具有意义 (樊卫国等,2006)。植物叶片的光合特性与叶片结构和环境密切相关,同时对不同的生境条件会表现出不同的适应机制与适应特性 (Robert等,2007;Ueda等,2000)。喀斯特生态环境系统是严重受喀斯特环境制约的生态系统(熊康宁等,2015)。中国南方喀斯特地处热带、亚热带雨水非常丰富,但受特殊地质环境影响,形成一种特殊的喀斯特干旱环境,植物常表现出特有的旱生性、石生性和喜钙性(Lee 等,2013;曹建华,2005;袁道先等,1988)。因此喀斯特石漠化地区发展饲用林要充分考虑经济林木的光合特征,充分利用光合特点提升质量。
小叶黄杨(Buxus microphylla)属黄杨科黄杨属,是我国南方常见的树种,同时营养含量较高适合黑山羊等动物采食 (孔德顺等,2014)。白刺花(Sophora davidii)属豆科槐属,是南方喀斯特地区广泛分布的旱生灌丛群落优势种或广布种(Khairun 等,2016;李芳兰,2007)。有研究表明,两种林木饲用价值高,抗旱性能良好,同时在喀斯特山区植物群落演替、物种多样性维持、土壤改良与防治水土流失、医药研究等方面有着重要生态经济价值,有望成为喀斯特干旱、贫瘠山区植被恢复和发展畜牧业的理想材料(李芳兰等,2006;王海珍等,2005)。因而深入揭示小叶黄杨和白刺花的光合特性,对其饲用资源开发以及喀斯特山区植被恢复都具有重要的理论与实践指导意义。
1 研究区自然概况
研究区位于贵州省毕节市七星关区撒拉溪国家石漠化综合治理示范区 (105°02′01′′-105°08′09′′E,27°11′36′′-27°16′51′′N),属于典型的喀斯特高原山地潜在-轻度石漠化地区,平均海拔 1600 m。主要地貌类型为高中山地,基岩裸露面积大,水土流失严重。主要土壤类型为黄壤。耕地多分布于坡面、台地和山间谷地,耕层浅薄、肥力较低。该区属亚热带季风湿润气候,年均温约为12℃,无霜期245 d,年均日照时数为1360 h,多年平均降水量984.4 mm,降水季节分布不均,80%以上的降水集中在6~9月,降水多由岩隙渗入地下,区域内生活及农业用水困难。整个研究区因为季节性降水不均以及喀斯特地貌发育漏水严重,可利用水资源相对匮乏。
2 材料与方法
试验材料为3年生小叶黄杨、白刺花。2014年9月,选择晴天进行光合测定田间试验,在8:00、10:00、12:00、14:00、16:00、18:00 时段内,取中间时段每隔2 h测定1次,共测6次,小叶黄杨和白刺花各随机选取15株,每株选取植株3片中上部成熟向阳叶片进行试验,取平均值计算。光合测定仪器为英国ADC Bioscientific公司生产的Lcpro+便携式光合测定仪,使用红蓝光源叶室测定叶片瞬时光合速率等指标。主要测定和统计指标包括:净光合速率(Pn,μmolCO2·m-2·s-1)、蒸腾速率 (Tr,mmolH2O·m-2·s-1)、CO2气孔导度(Gs,molH2O·m-2·s-1)、细胞间 CO2浓度(Ci,μmol·mol-1)、大气温度 [叶室温度 (Tch,℃)]和叶片温度(Tleaf,℃)、光合有效辐射(PAR,μmol·m-2·s-1)、水分利用效率 (WUE,μmolCO2·mmol-1H2O)。 采用SPSS 20、Microsoft Excel 2007软件进行数据统计分析,OriginPro 9.0软件进行绘图。
水分利用效率/(μmolCO2·mmol-1H2O)=净光合速率/蒸腾速率。
3 结果与分析
3.1 大气参数日变化特征 如图1所示,试验地光合有效辐射实测值在8:00~18:00的日变化为 189.31 ~ 1685.40 μmol·m-2·s-1,8:00 左 右 为(888.53 ±76.53)μmol·m-2·s-1,12:00 达 到 峰 值(1685.40±97.63)μmol·m-2·s-1,一天中最低值出现在 18:00,为(189.31±47.32)μmol·m-2·s-1,光合有效辐射日均值为(1177.78±592.16)μmol·m-2·s-1。
叶室气温主要受光合有效辐射影响以及部分叶室的遮挡,由图1可知,其变化为25.38~36.64 ℃,8:00 叶室气温最低为 (25.385±1.29)℃,此后,由于光合有效辐射的迅速增加而上升较快,到14:00左右叶室气温达到最高值(36.64±0.66)℃, 叶室平均气温为 (32.36±4.36)℃。叶片的温度受光合有效辐射影响较叶室温度高1℃左右,其变化为26.27~38.14℃,同样最低温出现在 8:00,为(26.27±1.22)℃,最高温出现在 14:00,为(38.14±0.67)℃,叶片平均气温为(33.50±4.73)℃。

图1 主要环境因子日动态变化
试验日大气压变化为882.86×104~886.02×104Pa,最高大气压出现在 10:00 ~ 12:00,为(886.02±0.10)×104Pa,最低值出现在 18:00,为(881.98±0.10)×104Pa,日均大气压为(884.11±1.54)×104Pa。
3.2 净光合速率日变化特征 如图2所示,小叶黄杨和白刺花的净光合速率日均值分别为(2.66±0.69)、(7.32±3.87)μmolCO2·m-2·s-1, 白刺花的日均净光合速率高于小叶黄杨。小叶黄杨和白刺花的净光合速率日变化曲线差异较大,小叶黄杨的净光合速率日变化曲线呈现倒“L”型,在8:00出现最大值,此后一直呈平缓下降趋势,在16:00之后又开始增长。 而白刺花在 8:00~12:00保持较高的水平,在12:00出现当日最大值,此后一直呈下降趋势,在18:00出现最低值。结果表明,小叶黄杨对光照较为敏感,且受高光照辐射影响较为严重,出现长时间“午休”现象,白刺花则表现为12:00之前高净光合速率,之后低净光合速率的特征,这可能主要是受高光照辐射的影响。

图2 净光合速率日动态变化
3.3 蒸腾速率日变化特征 如图3所示,小叶黄杨和白刺花的蒸腾速率日均值分别为 (1.64±0.97)、(4.77±1.34)mmolH2O·m-2·s-1, 小叶黄杨的日均蒸腾速率明显低于白刺花。小叶黄杨和白刺花的蒸腾速率日变化在8:00基本一致,此后曲线变化呈现负相关。小叶黄杨在8:00出现最大值,8:00到10:00下降较快,此后曲线变化相对比较平稳。白刺花在8:00到10:00曲线上升较快,在10:00出现最大值,10:00之后一直呈下降趋势。从小叶黄杨和白刺花的蒸腾速率曲线来看,小叶黄杨受高光辐射、高温影响,蒸腾速率明显降低,而白刺花蒸腾速率受高光辐射、高温影响没有小叶黄杨剧烈。
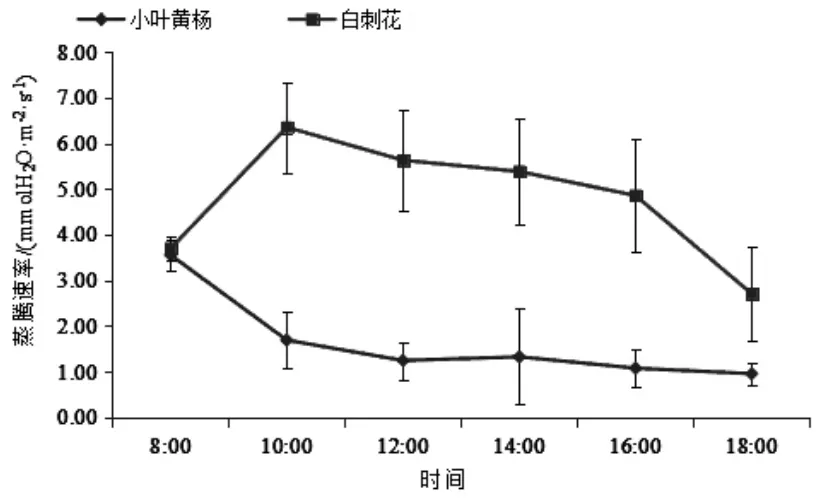
图3 蒸腾速率日动态变化
3.4 水分利用效率日变化特征 水分利用效率的大小一般可以反映植物适应能力的强弱与水分利用性能(Denis等,2005)。由图 4可知,小叶黄杨和白刺花的水分利用效率日均值分别为 (1.40±0.78)、(1.52±0.84)μmolCO2·mmol-1H2O。小叶黄杨和白刺花的水分利用效率日变化曲线具有一定的相似性,但也存在不少差异,日均值相差不大。小叶黄杨水分利用效率在8:00最高,超过白刺花,在 10:00~14:00呈缓慢上升趋势, 在 16:00出现一日内最低值,之后又缓慢上升。白刺花水分利用效率在8:00出现最大值,在10:00出现一个低值,12:00出现一个次峰值,在16:00略有上升,变化不大。

图4 水分利用效率日动态变化
3.5 气孔导度日变化特征 如图5所示,小叶黄杨和白刺花的气孔导度曲线均为倒“L”型,两条曲线走势较为相似,均在8:00出现最大值,10:00后变化幅度均较小,曲线平稳。小叶黄杨和白刺花的气孔导度日均值分别为(0.09±0.17)、(0.20±0.17)molH2O·m-2·s-1,小叶黄杨气孔导度各观测点均低于白刺花。

图5 气孔导度日动态变化
3.6 胞间CO2浓度日变化特征 如图6所示,小叶黄杨和白刺花的胞间CO2浓度日均值分别为(80.39±32.13)、(275.38±55.70)μmol·mol-1。 小叶黄杨的胞间CO2浓度曲线变化较为平缓,胞间CO2浓度值在8:00出现最大值,在14:00出现最低值,此后缓慢上升。白刺花的胞间CO2浓度曲线两边高中间低,最低值出现在12:00,比小叶黄杨最低值提前2 h;白刺花胞间CO2浓度最大值出现在18:00,次峰值出现在8:00,从总体上看两者曲线变化趋势较为相似。
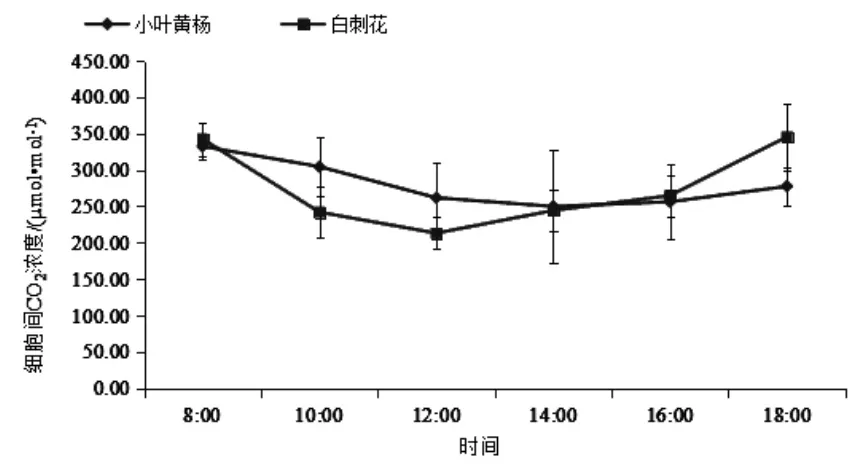
图6 胞间CO2浓度日动态变化
3.7 各因子的相关性 通常各因子的相关性多采取综合均值分析,具有直观简单的特点,但也存在某个节点分析不够明确的情况。为此,本文将各个光合因子在6个时间节点分别分析,更有利于不同时间节点光合植物的利用。在8:00、10:00小叶黄杨的净光合速率与温度极显著相关 (P≤0.01),说明温度是影响净光合速率的重要因素;在 8:00、12:00、16:00 小叶黄杨的蒸腾速率与温度显著(P≤ 0.05)或极显著(P≤ 0.01)相关,说明这些时间节点,温度因素影响十分显著。各个节点综合分析可知道,小叶黄杨的净光合速率与蒸腾速率、气孔导度极显著相关(P≤0.01),与细胞间CO2浓度显著相关(P≤ 0.05);蒸腾速率与细胞间CO2浓度、气孔导度显著相关(P≤ 0.05)。详见表1。
在8:00白刺花的净光合速率与除蒸腾速率外的各项因子均呈极显著相关 (P≤0.01)或显著相关(P≤ 0.05),说明各项因子均对净光合速率产生影响;在10:00、12:00白刺花的净光合速率与蒸腾速率极显著相关 (P≤ 0.01),14:00显著相关。各个节点综合分析可知道,白刺花的蒸腾速率与光合有效辐射、温度、胞间CO2浓度显著相关(P≤ 0.05)。从相关性关系上可知,叶片温度较叶室温度更接近实际值。详见表 2。
4 结论与讨论
林木的光合作用主要通过净光合速率和蒸腾速率两个生理指标表现出来,其中光合速率的强弱是植物积累物质多少的表现,蒸腾速率高低是植物控制利用水分能力强弱的表现,水分利用效率则是植物水分消耗和同化利用能力的主要评价指标(Monteiro 等,2016;Hidayati等,2016;池永宽等,2015;蒋赕等,2013)。同时,林木的光合作用与生长环境还存在密切的联系,林木通过改变自身代谢机制来适应不利环境,在长时间的适应过程中形成匹配当地环境的光合机制,以完成正常的生命周期(Farber等,2016)。植物叶片受温度和光照有效辐射的影响,形成不同的细胞间CO2浓度和气孔导度调节机制,影响光合速率不断变化。林木的蒸腾速率高低是评价其运导水分能力的重要指标,植物体内物质的积累需要水分以及通过水分运载的矿物质营养成分的不断供应 (Chi等,2016)。随着光照辐射增强、气温升高,导致植物出于保护自身不至于脱水,主动或被动的关闭部分气孔,造成光合作用原料即胞间CO2的浓度降低,叶片水分减少,使光合速率和蒸腾速率相对下降,变化趋势出现一个较为明显或滞后的 “午休”现象。
小叶黄杨和白刺花是西南喀斯特地区具有重要潜在价值经济林木,分布广泛,但又分属于不同科属,光合特性存在较大差异。小叶黄杨的日均净光合速率为(2.66±0.69)μmolCO2·m-2·s-1,远低于白刺花的(7.32±3.87)μmolCO2·m-2·s-1,这基本符合豆科植物综合光合效率高于黄杨科的规律(Xiong等,2017)。总体观察小叶黄杨光合速率最活跃的时间为 8:00,(10.48±2.12)μmolCO2·m-2·s-1,蒸腾速率最高值也是出现在了 8:00,(3.55±0.34)mmolH2O·m-2·s-1,可知小叶黄杨光合生理特性最活跃时间是在8:00左右,其他时间段较低,可能是其不耐强光和高温的原因。白刺花的光合特性则表现出另一种趋势,净光合速率在8:00~12:00 都是高光合特征,在 10 μmolCO2·m-2·s-1以上,12:00之后也是一直保持在相对较高状态,除18:00 外,均在 3 mmolH2O·m-2·s-1以上,白刺花的蒸腾速率也是相对高蒸腾状态,造成这种差异原因与两种植物科属不同的叶片生理结构有关,也与试验时间、试验地点和树龄等因素有关,有待进一步研究。
植物的水分利用效率高低主要取决于净光合速率与蒸腾速率(刘红明等,2018)。小叶黄杨和白刺花的水分利用效率最高值均出现在8:00,说明在8:00是植物同化作用最佳的时间之一。本试验中在其他时段小叶黄杨和白刺花的水分利用效率存在较大差异,但存在“高-低-高-低”的交叉变化趋势。整体上讲,小叶黄杨属于低光合速率、低蒸腾速率、低水分利用效率的特点;白刺花属于高光合速率、高蒸腾速率、低水分利用效率的特点,从日均值上白刺花[(1.40±0.78)μmolCO2·mmol-1H2O]略高于小叶黄杨 [(1.52±0.84)μmolCO2·mmol-1H2O]。在试验地长期野外观测结果也表明,白刺花长势要稍好于小叶黄杨。
综上所述,在石漠化这种特殊的环境背景下栽植饲用灌木林,特别是挂果期的时候要充分考虑不同时间节点的光合特点,采取必要措施,如遮阴、补水等手段,提高其水分利用效率,发掘潜在生态价值与饲用价值。结合小叶黄杨和白刺花的不同时间光合适应特点,对进一步的饲用林经济价值开发以及喀斯特山区植被恢复都具有重要的理论与实践指导意义。
猜你喜欢
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
科学(2020年3期)2020-11-26
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
乡村地理(2019年2期)2019-11-16
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
湖北农业科学(2014年13期)2014-08-28
文化月刊·下旬刊(2014年6期)2014-08-28
