
人工模拟增雨对唐古特白刺种子形态性状和萌发特性的影响
2019-06-11 06:13张景波辛智鸣刘明虎张冉浩黄雅茹
西南农业学报 2019年5期
张景波,董 雪,辛智鸣,3,刘明虎,张冉浩,黄雅茹,孙 非
(1.中国林业科学研究院荒漠化研究所, 北京 100091; 2中国林业科学研究院沙漠林业实验中心,内蒙古 磴口 015200; 3.内蒙古磴口荒漠生态系统国家定位观测研究站,内蒙古 磴口 015200)
【研究意义】种子形态是重要的植物生活史特性之一,主要包括种子重量、形状、附属物和表面结构等性状[1-2]。它与种子散布、休眠、萌发、出土、幼苗生长及种子库持久性等密切相关[3-6]。种子形态不仅制约着物种的分布和丰富度[6],而且影响植物群落演替过程[7-8],因此具有重要的适应意义[3, 9-11]。【前人研究进展】唐古特白刺(Nitrariatangutorum)是蒺藜科(Zygophyllaceae)白刺属匍匐性小灌木,抗旱,耐瘠薄,喜沙埋,拦阻并积聚风沙逐渐形成丘状沙堆,在保障干旱荒漠区生态安全中发挥着极其重要的作用,广泛分布在西北干旱区[12-16]。国内已经在干旱区对白刺进行了增雨实验,结果表明,甘肃省民勤县的增雨量达到当地年均降水量(115 mm)时即增雨100 %,白刺对增加水分的利用比例才能明显增加[17]。增雨同时可以使白刺的生长期延长,包括春季展叶期和花期提前,秋季落叶期延迟[18]。每增加100 mm水分白刺叶片长度增加0.52 cm、而宽度增加0.2 cm[19]。100 %增雨处理能够显著促进白刺当年生枝叶生长,提高生物量[20]。但目前对其种子形态如何响应降水增加却未涉及,限制了我们对唐古特白刺有性繁殖策略的认识。【本研究切入点】气候变化尤其是降水变化对生态系统的结构和功能有重要影响[21]。但降水变化是否影响种子的形态属性?至今鲜见报道。降水格局变化将对荒漠生态系统产生重要影响[22-23]。随着降水的增加,不同种子大小的物种丰富度都有增加,但是大种子的优势将降低,植株数目下降,小种子植物的数目极显著增加[24]。目前,学者们在群落调查基础上,探索种子的尺寸、种子的形状、种子的颜色、种子的有无光泽等这些性状与温度、降水、光照等生态因子之间的关系及原因做了一些研究[5,24,28]。研究表明:种子性状不仅随气候变化而变化,而且同一地区的不同坡向、不同演替阶段也影响种子特性[10]。但是有关模拟增雨对荒漠植物唐古特白刺种子形态及所造成对干旱环境的适应机制方面的变异未见报道。【拟解决的关键问题】本研究对自然生长的白刺沙包进行长期人工模拟增雨,测定唐古特白刺的种子形态性状,以期揭示它对模拟增雨的响应,促进唐古特白刺灌丛的保护,为荒漠化土地防治提供理论依据。
1 研究区概况
研究区位于内蒙古自治区巴彦淖尔市的磴口县中国林业科学研究院沙林中心第二实验场内,地理坐标为 E106°43′,N40°24′。该地区属于中温带大陆性季风气候,夏季炎热,冬季寒冷。降雨多集中在 6-9月,多年平均降水量约为145 mm;年均潜在蒸发量为2397 mm;年平均气温7.6 ℃,昼夜温差较大,生长季日温差可达15 ℃;全年平均相对湿度为48 %。年平均日照时数3210 h,占全年可照时数的72 %,无霜期在136~205 d;风沙危害是主要的自然灾害,年平均风速为3.4 m/s,最大风力可达18 m/s,平均大风日数为12.5 d。样地区域地势平坦,土壤类型为风沙土,地下水埋深在 3~4 m。试验区广泛分布着唐古特白刺,群落内伴生有少量沙蒿(Artemisiadesterorum)和沙鞭(Psammochloavillosa)等多年生植物以及虫实(Corispermumhyssopifolium)、沙米(Agriophyllumsquarrosum)、盐生草(Halogetonglomeratus)和猪毛菜(Salsolacollina)等一年生植物,群落盖度为30 %~40 %。
2 材料与方法
2.1 试验设计与种子采集
模拟增雨实验于2008年开始,每年的5-9月进行。以磴口县多年(1961-2000年)平均降水量 145 mm为依据设置 4个增雨梯度,即分别增加年降水量 25 %、50 %、75 %以及100 %,同时设置 0 %的增雨为对照样地(CK),每次增雨量分别为0、7.25、14.5、21.75和29 mm, 增雨时长约5 h。每个处理和对照都设置 4个重复,共 20个实验样地,样地之间的间隔在5 m以上,以尽量减少相邻样地之间的相互干扰。每个样地为 1个半径为 6 m的圆形,面积约为 113 m2。每个样地内有一个白刺沙包,沙包位于样地中间,其平均高度和面积分别为(1.24±0.14)m和(29.9±0.39)m2。除对照样地外,每个样地安装1套全光照喷雾灌溉系统,进行人工模拟增雨,增雨用水取自样地附近的水井,由输水管道输送到样地内。所有实验测定和样品采集都在增雨能够稳定覆盖的范围内进行,2017年7月上旬在每棵母树采集生长发育正常的新鲜果实,果实采收后搓去果肉,淘洗干净后晾干,风干至恒重,于0~4 ℃下保存。
2.2 种子性状测定
本研究中测定每颗种子的表型性状采用Winseedle种子和针叶图象分析系统软件。该软件可获得种子形态指标包括种长(平行子叶方向)、种宽(垂直子叶方向)、种子体积、种子表面积、种子长宽比等。种子形态指标测定以单株为单位,不同增雨处理的指标值均以该模拟增雨条件下各单株的均值来表示,以50粒种子为一个样本。千粒重用 1/10000 电子天平测定。每次随机取100粒种子,5次重复。
种子长宽比(L/W)=种子长度/种子宽度的比值
变异系数=性状标准差/性状平均值×100 %
2.3 唐古特白刺种子分级发芽试验
实验于2018年4月15日在中国林业科学研究院沙漠林业实验中心进行。将种子播种于16 cm×8 cm的塑料花盆内,把不同增雨处理0 %、25 %、50 %、75 %和100 %的种子分别划分成特大粒、大粒、中粒、小粒和特小粒5个等级,每盆30粒,6个重复,播种深0.5 cm,播种后每天记录出苗情况,18 d后,将盆内沙土全部倒出,仔细核对幼苗的数量,同时记录花盆内萌发但未出土的种子数量。
2.4 数据处理
采用 Excel 2003和SPSS 17.0软件将各性状进行方差分析、差异显著性检验、相关性分析。采用回归分析种子形态指标(种长、种宽、种子体积、种子表面积、种子长宽比和千粒重)随着模拟增雨的变异趋势,以增雨量为自变量,各性状分别为因变量,作多项式回归分析。
3 结果与分析
3.1 试验样地2008-2017年天然降水量
由图1可知,研究区2008-2017年自然年平均降雨量为128.54 mm,其中年降雨量和生长季5-9月最大降雨量出现在2012年,分别为219.5和218.2 mm,而年降雨量和生长季最小降雨量出现在2017年,分别为86.0和54.5 mm,由2008-2017年降雨数据分析可知生长季降雨量占全年降雨量的85.36 %。由此可以看出干旱地区降水量具有极大的波动性和不确定性,唐古特白刺种子表型特征变化是在以上天然降水背景条件下进行测定的。
3.2 增雨对唐古特白刺种子性状变异特征
从表1可以看出,不同增雨条件下,种子长度的变异范围为5.710~6.753 mm,变异系数为 6.313 %。其中0 %处理的种子长度最大,且与50 %、75 %和100 %增雨处理差异性显著(P<0.05)。100 %增雨处理后种子长度最小且与其它4种处理均显著(P<0.05)。种子宽度的变异范围为3.095~3.580 mm,变异系数为5.868 %。其中0 %处理的种子宽度最大,且与其它四种增雨处理差异性均显著(P<0.05)。种子千粒重、体积和表面积的变异系数分别为20.386 %、17.239 %和11.488 %,且各处理间差异性均较显著。种长/种宽的变异系数为5.650 %。
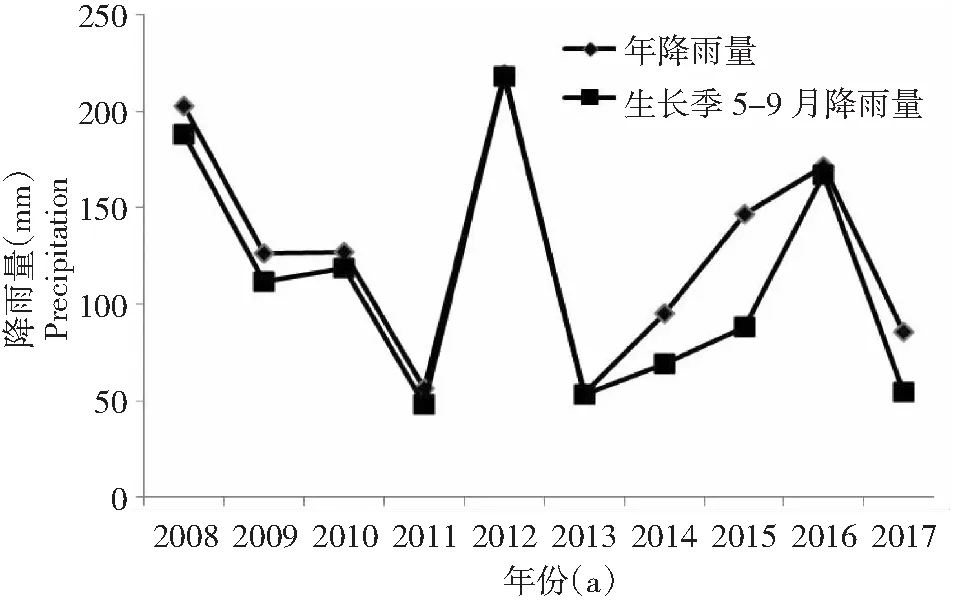
图1 样地2008-2017年天然降雨量Fig.1 The natural rainfall of plots from 2008 to 2017
3.3 唐古特白刺种子性状指标间的相关性
从表2可以看出:种长与种子表面积呈极显著正相关(P<0.01), 相关系数为0.941,说明种子长度对种子表面积的贡献较大。种子长度与种子体积和千粒重呈显著正相关(P<0.05)。种子宽度与种子体积呈显著正相关(P<0.05)。种子千粒重与种子体积和表面积均呈极显著正相关(P<0.01)。种子体积与表面积也呈极显著正相关(P<0.01),且相关系数最大为0.983。种子长宽比与其它各指标的间的相关性均不显著(P>0.05)。唐古特白刺种子各指标间的相关性大部分都达到了显著水平,说明种子形态指标之间存有复杂的联系,而且不是独立的。
3.4 唐古特白刺种子性状指标与增雨处理的相关性
唐古特白刺种子受增雨的影响,其形态性状在表达上呈现梯度性变异(图2)。种子各性状与模拟增雨因子的相关性分析表明,唐古特白刺种子千粒重、体积和表面积与增雨量均呈极显著负相关(P<0.01),相关系数依次为0.692、0.777、0.657。种子长度与增雨量呈显著负相关(P<0.05), 相关系数为0.480。种子宽度和长宽比与增雨量的相关性均不显著(P>0.05)。增雨处理是种子性状指标变异的缘由,越干旱地区的种子相对瘦长,这是对干旱条件适应性的响应。

表1 不同增雨处理唐古特白刺种子的形态指标(平均值±标准差)

表2 唐古特白刺种子形态指标相关性
注:*,P<0.05; **,P<0.01。

图2 种子性状与增雨处理的相关性Fig.2 Correlation coefficients between seed characters of Nitraria tangutorum and simulated precipitation

图3 不同大小唐古特白刺种子的出苗率Fig.3 Seed rate of Nitraria tangutorum different size seeds
3.5 唐古特白刺种子大小变异对出苗的影响
如图3所示,出苗前期即播种5~7 d后,小种子有较大优势,发芽较早,100 %处理的种子(特小粒),即小种子出苗迅速,且前期出苗率明显高于大种子(0 %处理特大粒和25 %处理大粒);出苗后期即播种第9天后,大种子发芽晚,但具有较高的发芽率,总发芽率和出苗率表现为:0 %处理(特大粒)>25 %处理(大粒)>50 %处理(中粒)>75 %处理(小粒)>100 %处理(特小粒),各处理间大小种子出苗率逐渐平缓,但大种子和小种子的总体出苗趋势相似。计算结果显示,大小种子的萌发率均在40 %左右,大种子的萌发率略高,但各处理间大小种子萌发率不存在显著差异(P>0.05)。各处理0 %、25 %、50 %、75 %和100 %种子由大到小种子的出苗率分别为43 %、40 %、38 %、35 %和32 %,大种子出苗率高于小种子,但方差分析表明,出苗率之间没有显著差异(P>0.05)。这说明无论种子大小,在其它因素一致的条件下,唐古特白刺种子的萌发与出苗情况基本是一致的。
4 讨 论
种子性状是植物相对稳定的生活史特征之一,但为适应复杂多变的环境会产生适应的变异[25]。不同增雨量处理对唐古特白刺种子形态指标均造成显著影响,在一定程度上反映了唐古特白刺对降水变化比较敏感。种子千粒重、体积和表面积变异幅度较大,说明这3个种子形态指标受增雨量的影响较大,而种子的长度/宽度的比值变异最小,说明种子长宽比是植物对自然环境长期适应、相对稳定的指标。唐古特白刺种子长度、宽度、体积、表面积、长宽比及千粒重在不同增雨处理间差异均为极显著水平,说明种子性状表现出来的差异并不是随机性的,而是与种子形态对不同增雨处理的响应密切相关。唐古特白刺种子主要借助于动物和重力传播,其体积、表面积及千粒重的变化可能造成种子传播能力的变化,进而影响种群格局和群落结构。种子千粒重的变化可能造成出苗率及幼苗生长状态的变化[26]。增雨处理对种子体积影响最大。体积是反映种子大小的指标,体积越大即吸水的表面积越大,其内含的营养物质越丰富,可以提供发芽的物质越多,使发芽迅速整齐,进而影响到幼苗乃至后期的生长。不同增雨处理间种子体积差异越大,在一定程度上说明了唐古特白刺在育种选择上的潜力较大,为唐古特白刺的优良群体选择提供了基础和条件。
大多数旱生植物长期生活在干旱生境中,限制其生长的主要因子是水分,带有较少贮存物质的小种子遇到环境中的各种萌发暗示后会快速萌发。随着降水的增加,唐古特白刺的生存环境改善,小种子的幼苗死亡率降低,并利用先萌发的优势迅速成长,使大种子的竞争优势降低,从而拥有更多的成株数目,产生更多的后代。所以,随着环境的改善,大种子的优势将逐渐降低。大小种子在分配于繁殖的能量相同的情况下,小种子的数目会比大种子多很多,依靠风力扩散的能力强,距离也较远,所以能够充分利用随机降水,迅速萌发产生更多的幼苗。在降水稀少的地区,大种子更能适应恶劣的环境,使幼苗在短时间内不需要对外界环境的依赖,并且成株有强壮的根系,能有效利用稳定的深层土壤水。种子形状与土壤种子库寿命有关,小粒、近圆球形种子易于形成持久种子库[1]。推测如果未来降水持续增加,唐古特白刺种子会变小,适应荒漠环境的变化。
表型多样性是遗传多样性与环境异质性的综合体现[1]。唐古特白刺种子性状在不同增雨处理的群体内都具有丰富的多样性。其中种子千粒重的变异系数最大20.386 %,种子长宽比的变异系数最小为5.650 %,说明种子的长宽比有较稳定的遗传特征。表型变异研究对其遗传改良、优良种质资源保存及开发利用具有极其重要的意义。表型变异必然蕴藏着遗传变异,即表型变异愈大,可能存在的遗传变异就越大[27-28]。种子大小对其萌发特性的影响进行初步研究表明唐古特白刺大小种子平均发芽率37.6 %。其中小种子发芽最早,在发芽初期有较大优势;大种子发芽最晚,但具最高发芽率。总发芽率和出苗率均为特大粒(0 %处理)>大粒(25 %处理)>中粒(50 %处理)>小粒(75 %处理)>特小粒(100 %处理),但进一步的方差分析则表明,种子大小差异对唐古特白刺种子萌发和出苗没有显著的影响。因此,可以认为唐古特白刺的种子萌发率和出苗率不受种子大小的影响。这与普遍认为的植物大种子具有较高萌发率和出苗率的观点相同[29-30]。唐古特白刺不同分级种子的出苗以特小粒种子最早,且出苗早期出苗率相对最高;特大粒种子最晚,但具最高出苗率。本研究是以持续模拟增雨近十年种子的形态指标进行分析,由于所测的指标及所考虑的气候因子较少,唐古特白刺发育状况、生理生化特性以及外因影响如何还需进一步探讨,可能此时不同增雨对种子的遗传性状尚未得到充分表达与稳定,这些早期表现优良的性状是否与晚期萌发生长相关尚需进一步研究。
5 结 论
不同增雨量对唐古特白刺种子长度、宽度、体积、表面积、长宽比及千粒重均有影响,其中种子体积、表面积和千粒重在不同增雨处理间差异均显著,且三者与增雨量均显著负相关。因此,降雨增加后唐古特白刺会产生更小的种子来适应环境变化。特小粒种子发芽最早,特大粒种子发芽最晚,但具最高出苗率,大小不同的唐古特白刺种子在萌发率和出苗率上并不存在显著差异。
猜你喜欢
农业工程学报(2022年14期)2022-10-19
经济林研究(2022年2期)2022-07-20
贵州畜牧兽医(2022年3期)2022-06-28
现代园艺(2021年23期)2021-12-01
辽宁农业科学(2021年5期)2021-11-03
课堂内外(高中版)(2021年9期)2021-01-17
新农业(2020年18期)2021-01-07
大灰狼(2020年3期)2020-04-24
山西林业科技(2020年4期)2020-03-09
大众文艺(2017年1期)2017-03-08
