
去甲基化酶ALKBH5高、低表达食管鳞癌细胞株的构建
2019-05-31 02:59:18方婷晓翟坚学林桃燕别亚男肖东蔡开灿
山东医药 2019年12期
方婷晓,翟坚学,林桃燕,别亚男,肖东,蔡开灿
(1 南方医科大学南方医院,广州510515;2 南方医科大学肿瘤研究所;3 南方医科大学比较医学研究所暨实验动物中心)
食管癌是一种恶性肿瘤,其发病率在全部恶性肿瘤中排第9位[1]。目前我国每年新发食管癌病例47.7万,死亡病例37.5万[2]。食管癌分为鳞癌和腺癌两种类型,其中90%以上为食管鳞癌(ESCC)[3]。由于许多食管癌患者发现的时候已经是中晚期,因此,研究ESCC的相关分子标志物,找到相关靶点来预测ESCC的发展和预后显得非常重要。RNA修饰是一种极为重要的基因表达过程调节方式,参与真核生物的多种病理生理过程[4]。N6-甲基腺苷化修饰(N6-methyladenosine,m6A)是一种极为常见的RNA甲基化修饰,参与真核生物mRNA的运输、剪切、翻译及加工等过程,它的异常变化常常导致相应下游靶蛋白的变化,从而导致癌症的发生[5]。ALKBH5是m6A的一个去甲基化酶,是Alk B家族成员之一,亦是肥胖相关基因(FTO)的同源物[6]。研究[7~10]显示,ALKBH5与乳腺癌、神经胶质瘤和胰腺癌的增殖和侵袭转移相关。但是目前有关ALKBH5在ESCC中作用的研究较少。本研究通过慢病毒感染技术,构建了高、低表达ALKBH5的ESCC细胞株,为进一步探讨ALKBH5在ESCC中的调控作用提供细胞模型。
1 材料与方法
1.1 细胞、质粒及试剂 ESCC细胞株(Eca109细胞、Kyse510细胞、TE-13细胞)由中山大学中山医学院李隽教授惠赠,培养基为含10%胎牛血清的DMEM。人源胚胎肾细胞293T 来源及培养方法见文献[11]。高表达ALKBH5的质粒pLV-ALKBH5及对照质粒pLV-con购自广州复能基因有限公司,上述2个质粒中均带有GFP蛋白和Flag标签蛋白,用于后续感染细胞的筛选。低表达ALKBH5的质粒pLKO-shALKBH5-1、pLKO-shALKBH5-2(考虑到不同的干扰序列可能干扰效果不同,本实验选用了两个干扰序列进行了两个位点的干扰)及对照质粒pLKO-shSCR由美国哈佛医学院施扬教授惠赠[12],上述3个质粒中均带有嘌呤霉素抗性,用于后续感染细胞的筛选。慢病毒包装质粒psPAX2、pMD2.G由瑞士日内瓦大学Didier Trono博士惠赠。质粒提取及纯化试剂盒购自QIAGEN公司。DMEM细胞培养基购自美国Corning公司。胰蛋白酶购自Gibco BRL公司。TRIzol、PrimeScriptTMRT Reagent Kit、SYBR Premix Ex TaqTM荧光定量 PCR 试剂盒均购自TaKaRa公司。蛋白分子量Marker购自Thermo公司。BCA蛋白浓度测定试剂盒购自南京凯基生物科技发展有限公司。脂质体转染剂Lipofectamine 2000、RIPA 裂解液、高糖DMEM、Opti-MEM I Medium购自 Invitrogen公司。ALKBH5抗体(ABE547)购自MILLIPORE公司。内参GAPDH 抗体(10494-1-AP)购自武汉三鹰Proteintech公司。FLAG抗体(F1804)购自Sigma公司。辣根过氧化物酶标记的羊抗兔IgG二抗(BS13278)、辣根过氧化物酶标记的羊抗鼠IgG二抗(BS12478)购自Bioworld公司。
1.2 293T细胞分组和质粒转染 将生长状态良好的293T细胞于25 cm2培养瓶内培养,分别记为A、B、a、b、c组,待细胞融合80%~90%后,按照质量比pMD2.G∶psPAX2∶目的质粒=8∶15∶20的比例进行质粒转染。A组293T细胞用pLV-ALKBH5质粒转染,B组293T细胞用pLV-con质粒转染,a组293T细胞用pLKO-shALKBH5-1质粒转染,b组293T细胞用pLKO-shALKBH5-2质粒转染,c组293T细胞用pLKO-shSCR质粒转染。质粒转染72 h后收集各组病毒上清,3 000 rpm离心15 min,用0.45 μm滤器过滤后分装于1.5 mL离心管。
1.3 高表达ALKBH5的ESCC细胞株的构建 分别用A、B组细胞上清液感染Eca109细胞和Kyse510细胞,感染6~8 h后用含10% FBS的DMEM换液1次,48 h后于倒置荧光显微镜下观察。pLV-ALKBH5质粒和pLV-con质粒携带的GFP蛋白在倒置荧光显微镜下为绿色荧光,观察到绿色荧光说明细胞感染成功。选取各组感染成功的细胞,采用流式细胞术筛选GFP蛋白(+)的细胞待测。
1.4 低表达ALKBH5的ESCC细胞株的构建 分别用a、b、c组细胞上清液感染Eca109细胞和TE-13细胞,感染6~8 h后用含10% FBS的DMEM换液1次,待细胞融合60%~70%时用嘌呤霉素筛选细胞,选取筛选2周后仍然处于良好生长状态的细胞待测。
1.5 分别感染高、低表达ALKBH5的293T细胞上清液的ESCC细胞中ALKBH5 mRNA和蛋白的检测 ①采用qRT-PCR法检测ALKBH5 mRNA。收集各组细胞,TRIzol提取总RNA,按照逆转录试剂盒说明书将RNA逆转录成cDNA,以GAPDH为内参,进行PCR扩增。引物序列如下:ALKBH5 上游引物为5′-CGGCGAAGGCTACACTTACG-3′,下游引物为5′-CCACCAGCTTTTGGATCACCA-3′;GAPDH 上游引物为5′-ACCCAGAAGACTGTGGATGG-3′,下游引物为5′-TCTAGACGGCAGGTCAGGTC-3′。建立10 μL PCR反应体系:cDNA 1 μL、SYBR Green 5 μL、ddH2O 3 μL、上下游引物各0.5 μL。PCR反应条件为:95 ℃预变性30 s,40 个循环(95℃ 5 s、60 ℃ 30 s、72 ℃ 30 s)。以 2-ΔΔCt表示ALKBH5 mRNA 的相对表达量。实验重复3次,取平均值。②采用Western blotting法检测ALKBH5蛋白。收集细胞并提取蛋白,BCA法测蛋白浓度,蛋白变性后上样,恒压电泳和恒流转膜后封闭2 h,封闭后将带有目的蛋白的条带放在装有一抗的抗体孵育盒中,将盒子置于4 ℃冰箱慢摇过夜,次日拿出室温慢摇30 min后回收一抗,PBS清洗3次,每次10 min,然后二抗孵育条带1 h,再次用PBS清洗3次,每次10 min,最后用凝胶成像系统进行显像,用image J软件分析条带灰度值,计算受检蛋白的相对表达量。受检蛋白的相对表达量=受检蛋白条带灰度值/内参条带灰度值。实验重复3次,取平均值。
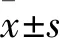
2 结果
分别感染高、低表达ALKBH5细胞上清液的ESCC细胞中ALKBH5 mRNA和蛋白的检测结果显示,感染A组细胞上清液的Eca109细胞、Kyse510细胞中ALKBH5 mRNA和蛋白的相对表达量均显著高于感染B组细胞上清液的Eca109细胞、Kyse510细胞(P均<0.05),感染a、b组细胞上清液的Eca109细胞、TE-13细胞中ALKBH5 mRNA和蛋白的相对表达量均显著低于感染c组细胞上清液的Eca109细胞、TE-13细胞(P均<0.05)。本实验成功构建高、低表达ALKBH5的ESCC细胞株,详见表1、2。
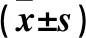
表1 感染高表达ALKBH5细胞上清液的ESCC细胞中ALKBH5 mRNA和蛋白的相对表达量比较
注:与B组相比,﹡P<0.05。
表2 感染低表达ALKBH5细胞上清液的ESCC细胞中ALKBH5 mRNA和蛋白的相对表达量比较
注:与c组相比,﹟P<0.05。
3 讨论
RNA表观遗传学修饰是转录后水平的调控方式,目前已鉴定了150多种修饰类型,它们分布于转运RNA、核糖体RNA、mRNA、非编码小RNA和长链非编码RNA上,其中m6A是最常见的一种发生在真核生物mRNA上的RNA修饰[13~19]。随着高通量测序技术的发展,证实转录本的m6A修饰水平受编码器(甲基转移酶)、读码器(结合蛋白)和消码器(去甲基化酶)的动态调控,表明RNA的m6A修饰是一个动态可逆的过程[19]。作为真核生物mRNA上除了5′帽子结构外最多的转录后修饰,m6A成为近年来RNA研究领域的热点。m6A修饰受特异性的甲基转移酶和去甲基化酶动态调控,进而影响干细胞的多能性和分化[7~10]。近年来,随着m6A第一个去甲基化酶FTO的发现,首次证实了RNA修饰存在可逆性,扩大了表观基因组学的丰富程度[20]。研究[13]显示,m6A可通过去甲基化酶FTO和ALKBH5可逆性调节去甲基。FTO和ALKBH5均是Alk B家族成员,其中ALKBH5可催化m6A去甲基化。
研究[7~10,18]显示,ALKBH5的沉默除了会影响mRNA的核输出及精子形成外,还与乳腺癌、神经胶质瘤和胰腺癌的进展和侵袭转移相关。研究[7]发现,ALKBH5在乳腺癌样干细胞中高表达,且与NANOG基因协同表达,高表达ALKBH5可以增加NANOG的蛋白水平和稳定性,提示ALKBH5可以影响NANOG mRNA的去甲基化或者对NANOG mRNA m6A水平有间接的影响。研究[7]同时证明,抑制ALKBH5或缺氧诱导因子的表达,可以激活缺氧乳腺癌细胞中ALKBH5的表达,同时减少NANOG表达,并在体内靶向乳腺癌样干细胞。另一项研究[9]表明,ALKBH5在神经胶质瘤样干细胞中高表达,生存分析提示ALKBH5是神经胶质瘤患者的负性预后因子,沉默ALKBH5可抑制干细胞增殖和肿瘤形成。为进一步探讨ALKBH5在神经胶质瘤中的作用,研究[9]还进行了微阵列分析和m6A 的高通量测序,在沉默ALKBH5的细胞中下调的206个基因中筛选出13个与表型及增殖相关的候选基因,进行分析并筛选出转录因子FOXM1,提示ALKBH5可以通过影响FOXM1的去甲基化来影响神经胶质瘤样干细胞的增殖。He等[10]最新研究表明,高表达ALKBH5可以抑制胰腺癌的侵袭和迁移,并首次发现ALKBH5可以通过下调长链非编码RNA KCNK15-AS1的甲基化来抑制胰腺癌的侵袭转移能力。以上研究均提示ALKBH5有可能成为新的抗癌靶点,为癌症的诊断及预后判断提供新的依据,但是具体的分子机制和生物学功能还有待阐明。
纵观现有文献,ALKBH5尚未在ESCC有过相关的研究,本课题组前期通过20例临床标本检测得到,ALKBH5在ESCC中普遍低表达,故本研究基于此基础,构建了慢病毒介导的ALKBH5高、低表达的ESCC细胞株。本研究预测,高表达ALKBH5之后,ESCC的增殖能力可能会减弱,侵袭转移能力减弱,同时将会抑制ESCC干细胞的生长,肿瘤球数量变少,干细胞的一系列标志蛋白和mRNA水平可能会上升;干扰ALKBH5之后,ESCC的增殖可能会加快,侵袭转移能力增强,同时促进ESCC干细胞的生长,肿瘤球数量变多,直径更大,干细胞的一系列标志蛋白和mRNA水平可能会下调。
综上所述,为进一步阐明ALKBH5在食管鳞癌中的生物学作用及机制,本研究分别将高表达ALKBH5的质粒pLV-ALKBH5和低表达ALKBH5的质粒LKO-shALKBH5-1、LKO-shALKBH5-2感染ESCC细胞株,成功构建高、低表达ALKBH5的ESCC细胞株,为进一步探讨ALKBH5在ESCC发生发展中的作用提供了细胞模型。
猜你喜欢
酿酒科技(2023年10期)2023-11-23 11:09:42
食品安全导刊(2019年27期)2019-12-09 07:34:16
名城绘(2019年4期)2019-10-21 05:09:13
广东农业科学(2017年10期)2018-01-25 03:22:15
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
医学研究杂志(2015年11期)2015-06-10 06:44:03
饲料博览(2015年12期)2015-04-04 04:28:36
中国当代医药(2015年16期)2015-03-01 02:03:11
中国医药导报(2015年27期)2015-02-28 22:08:02
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
