
水痘-带状疱疹病毒主要衣壳蛋白ORF40单克隆抗体的制备及初步应用①
2019-06-10 01:01蔡琳俐付文锟潘德全叶江辉夏宁邵
中国免疫学杂志 2019年9期
蔡琳俐 王 玮 付文锟 潘德全 叶江辉 朱 桦 程 通 夏宁邵
(厦门大学生命科学学院,国家传染病诊断试剂与疫苗工程技术研究中心,厦门361102)
水痘-带状疱疹病毒(Varicella-zoster virus,VZV)是水痘及带状疱疹两种疾病的共同病原体。水痘一般发生于人的婴幼儿及儿童时期,由VZV初次感染所引起,随后VZV可长期潜伏于人的神经节,在患者年老或免疫力降低时可能再激活引发带状疱疹,愈后常导致后遗神经痛,严重影响患者生活质量[1-5]。
VZV是疱疹病毒科α亚科的双链DNA病毒,其基因组大小约为125 kb,包含至少70个开放阅读框(Open reading frame,ORF),编码至少68个氨基酸[6]。VZV病毒颗粒具有典型的疱疹病毒结构,从内到外分别为:包裹DNA基因组的核衣壳、无固定形状的皮层、宿主细胞膜及病毒糖蛋白组成的囊膜[7]。衣壳的形成是VZV成熟病毒颗粒产生过程的第一步且也是关键步骤之一。根据与VZV 同源性相近的1型单纯疱疹病毒(Herpes simplex virus 1,HSV-1)的基因组推测,VZV 衣壳组装可能需要ORF20、ORF23、ORF33、ORF33.5、ORF40 和ORF41 6种病毒蛋白,然而目前关于确切VZV衣壳组成蛋白的种类还有待证实[8]。VZV ORF40类似同源蛋白HSV-1 VP5(UL19)可能是最主要的病毒衣壳蛋白,其基因组包含4 224个碱基,编码1 396个氨基酸,蛋白分子量大小约为153 kD。
近年来,关于VZV ORF40蛋白的研究报道较少。Chaudhuri等[8]发现ORF40蛋白不具备核定位信号,但可在与另一个具有核定位信号的衣壳蛋白ORF23共表达时被携带进入细胞核,这可能对VZV衣壳在细胞核内组装十分重要。另外,Inoue等[9]发现一种新型的抗VZV药物(35B2)可通过与ORF40蛋白相互作用抑制VZV衣壳组装,进而影响VZV的复制,这说明ORF40蛋白在VZV衣壳组装过程中扮演重要角色。然而,目前ORF40蛋白在VZV感染细胞中参与的蛋白-蛋白相互作用及其在VZV衣壳组装过程中的功能等都尚不清楚,而获得其特异性单克隆抗体(monoclonal antibody,mAb)对ORF40蛋白性质与功能研究具有非常重要的意义。
本研究应用生物软件分析设计并合成了ORF40蛋白中亲水性与免疫原性较好的多肽片段, 将合成多肽与牛血清蛋白偶联后免疫小鼠,采用杂交瘤技术制备获得特异性高、反应活性良好的抗VZV ORF40蛋白的单克隆抗体。同时,本研究将该抗体用于Western blot与免疫荧光检测,对VZV感染细胞中的ORF40蛋白表达、定位以及对纯化VZV衣壳颗粒ORF40蛋白的包含情况进行了初步探究。
1 材料与方法
1.1材料
1.1.1主要试剂 次黄嘌呤、胸腺嘧啶、氨基喋呤、弗式完全佐剂和不完全佐剂、TRITC标记的 GAM IgG抗体、牛血清白蛋白(BSA)、1-(3-二甲氨基丙基)-3-乙基碳二亚胺(EDC)、山羊血清购自美国Sigma公司;DAPI购自美国Invitrogen公司; Anti-β-tubulin的小鼠单克隆抗体、HRP 标记的 GAM IgG抗体购自美国 CST 公司;Super Signal West Pico Substrate试剂盒购自Pierce公司;RPMI1640、DMEM/F12培养基、胰酶及胎牛血清(FBS)购自 Thermo Fisher Scientific 公司;银染、ELISA试剂盒、纯化实验等其他常用化学试剂购自西陇化工股份有限公司。Anti-gE、anti-ORF23的小鼠单克隆抗体由本实验室自行生产。
1.1.2菌种与质粒 大肠杆菌DH5α购自TianGen公司;pCDNA3.1-ORF40真核表达质粒由北京金唯智生物技术有限公司合成。
1.1.3细胞株、病毒株和实验动物 小鼠骨髓瘤细胞SP2/0、人胚胎肾细胞293T、人视网膜色素上皮细胞ARPE-19由本实验室保存;VZV pOka病毒株由美国罗格斯大学朱桦教授实验室提供[10];6~8周龄的BALB/c 雌鼠和F1小鼠购自上海斯莱克实验动物有限公司。
1.2方法
1.2.1多肽合成与偶联 从NCBI获得VZV pOka毒株的ORF40基因与氨基酸序列,将获得的序列信息通过Protean软件进行亲疏水性及免疫原性分析,选择ORF40 N端亲水性较高的包含15个氨基酸的多肽片段ORF40-N1-15:MTTVSCPANVITTTE,委托上海生工进行合成。分别取2 mg多肽干粉溶于500 μl 2-(N-吗啉)乙磺酸一水合物(MES)缓冲液、2 mg BSA 溶于200 μl MES缓冲液以及10 mg EDC溶于1 ml ddH2O,先将500 μl的多肽溶液与200 μl的BSA溶液混合,再加入100 μl EDC溶液,室温混匀2 h 后至PBS透析,-20℃保存备用。
1.2.2单克隆抗体制备 将BSA偶联的ORF40多肽抗原与等体积的弗氏完全佐剂(初次免疫)或不完全佐剂(加强免疫)混合成均匀的乳化状态后,以多点注射的方式、按100 μg/只的剂量注射到BALB/c 雌鼠四肢皮下,首次免疫后分别在第2和第4周进行加强免疫,每次免疫前对小鼠进行眼内眦采血。最后在第6周使用相同的抗原进行脾脏免疫,3 d后取免疫小鼠的脾脏与SP2/0按照常规方法进行细胞融合获得杂交瘤细胞[11]。采用有限稀释法对杂交瘤细胞进行克隆化,取杂交瘤细胞培养上清通过ELISA方法筛选特异性好、与免疫原反应强的单克隆。经过至少2轮克隆化筛选后,最终获得能稳定分泌单克隆抗体的杂交瘤细胞株。将获得的单克隆杂交瘤细胞扩大培养后,注射至F1小鼠腹腔,待小鼠腹部肿胀后收集腹水,并通过Protein-A亲和层析柱纯化获得单克隆抗体。
1.2.3筛选杂交瘤细胞的间接ELISA检测 将合成的ORF40蛋白多肽用碳酸钠/碳酸氢钠缓冲液(pH9.6)进行稀释,使其浓度为100 μg/ml,再按100 μl/孔 的量包被在96孔微孔板上,4℃包被过夜。用1×PBST缓冲液洗板3次后,每孔加入200 μl含有3%BSA的PBS缓冲液于37℃孵育2 h,甩干后真空密封保存备用。检测时每孔加入50 μl杂交瘤细胞上清,37℃孵育1 h。用1×PBST缓冲液清洗5次后,每孔加入100 μl HRP标记的GAM IgG抗体(1∶5 000稀释)于37℃孵育30 min。再次用1×PBST清洗5次后,每孔加入100 μl显色液于37℃孵育15 min,加入50 μl终止液后在酶标仪上进行读值。
1.2.4VZV病毒衣壳的初步纯化 准备5×107个已感染VZV pOka且完全病变的ARPE-19细胞,清洗后消化并收集细胞悬液。经Dounce匀浆处理后1 000 g、4℃离心10 min,收取上清。加入1%NP40处理后,通过20%~50%(w/v)蔗糖密度梯度离心,使用Beckman SW28转头17 000 r/min、4℃离心1 h,取最下层分离液至PBS溶液中透析,透射电镜观察并在-80℃保存备用。
1.2.5银染检测 向纯化的病毒衣壳或VZV pOka感染的ARPE-19细胞裂解液加入蛋白上样缓冲液,沸水浴处理10 min后跑SDS-PAGE胶。将蛋白胶置于固定液(200 ml甲醇加250 μl甲醛后,ddH2O定容至500 ml)中固定10 min。ddH2O洗2次后,加入0.02%(w/v)硫代硫酸钠反应1 min。ddH2O洗2次后,加入0.5%(w/v)AgNO3反应10 min左右。ddH2O洗2次后,加显色液显影后终止,再置于ddH2O中拍照。
1.2.6Western blot检测 分别将转染真核表达质粒pCDNA3.1-ORF40或pCDNA3.1后48 h收获的293T细胞以及感染(感染复数MOI=0.3)或未感染VZV pOka病毒后48 h收获的ARPE-19细胞,用含1 mmol/L PMSF和1%Triton X-100 的PBS溶液裂解后获得蛋白样品。经12%SDS-PAGE分离后通过湿电转膜仪使胶上的蛋白转移至硝酸纤维素膜上,使用5%脱脂牛奶4℃封闭过夜,以VZV ORF40单克隆抗体(1 μg/ml)为一抗于37℃孵育1 h,用PBST缓冲液洗涤5次后,以HRP标记的GAM IgG抗体(按1∶5 000稀释)为二抗于37℃孵育30 min,用PBST洗涤5次后使用Super Signal West Pico Substrate试剂盒显色并拍照。
1.2.7免疫荧光检测 分别将293T细胞和ARPE-19细胞接种于预先加入洁净玻片的24孔细胞培养板中,质粒pCDNA3.1-ORF40或pCDNA3.1转染到293T细胞,VZV pOka病毒(MOI=0.001)感染或不感染ARPE-19细胞,37℃培养48 h。随后在室温环境下,使用4%多聚甲醛固定细胞,15 min后PBS洗1次。含0.3%Triton X-100的PBS通透10 min,PBS洗1次。山羊血清封闭1 h,再以VZV ORF40单克隆抗体(1 μg/ml)为一抗于37℃孵育1 h。PBS洗3次后,以TRITC标记的GAM IgG为二抗避光孵育30 min,DAPI染色5 min,PBS洗涤3次,封片后使用激光共聚焦显微镜进行拍照。
2 结果
2.1VZV ORF40单克隆抗体的制备与鉴定 本研究将合成多肽ORF40-N1-15偶联BSA后免疫小鼠,通过杂交瘤技术、克隆化结合间接ELISA筛选,获得1株可稳定分泌抗VZV ORF40蛋白的单克隆抗体的杂交瘤细胞株,经腹水生产与纯化获得相应的单克隆抗体10F2。为进一步鉴定其特异性、反应性,使用单抗10F2对转染pCDNA3.1-ORF40或pCDNA3.1 48 h的以及未做处理的293T细胞进行Western blot和免疫荧光检测。Western blot检测结果显示,单抗10F2可与转染pCDNA3.1-ORF40的293T细胞裂解液有良好的特异性反应,在130~180 kD 的蛋白marker之间检出清晰且单一的蛋白条带(图1A),约153 kD,与ORF40蛋白的理论大小相一致,同时与未处理和转染pCDNA3.1的293T细胞裂解液无任何反应。免疫荧光检测结果显示,单抗10F2与转染pCDNA3.1-ORF40的293T细胞有较强的特异性反应,且发现外源表达的ORF40蛋白主要弥散分布在细胞质中,同时与转染pCDNA3.1空载的293T细胞没有非特异性反应(图1B)。以上结果表明,单抗10F2可特异识别VZV ORF40蛋白,可用于后续实验。
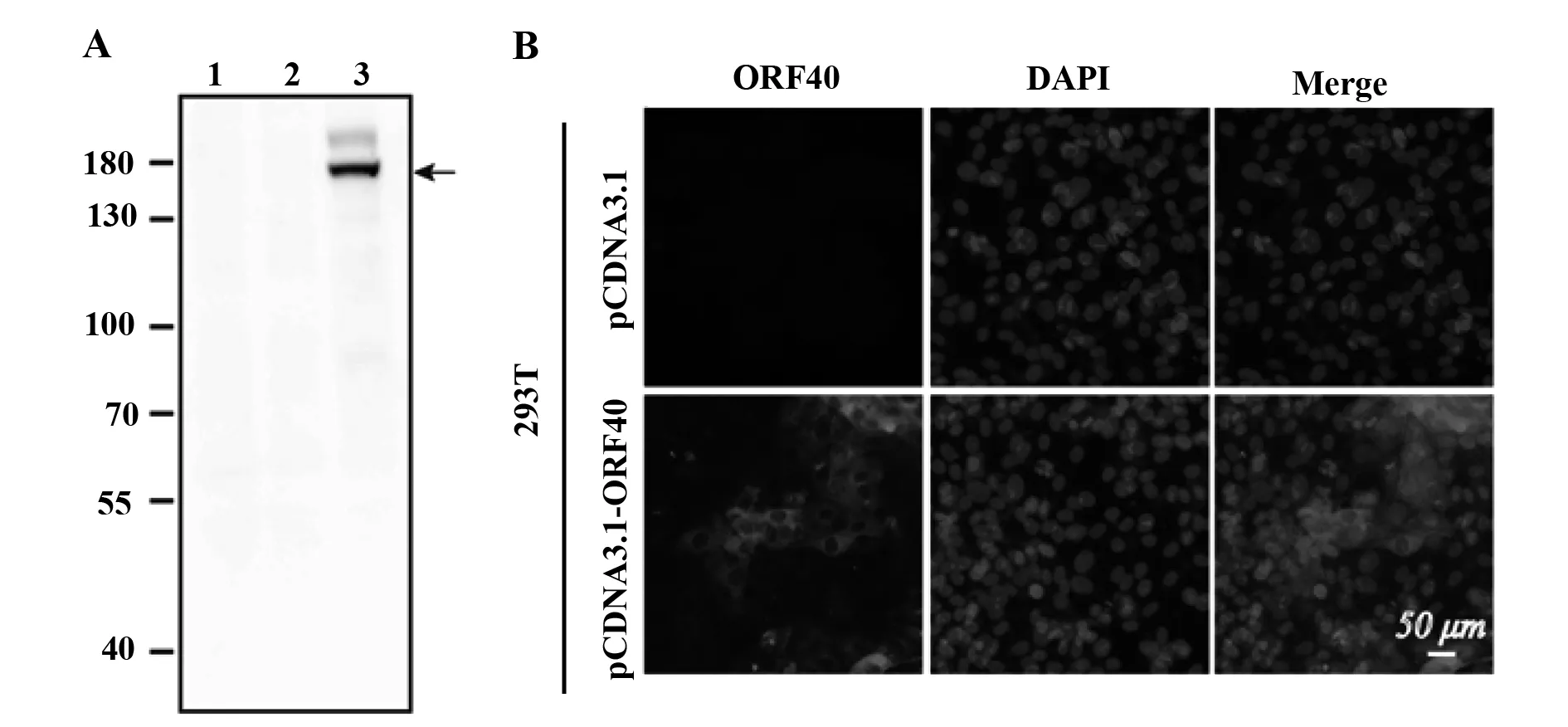
图1 抗VZV-ORF40单克隆抗体10F2性质鉴定Fig.1 Characterization of anti-VZV-ORF40 mAb 10F2Note:A.Western blot of cell extracts of 293T cells transfected or not with pCNDA3.1 or pCNDA3.1-ORF40 using anti-VZV-ORF40 mAb 10F2.1.Untreated 293T cells;2.293T cells transfected with pCNDA3.1;3.293T cells transfected with pCNDA3.1-ORF40.B.Immunofluorescence analysis of 293T cells transfected with pCNDA3.1 or pCNDA3.1-ORF40 using anti-VZV-ORF40 mAb 10F2.

图2 ORF40在VZV感染细胞中的表达与亚细胞定位Fig.2 Expression and subcellular localization of ORF40 in VZV-infected cellsNote:A.Western blot of cell extracts of ARPE-19 cells uninfected or infected with VZV pOka using anti-VZV-ORF40 mAb 10F2.1.Uninfected ARPE-19 cells;2.ARPE-19 cells infected with VZV pOka.B.Immunofluorescence analysis of ARPE-19 cells uninfected or infected with VZV pOka using anti-VZV-ORF40 mAb 10F2.
2.2VZV 感染细胞中ORF40的Western blot及免疫荧光检测 收集VZV pOka感染48 h后的ARPE-19细胞及未感染的ARPE-19阴性细胞,分别裂解后使用单抗10F2对其进行Western blot检测。结果显示,单抗10F2与VZV pOka感染的ARPE-19细胞裂解液发生特异性反应,且不与未感染的ARPE-19细胞发生反应,其检出的蛋白条带在130~180 kD之间,约为153 kD(图2A),与质粒转染表达的ORF40蛋白(图1A)以及理论大小一致。
进一步使用单抗10F2对VZV pOka感染48 h后的ARPE-19细胞及未感染的ARPE-19阴性细胞做免疫荧光检测。结果显示,单抗10F2同样与VZV感染的ARPE-19细胞发生较强的特异性反应,且发现与单独外源表达的ORF40细胞质定位不同,本实验检出的ORF40蛋白绝大部分定位于细胞核内,呈弥散分布或者在核周呈点状分布(图2B,白色箭头),部分细胞的细胞质内也能检测到ORF40(图2B,绿色箭头),这样的定位变化可能是由于ORF40蛋白和其他病毒蛋白相互作用,共同参与细胞核内的衣壳组装而引起的。同时,单抗10F2与未感染的ARPE-19细胞没有任何非特异性反应。
2.3VZV衣壳中ORF40蛋白的Western blot检测 为进一步探究单抗10F2是否可用于对VZV衣壳中ORF40蛋白的检测,本研究通过蔗糖密度梯度离心法从VZV感染的ARPE-19细胞中初步纯化获得VZV衣壳颗粒,通过银染(图3A)和透射电镜(图3B)分析可见获得的VZV衣壳纯度较好。随后,通过Western blot分析发现(图3C),ORF40蛋白与另一种小衣壳蛋白ORF23均能在纯化的VZV衣壳中检出,且无法检出病毒囊膜蛋白(如gE),也基本不包含细胞成分(如β-tubulin),而以上蛋白在VZV感染细胞裂解液中均能检出。该结果证明,单抗10F2对VZV衣壳中ORF40蛋白同样具有高效且特异的识别能力。

图3 10F2可用于VZV衣壳中ORF40蛋白检测Fig.3 Detection of ORF40 contained in VZV capsids using mAb 10F2Note:A.Silver staining analysis of purified VZV capsids and cell extracts of ARPE-19 cells infected with VZV pOka using anti-VZV-ORF40 mAb 10F2;B.Transmission electron microscopy of purified VZV capsids;C.Western blot analysis of purified VZV capsids and cell extracts of ARPE-19 cells infected with VZV pOka using anti-VZV-ORF40 mAb 10F2.1.Purified VZV capsids;2.ARPE-19 cells infected with VZV pOka;M.Protein marker.
3 讨论
VZV的有效扩散依赖于大量感染性病毒颗粒的产生,而高度有序的病毒衣壳蛋白元件的合成与衣壳组装是VZV感染与制周期的关键步骤,为设计抗病毒药物提供了良好靶标。虽然通过HSV-1的同源推测已经大致确定了VZV衣壳蛋白的种类及功能,但是关于VZV衣壳蛋白的性质、功能、蛋白-蛋白相互作用及其组装机制仍缺少研究报道。
本研究以VZV主要衣壳蛋白ORF40为研究对象,选取其N端的第1~15个氨基酸多肽片段偶联BSA作为免疫原,通过小鼠免疫、杂交瘤技术及3轮克隆化筛选获得了1株可稳定分泌抗VZV ORF40蛋白的杂交瘤单抗细胞,并将纯化获得的单抗命名为10F2。通过Western blot检测,单抗10F2可以特异性识别质粒转染的293T细胞和VZV感染的ARPE-19细胞中表达的ORF40蛋白,显示出约153 kD 条带。本研究将单抗10F2用于免疫荧光检测,发现VZV ORF40蛋白在单独质粒外源表达时因缺乏核定位信号而在细胞质内弥散定位,而在VZV感染细胞中主要是细胞核定位。而且,在VZV感染细胞中ORF40蛋白除了在细胞核内弥散定位,也呈核周点状聚集分布,这些点状部位可能是病毒集中组装或运送出核的场所。ORF40在VZV感染细胞中的定位变化可能与其和其他病毒蛋白、特别是衣壳蛋白相互作用并入核组装VZV衣壳颗粒相关。在HSV-1中,ORF40的同源蛋白VP5就是通过与其他衣壳蛋白VP19C 或VP22a 相互作用进入细胞核,进而再与pre-VP22a 组装成病毒壳粒的基本框架[12,13]。另外,VZV感染细胞的细胞质内偶尔也能检测到ORF40蛋白,这可能是出核后聚集在胞质内准备或正在进行二次组装过程的衣壳颗粒。本研究还应用单抗10F2对纯化的VZV衣壳颗粒进行检测,Western blot检测结果显示该单抗对衣壳颗粒中的ORF40蛋白同样具有较好的特异性识别能力,该结果也为ORF40蛋白属于VZV衣壳蛋白提供了直接证明。该单抗可用于后续VZV衣壳蛋白的成分分析与相互作用研究。
综上所述,本研究成功制备了抗VZV ORF40蛋白的单克隆抗体10F2,该抗体特异性强、反应性好。本研究应用该抗体确定了VZV感染细胞内ORF40蛋白的表达、亚细胞定位以及VZV衣壳颗粒包含的ORF40,这些工作为今后深入开展VZV ORF40蛋白性质、功能以及VZV衣壳组装相关研究奠定基础。
猜你喜欢
实用肿瘤学杂志(2022年3期)2022-11-30
医院管理论坛(2022年7期)2022-10-14
解放军医学院学报(2022年1期)2022-04-02
科学导报(2021年82期)2021-12-17
锦绣·中旬刊(2021年10期)2021-08-23
昆明医科大学学报(2021年2期)2021-03-29
中国现代医药杂志(2020年10期)2020-12-14
皮肤性病诊疗学杂志(2020年4期)2020-09-02
证券市场红周刊(2019年6期)2019-06-11
医药前沿(2019年7期)2019-01-05
